1. Introduction
Cotton is a Kharif season crop grown mainly for feed, fiber, and oil in the Punjab and Sindh regions of Pakistan. These are considered hot regions since the temperature reaches 47 °C during the growing season. Environmental stresses such as heat, and drought affect cotton plants by impeding normal physiological processes which lead to morphological abnormalities and yield reduction [
1]. Plants mostly invest their defense in the most valuable sections, such as reproductive parts under various stress conditions. Cotton production is vulnerable to abiotic stresses, particularly during the growth stages of blooming and boll formation, which have become more frequent as our climate changes [
2]. Any stress during this stage abruptly reduces the yield. Numerous efforts have been made to understand the physiological, molecular, and genetic pathways of the cotton plant related to sustaining yield under stress conditions [
3,
4].
The reproductive efficiency of the cotton crop is negatively impacted by temperatures above 32 °C in a variety of ways, including reduced metabolism as well as suppression of photosynthesis, pollination, fertilization, and crop growth rate [
5]. Heat and drought stress causes male gametes to undergo metabolic and structural changes that result in meiotic abnormalities or premature spore abortion [
6]. It also results in poor pollen germination and short pollen tube growth in cotton [
7,
8]. It was reported that pollen germination is better in flowers that have been pollinated under the canopy of the plant as compared to flowers that are directly exposed to sunlight and pollinated during high-temperature stress [
9].
Yield reduction is also associated with certain changes in metabolic and biochemical pathways in plant cells, i.e., excessive accumulation of reactive oxygen species (ROS) such as H
2O
2, singlet oxygen, hydroxyl ions, etc., during stress conditions [
10]. As a result of a dramatic accumulation of ROS during stress, programmed cell death has been observed in developing pollen grains [
11]. Hence, ROS scavenging through the action of antioxidants in anthers has a role in maintaining pollen viability under abiotic stress [
12]. Under high temperature or water deficit conditions, the role of the cell membrane in maintaining cell osmotic balance may be impeded due to leakage of electrolytes [
13]. In cotton, temperatures over 35 °C increased membrane leakage and reduced leaf size [
14]. High canopy temperature adversely affects the chlorophyll content in leaf tissues and lowers the rate of photosynthesis and carbohydrate production [
15]. Reduction in carbohydrate content is also associated with decreased lint yield [
16]. Cotton plants will shed bolls when they are stressed, thus boll retention drops significantly under harsh environmental conditions [
17,
18].
It was noticed that different portions of a single cotton plant of a cultivar exhibit variable responses to stress during reproductive and vegetative phases. Although every cell in a plant has the same genetic material, the different behavior might be due to epigenetic [
19,
20] or other effects. Every cell expresses itself according to the stimulus received from the environment. Young leaves are more resistant to insect damage compared to old ones [
21]. So, every part of the plant faces a different environment. As a result, these positions phenotypically behave differently. Cultivars also differ in canopy shape and intra-plant morphological features. Moreover, cultivars are grown in the same region exhibit variation among them. Environmental and genotypic effects both contribute to the phenotype. Therefore, the objective of this study was to identify inter- and intra-cultivar variability for physiological as well as morphological attributes associated with the yield of seed cotton under heat stress conditions.
4. Discussion
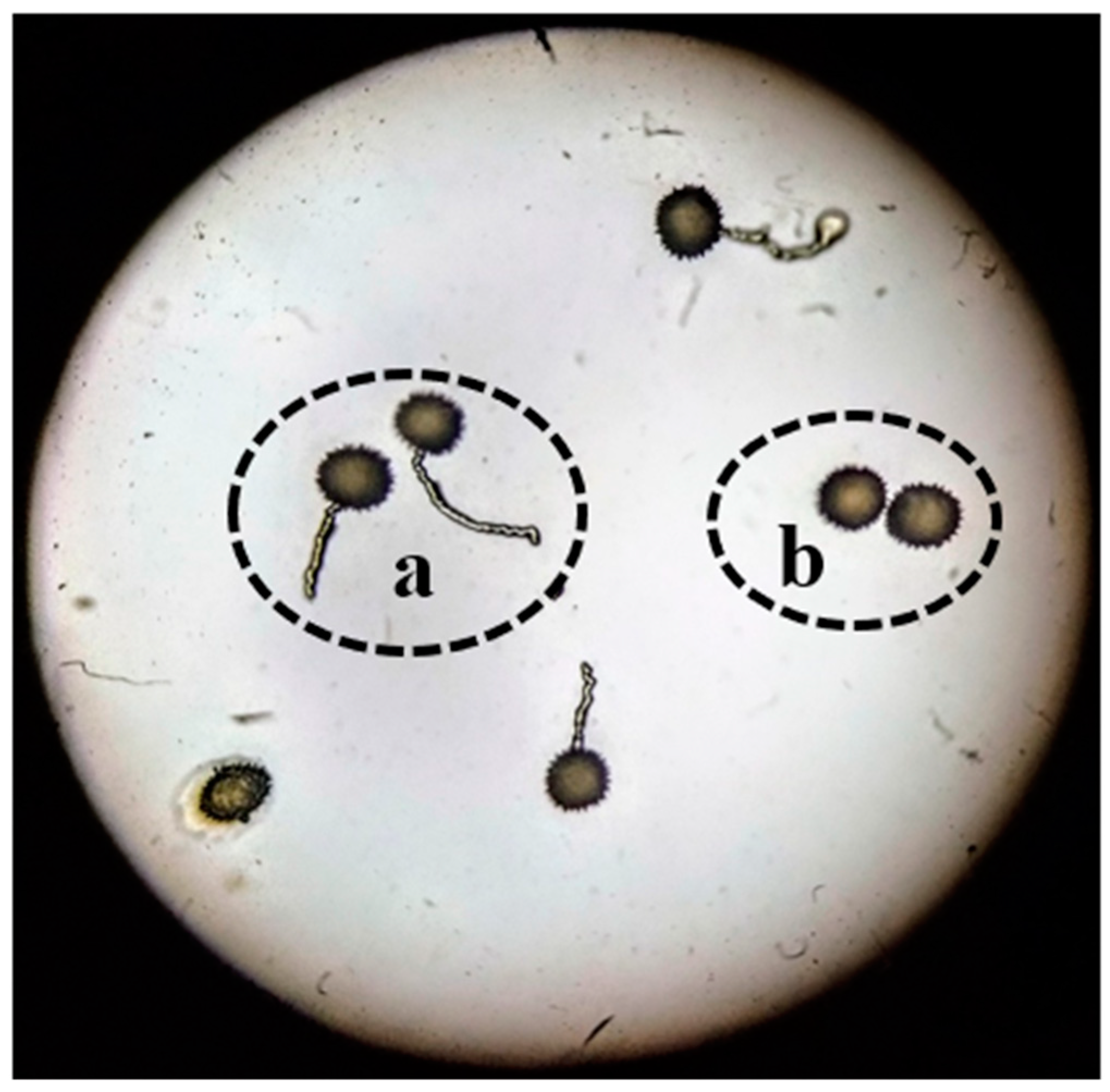
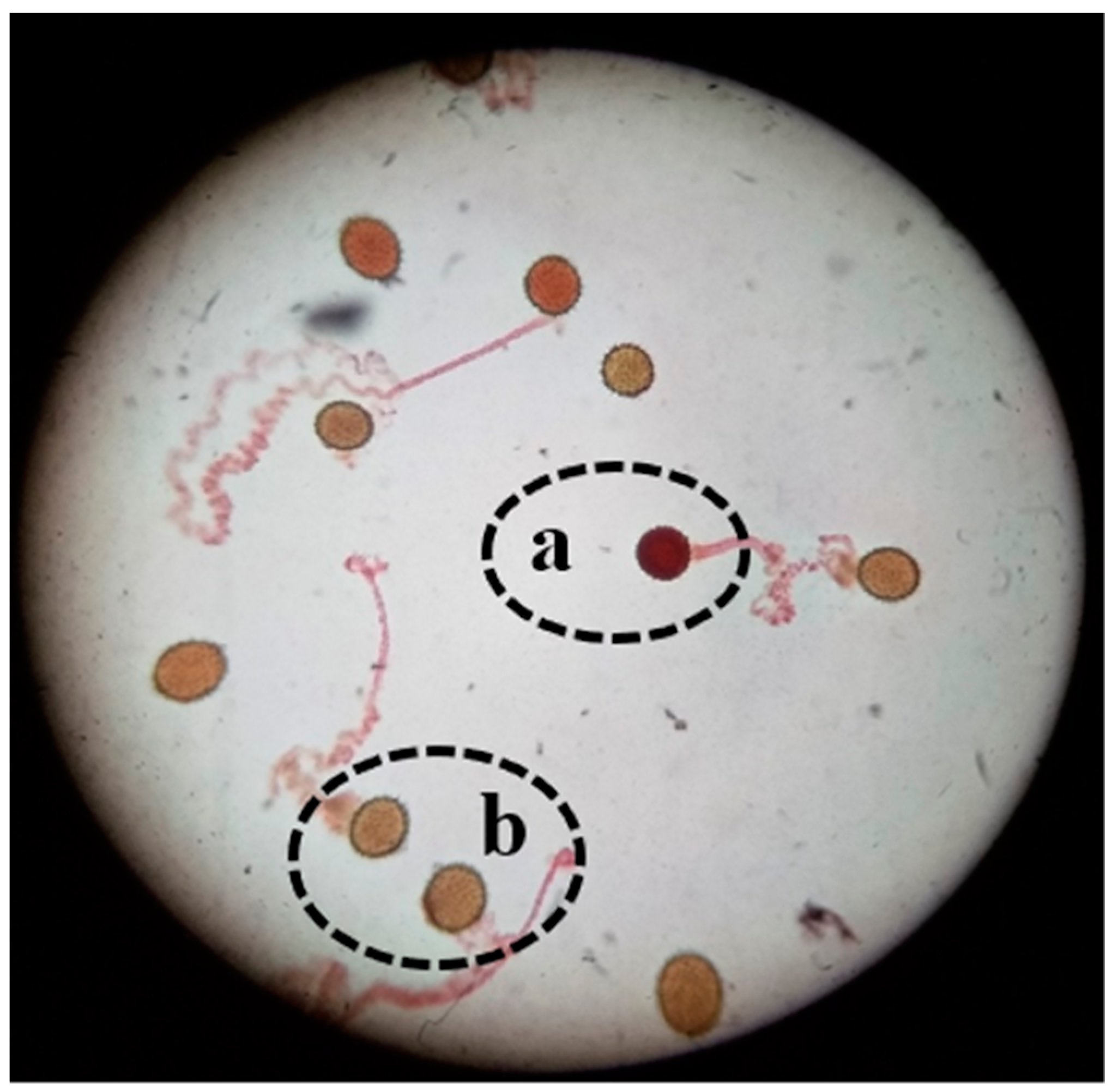
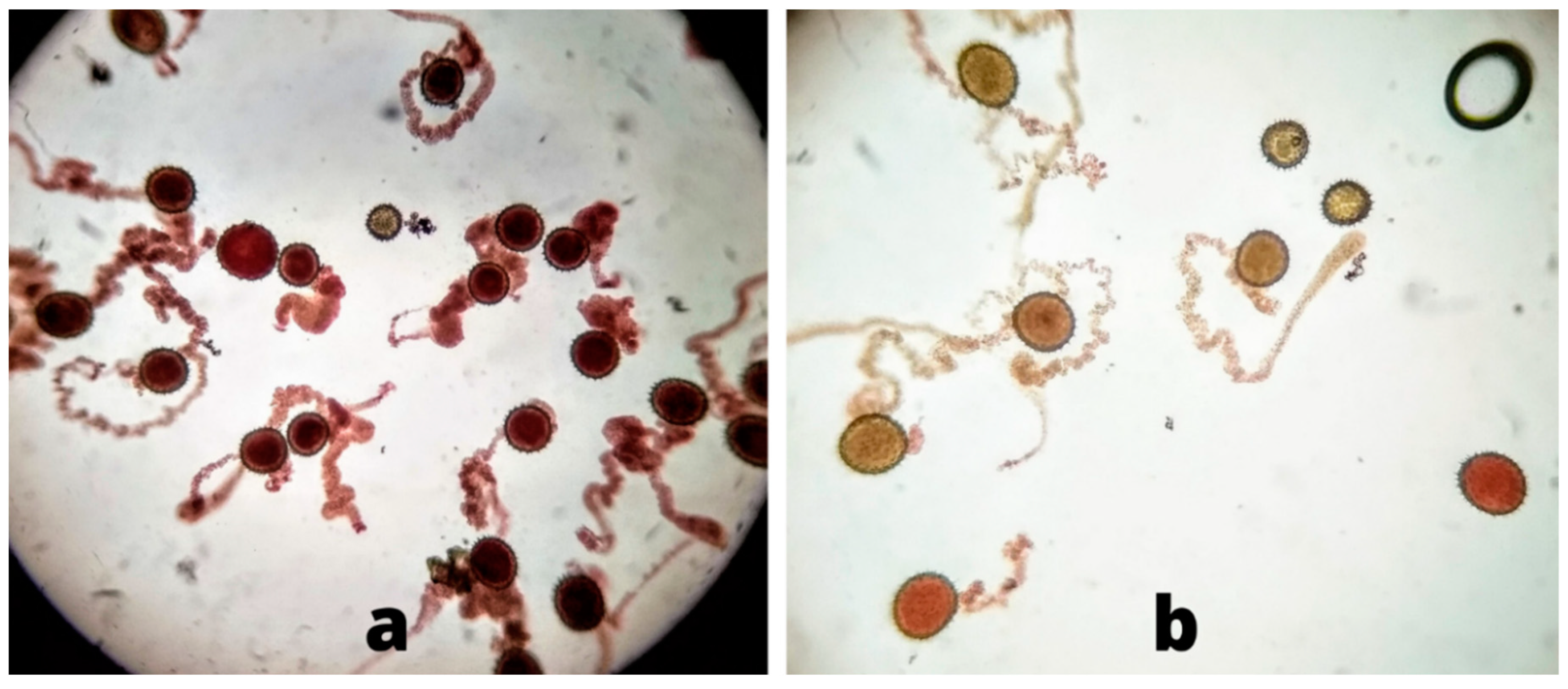
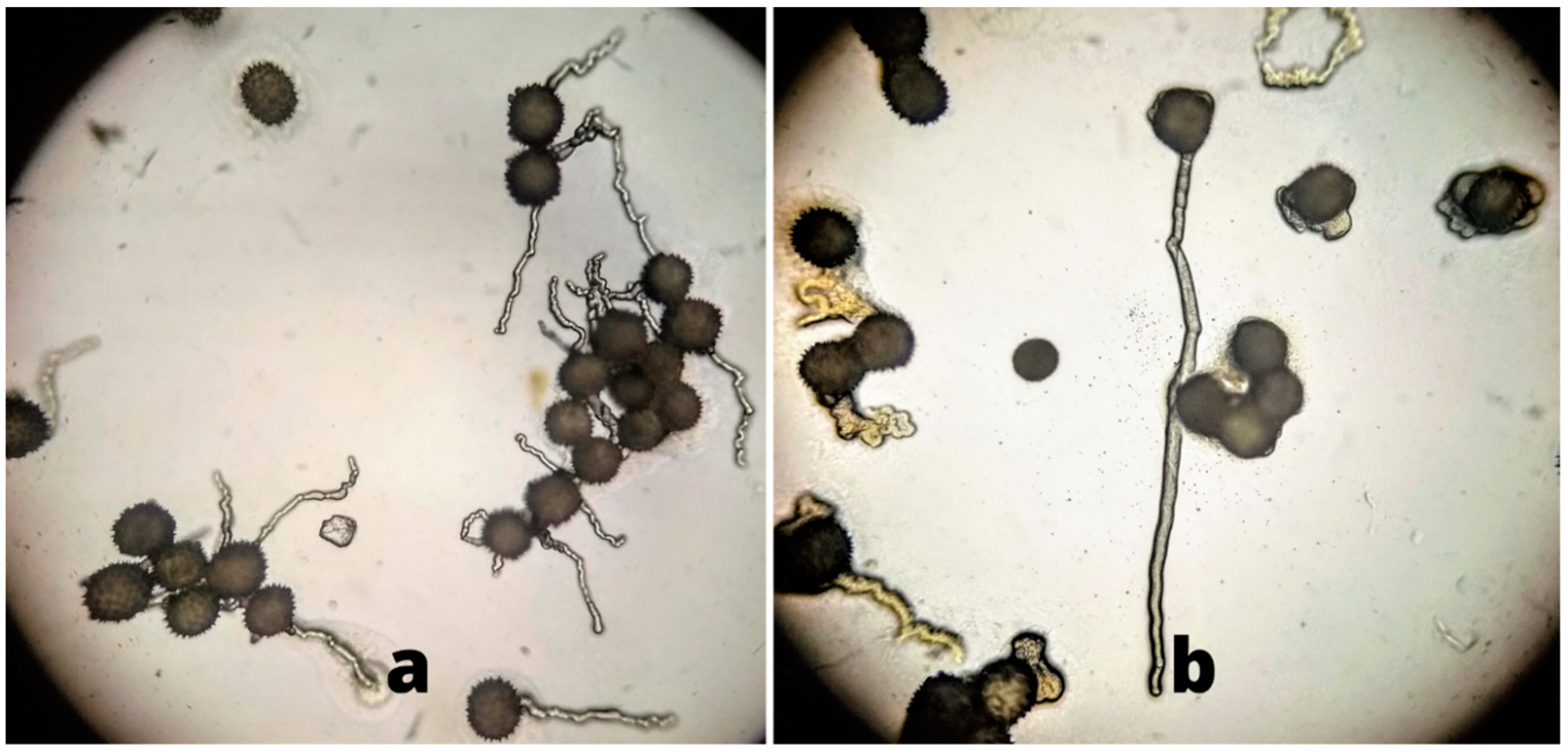
Higher pollen germination and viability percentages from flowers under the canopy of the plant as compared to flowers in direct sunlight were observed in this study. As the temperature in the experimental region in Pakistan rises to 47 °C during the time of cotton flowering, this damages the lipid as well as protein parts of the pollen membrane, thus resulting in decreases in pollen viability [
12]. Pollen viability was determined by analyzing dehydrogenase enzyme activity in the pollen grains—if the enzyme is active, viable pollen grains change to a red color after TTC staining. However, there may be damage to the pollen grains that reduces germination despite this enzyme activity. It has been reported that the distribution of cell organelles such as mitochondria, vacuoles, and endoplasmic reticulum of pollen cells become disturbed under high temperatures. Lipid and starch granules are also reduced in pollen cells during heat stress [
8].
In our study, lower pollen germination was observed as compared to pollen viability. Most pollen could not develop the pollen tube required for germination, likely due to metabolic or structural abnormalities of pollen grains [
31]. Drought or heat stress significantly lowers carbohydrate metabolism in the pistil, resulting in a lower energy supply to the pollen tube in the style, thus leading to a failure of fertilization [
32]. Under excessively high temperatures, heat shock proteins (HSPs) work to stabilize proteins that were damaged when exposed to stressful conditions. As the expression of HSPs varies between genotypes, some genotypes showed good pollen germination even in the top portion of the plant [
33].
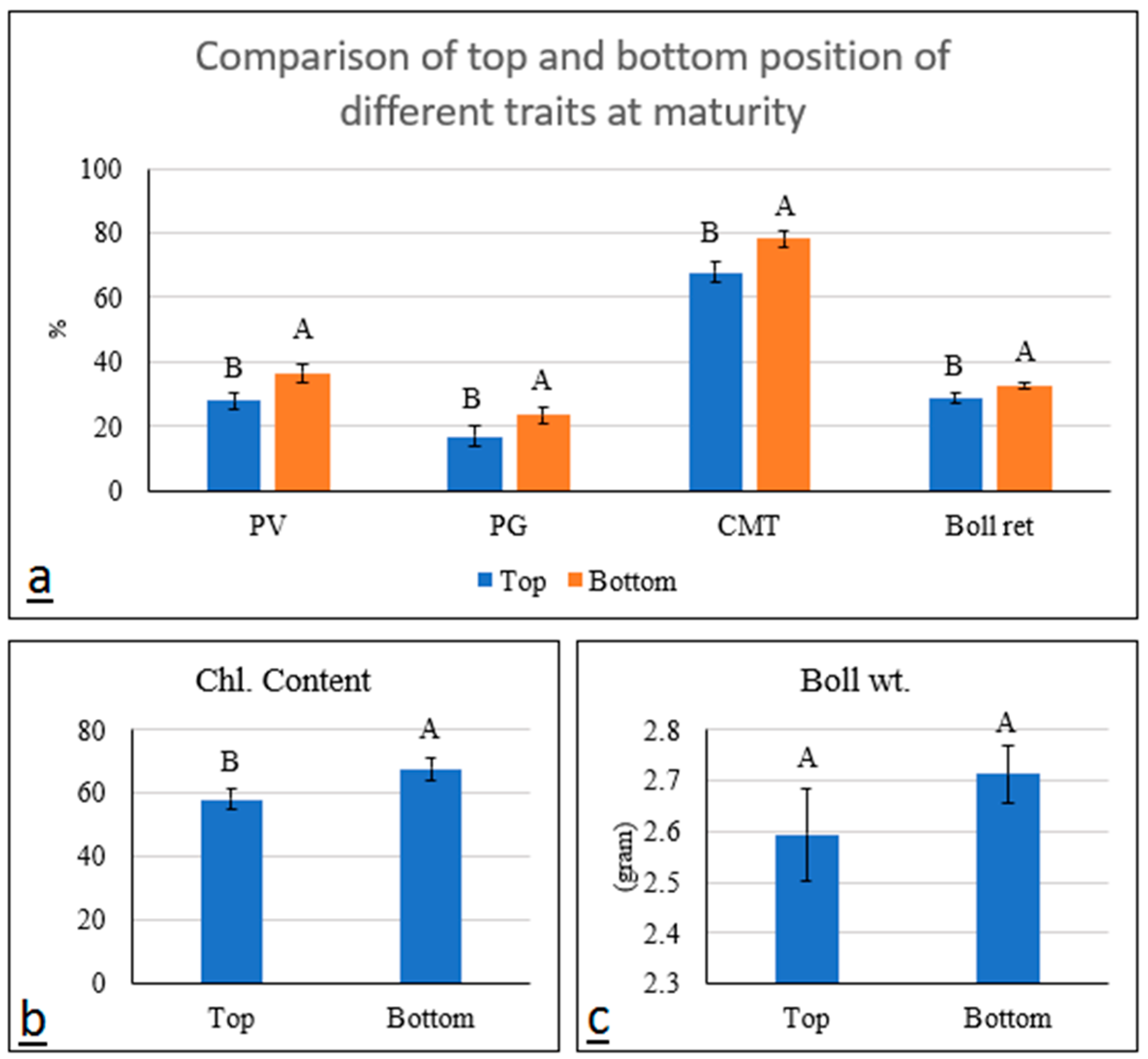
Genotypic variability for CMT has been previously reported [
3,
34]. Here, we have observed CMT differences between plant positions. The significant differences among cultivars are due to several factors including cuticle thickness, secondary metabolites, and heat shock proteins [
35,
36,
37]. Lower CMT estimates in top leaves were due to sunlight exposure. The membranes of leaves facing direct sunlight in high-temperature conditions were more prone to damage. Sun rays cause oxidative damages to both lipid and protein parts of the cellular membranes and cause the leakage of electrolytes [
38]. UV radiation from the sun causes irreversible damage to plant pigments [
39]. It causes conformational changes in the structure of nucleic acids, proteins, and macromolecules in the cell and degrades the chlorophyll pigment [
40,
41]. Heat stress directly affects the flow of fluid through the cell membrane as relative electrical conductivity increases with temperature [
42,
43].
Since the chlorophyll contents under the canopy were higher as compared to the top position, it has been assumed that chlorophyll loses its integrity under direct sunlight. In addition to direct sunlight, the higher temperature in the top portion of the plant also causes chlorophyll damage [
44]. Heat stress that denatures thylakoid membranes results in a loss of chlorophyll [
45]. Moreover, the enzymes required for the synthesis of chlorophyll and its normal activity were also denatured under high-temperature conditions [
46]. As a result, photosynthetic activity was reduced in the top portion of plants. On average, the genotypes AA-933 and CYTO-608 had good heat tolerance features in bottom positions; therefore, these genotypes would be useful as parents in a breeding program
Ascorbic acid has the potential to mitigate the negative effects of stress. It acts in ROS scavenging and maintains the integrity of membranes, including the thylakoid membrane [
47]. So, ascorbic acid could be used to overcome the heat stress problem. The cell membrane thermostability of cotton crops can be improved significantly by applying the foliar application of 40 mg L
−1 ascorbic acid [
48].
Boll weight is positively associated with seed cotton yield. It is a complex polygenic trait that depends upon numerous factors namely, the weight of seed, seed size, protein and oil content within the seed, and cellulose deposition during fiber development and maturity [
49]. It is one of the most important characters linked to improved yield, and significant variation for this trait has been reported in germplasm [
50,
51]. Although the genotypes used in this experiment were significantly different for boll weight, no significant differences for this trait were recorded between the top and bottom portions of the same genotypes. Retention of bolls during the developmental period varied significantly between the top and bottom branches. The bottom branches tend to hold more bolls as compared to the top branches. It was noted that the genotypes with higher pollen viability and germination also retained more bolls. This study revealed that the heat tolerance ability of the genotypes was associated with boll retention while heat stress has been considered one of the major factors in bolls dropping before maturity [
52,
53]. Thus, high temperature in the top portion of a plant due to direct exposure to sunlight can explain retaining a lower number of bolls in this portion of the plant.
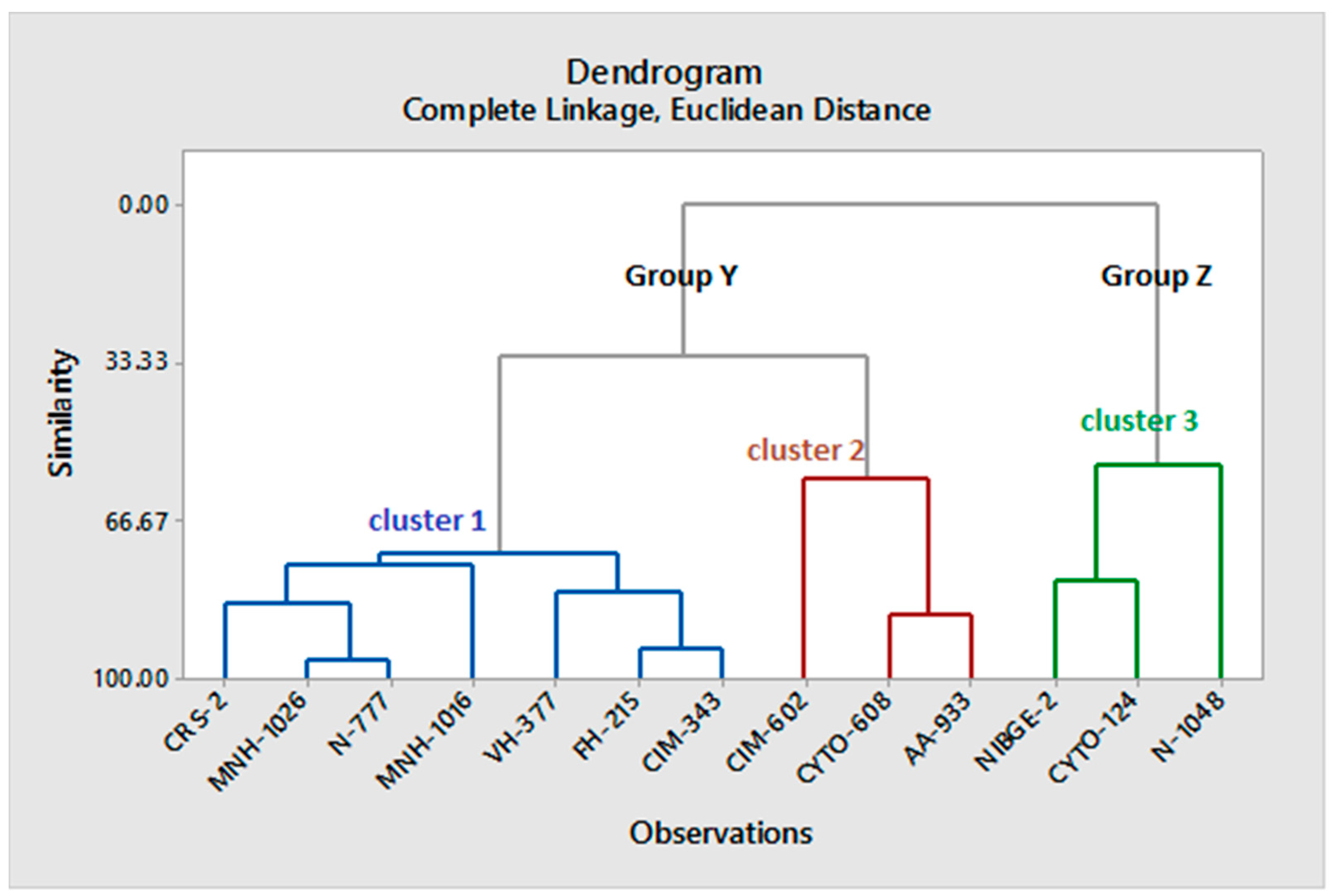
The variability was found between the genotypes as shown in
Table 3. Cluster analysis has revealed that CIM-602, Cyto-608, and AA-933 grouped in cluster 2 performed well and these genotypes could be used further in any breeding program. Since all genotypes are grown in the Punjab region of Pakistan, these are therefore acclimatized to this environment. These genotypes share some common, as well as different phenotypes, which showed variability based on six traits used in this study. The variability was also observed in the cotton genotypes cultivated in the Punjab region of Pakistan by khan [
54].
This study provides an understanding of the role of flowering in the top and bottom portions of the cotton plant in response to high-temperature stress because high temperature is a major factor in reducing yield. It is assumed that by increasing the vegetative growth and leaf surface area, the shading effect can be increased. The spreading-type behavior of the cotton plant could be able to produce more shading. The shading effect will reduce canopy temperature and hence yield could be increased. Likewise, screening for early maturing cultivars and for having more branches on the bottom part of the plant could be beneficial because the bottom branches have shown more productivity than top branches. Keeping in view the importance of the study, another study may be conducted to assess the temperature of the microenvironment i.e., the temperature of leaf, bud, and/or boll at the top and bottom regions of each genotype, followed by correlation analysis with each trait to understand the relationship of various traits during heat stress.




 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}