Behavioural Response of Triatoma infestans (Klug) (Hemiptera: Reduviidae) to Quinazolines

Abstract
:Introduction
Results

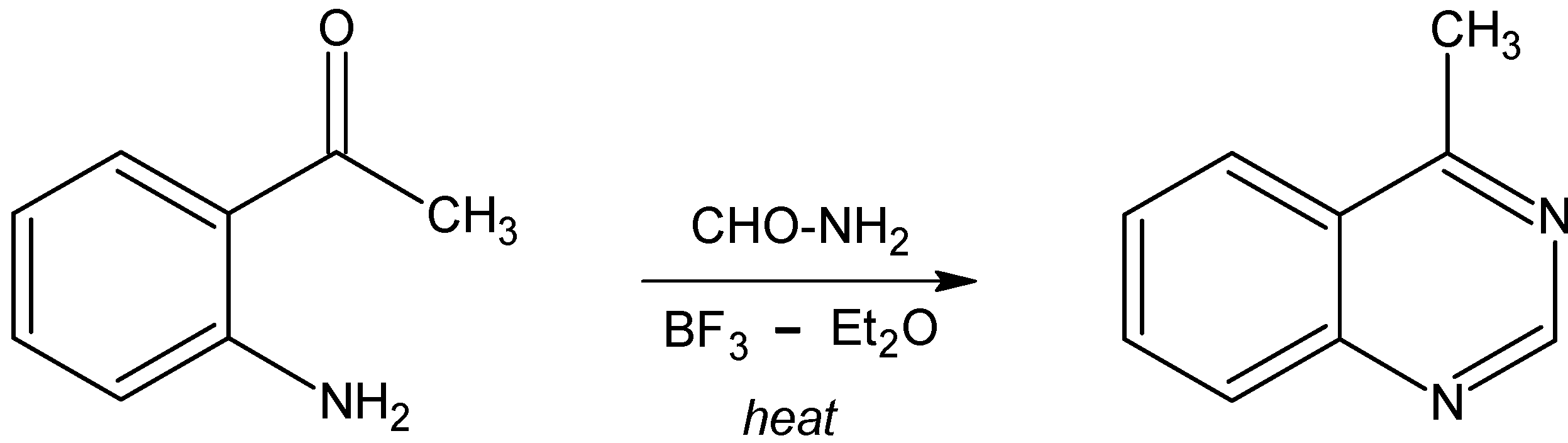{kind=link}
{kind=link}
| Behavioural response | Control | 4-methylquinazoline + 2,4-dimethylquinazoline | |
| (5 + 5) ng | (50 + 50) μg | ||
| Fifth instar nymphs Positive recordings (%) | 30 | 60 | 100 |
| Mean number of visits (± SE) | 0.4 ± 0.2a | 0.8 ± 0.3a | 2.2 ± 0.3b |
| Females Positive recordings (%) | 30 | 60 | 70 |
| Mean number of visits | 0.4 ± 0.2a | 1.2 ± 0.4a | 2.4 ± 0.7b |
| Males Positive recordings (%) | 30 | 40 | 60 |
| Mean number of visits (± SE) | 0.7 ± 0.4 a | 0.9 ± 0.5a | 1.2 ± 0.4a |
| Behavioural response | Control | 4-methyl quinazoline (50 μg) | 2,4-dimethyl quinazoline (50 μg) |
|---|---|---|---|
| Positive recordings (%) | 25 | 65 | 35 |
| Mean number of visits (± SE) | 0.2 ± 0.1 a | 1.5 ± 0.2 b | 0.7 ± 0.3 a |
| Behavioural response | Control | 4-methyl quinazoline (50 μg) | 2,4-dimethyl quinazoline (50 μg) |
|---|---|---|---|
| Positive recordings (%) | 30 | 70 | 80 |
| Mean number of visits (± SE) | 0.2 ± 0.1 a | 1.0 ± 0.3 a | 0.9 ± 0.2 a |
Discussion


Experimental
General
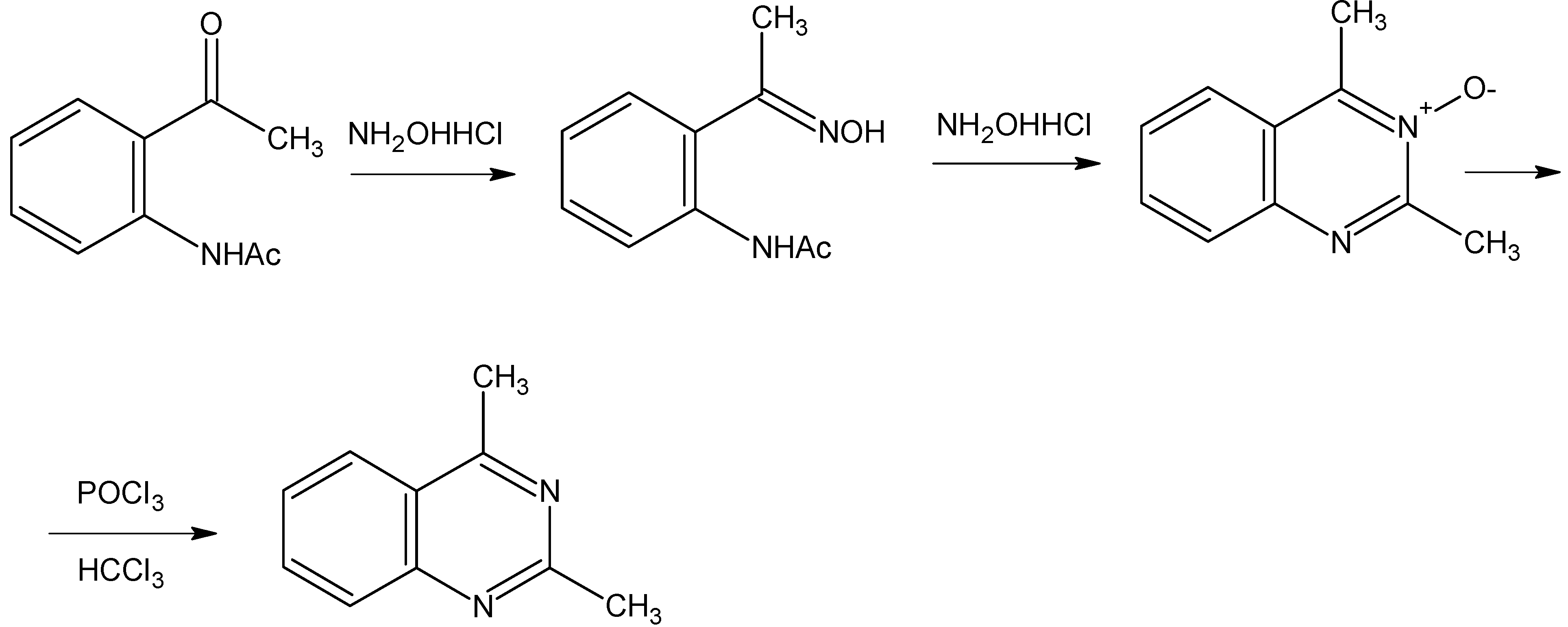
Syntheses of 4-methylquinazoline and 2,4-dimethylquinazoline.
Biological material
Bioassay
Acknowledgements
References
- Lorenzo Figueras, A.N.; Kenigsten, A.; Lazzari, C.R. Aggregation in the haematophagous bug Triatoma infestans: Chemical signals and temporal pattern. J. Insect Physiol. 1994, 40, 311–316. [Google Scholar]
- Schofield, C.J.; Patterson, J.W. Assembly pheromone of Triatoma infestans and Rhodnius prolixus nymphs (Hemiptera: Reduviidae). J. Med. Entomol. 1977, 13, 727–734. [Google Scholar]
- Taneja, J.; Guerin, P.M. Ammonia attracts the haematophagous bug Triatoma infestans: behavioral and neurophysiological data on nymphs. J. Comp. Physiol. A 1997, 181, 21–34. [Google Scholar] [CrossRef]
- López, L.C.; Morgan, E.D. Chemical investigation of aggregation behavior of Triatoma bugs (Hemiptera: Reduviidae). J. Chem. Ecol. 1995, 21, 2069–2078. [Google Scholar] [CrossRef]
- Alzogaray, R.A. Unpublished results.
- [WHO] - World Health Organisation [WHO]. Tropical Disease Research: Progress 1995-1996. Thirteenth programme report of the UNDP/World Bank/WHO Special Programme for Research and Training in Tropical diseases; World Health Organisation: Geneva, 1997. [Google Scholar]
- Schofield, C.J. Triatominae: Biología y Control; Eurocommunica Publications: Bognor Regis (U.K.), 1994. [Google Scholar]
- Wisnivesky-Colli, C.; Paulone, Y.; Pirez, A.; Chuit, R.; Gualtieri, J.; Solarz, N.; Smith, A.; Segura, E. A new tool for continuo detection of the presence of Triatominae bugs, vectors of Chagas disease, in rural households. Medicina 1987, 47, 45–50. [Google Scholar]
- Zerba, E.N. Chemical control of Chagas disease vectors. Biom. Environ. Sci. 1989, 2, 24–29. [Google Scholar]
- Byford, A.; Goadby, P.; Hooper, M.; Kamath, H.V.; Kulkarni, S.N. O-Aminophenyl alkyl-aralkyl ketones and their derivatives. Part V. An efficient synthetic route to some biologically active 4-substitued quinazolines. Indian J. Chem. 1988, 27, 396–397. [Google Scholar]
- Kovendi, A.; Kircz, M. New synthesis for quinazoline N3-oxides and 1,2-dihydroquinazoline N3-oxides. Chem. Berichte 1965, 98, 1049–1059. [Google Scholar]
- Bernard, J. Etude électrophysiologique de récepteurs impliqués dans l′orientation vers l′hote et dans l′acte hématophage chez un hémiptere: Triatoma infestans. Thesis, Univ. Rennes, France, 1974. [Google Scholar]
- Lorenzo, M.G.; Lazzari, C.R. The spatial pattern of defecation in Triatoma infestans and the role of faeces as a chemical mark of the refuge. J. Insect Physiol. 1996, 42, 903–907. [Google Scholar]
- Sample Availability: Contact the authors.
© 2005 by MDPI (http:www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Alzogaray, R.A.; Fontán, A.; Camps, F.; Masuh, H.; Santo Orihuela, P.; Fernández, D.; Cork, A.; Zerba, E. Behavioural Response of Triatoma infestans (Klug) (Hemiptera: Reduviidae) to Quinazolines. Molecules 2005, 10, 1190-1196. https://doi.org/10.3390/molecules10091190
Alzogaray RA, Fontán A, Camps F, Masuh H, Santo Orihuela P, Fernández D, Cork A, Zerba E. Behavioural Response of Triatoma infestans (Klug) (Hemiptera: Reduviidae) to Quinazolines. Molecules. 2005; 10(9):1190-1196. https://doi.org/10.3390/molecules10091190
Chicago/Turabian StyleAlzogaray, R. A., A. Fontán, F. Camps, H. Masuh, P. Santo Orihuela, D. Fernández, A. Cork, and E. Zerba. 2005. "Behavioural Response of Triatoma infestans (Klug) (Hemiptera: Reduviidae) to Quinazolines" Molecules 10, no. 9: 1190-1196. https://doi.org/10.3390/molecules10091190