Effect of Dehydroaltenusin-C12 Derivative, a Selective DNA Polymerase α Inhibitor, on DNA Replication in Cultured Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction
Results and Discussion
Effects of inhibitors on the activities of mammalian pol α

Effect of inhibitors on cell growth inhibition

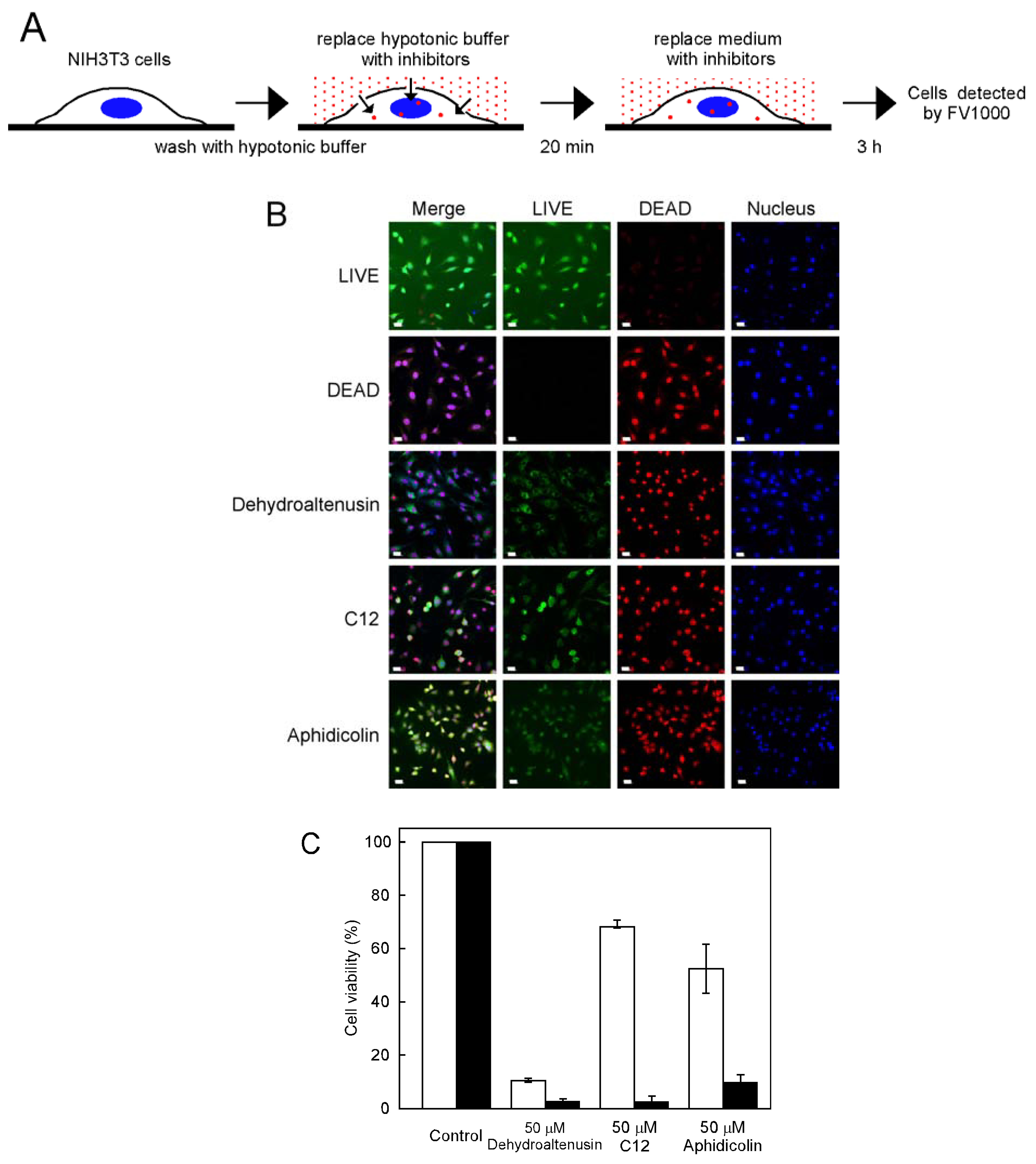
Effect of inhibitors on cell growth by using a hypotonic shift
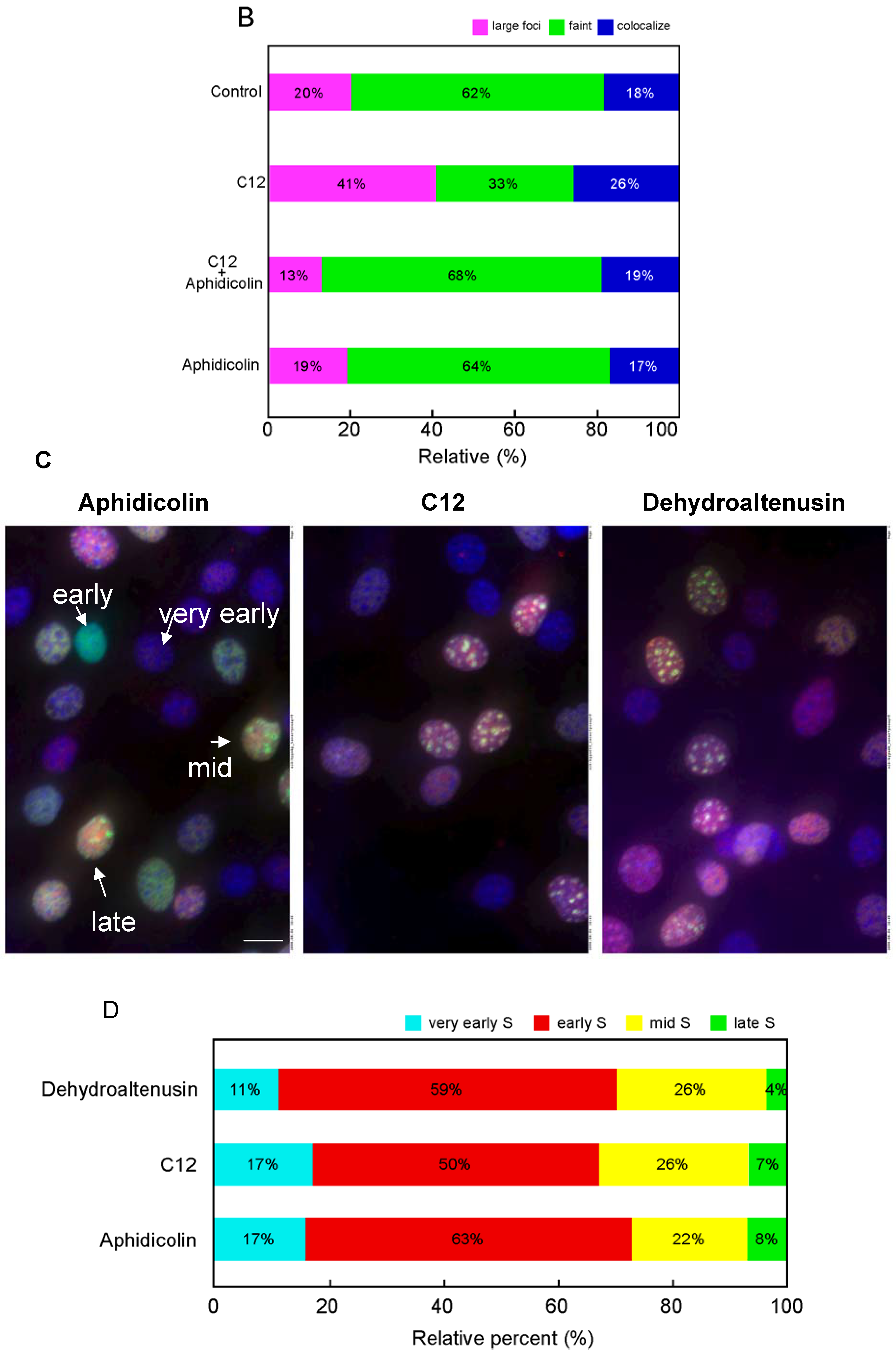
Effect of C12 on DNA replication



Conclusions
Experimental
General
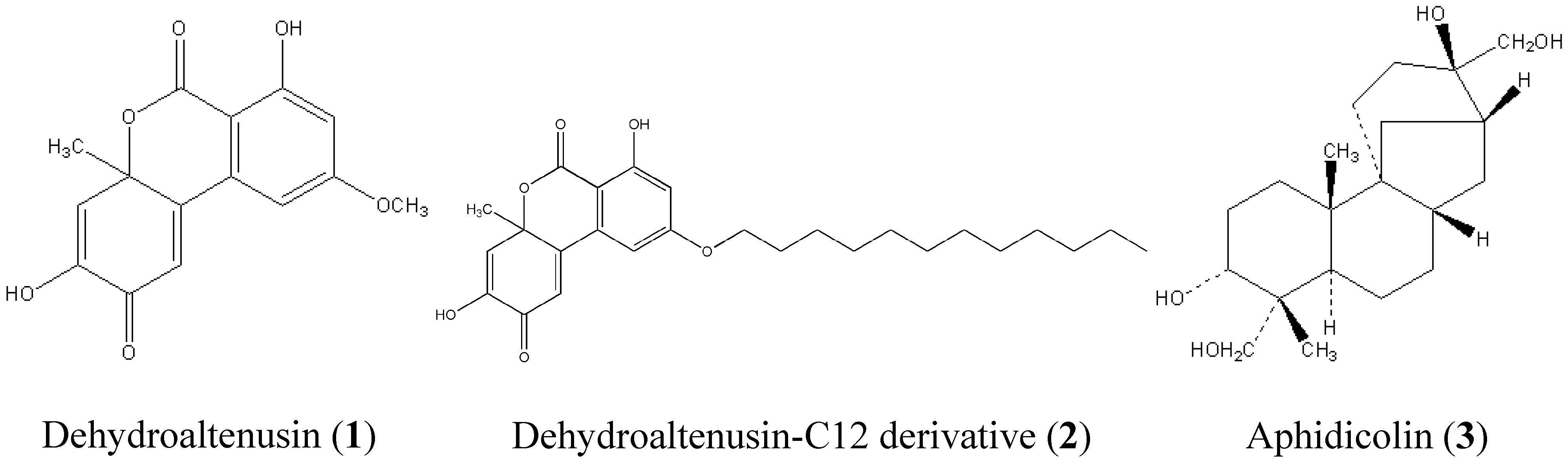
Preparation of dehydroaltenusin and dehydroaltenusin-C12 derivative
Measurement of DNA polymerase activity
Cell culture and measurement of cell viability
Hypotonic shift treatment
Immunofluorescence microscopy
Acknowledgements
References and notes
- Maga, G.; Hubscher, U. Repair and translesion DNA polymerases as anticancer drug targets. Anticancer Agents Med. Chem. 2008, 8, 431–447. [Google Scholar]
- Mizushina, Y.; Tanaka, N.; Yagi, H.; Kurosawa, T.; Onoue, M.; Seto, H.; Horie, T.; Aoyagi, N.; Yamaoka, M.; Matsukage, A.; Yoshida, S.; Sakaguchi, K. Fatty acids selectively inhibit eukaryotic DNA polymerase activities in vitro. Biochim. Biophys. Acta 1996, 1308, 256–262. [Google Scholar] [CrossRef]
- Mizushina, Y.; Tanaka, N.; Kitamura, A.; Tamai, K.; Ikeda, M.; Takemura, M.; Sugawara, F.; Arai, T.; Matsukage, A.; Yoshida, S.; Sakaguchi, K. The inhibitory effect of novel triterpenoid compounds, fomitellic acids, on DNA polymerase β. Biochem. J. 1998, 330, 1325–1332. [Google Scholar]
- Mizushina, Y.; Ohkubo, T.; Date, T.; Yamaguchi, T.; Saneyoshi, M.; Sugawara, F.; Sakaguchi, K. Mode analysis of a fatty acid molecule binding to the N-terminal 8-kDa domain of DNA polymerase β. A 1:1 complex and binding surface. J. Biol. Chem. 1999, 274, 25599–25607. [Google Scholar]
- So, A.G.; Downey, K.M. Eukaryotic DNA replication. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 129–155. [Google Scholar] [CrossRef]
- Ikegami, S.; Taguchi, T.; Ohashi, M.; Oguro, M.; Nagano, H.; Mano, Y. Aphidicolin prevents mitotic cell division by interfering with the activity of DNA polymerase-α. Nature 1978, 275, 458–460. [Google Scholar] [CrossRef]
- DePamphilis, M.L. DNA Replication in Eukaryotic Cells. Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1996; pp. 461–491. [Google Scholar]
- Lemoine, F.J.; Degtyareva, N.P.; Lobachev, K.; Petes, T.D. Chromosomal translocations in yeast induced by low levels of DNA polymerase a model for chromosome fragile sites. Cell 2005, 120, 587–598. [Google Scholar] [CrossRef]
- Nakamura, M.; Nabetani, A.; Mizuno, T.; Hanaoka, F.; Ishikawa, F. Alterations of DNA and chromatin structures at telomeres and genetic instability in mouse cells defective in DNA polymerase α. Mol. Cell. Biol. 2005, 25, 11073–11088. [Google Scholar] [CrossRef]
- Bae, S.H.; Bae, K.H.; Kim, J.A.; Seo, Y.S. RPA governs endonuclease switching during processing of Okazaki fragments in eukaryotes. Nature 2001, 412, 456–461. [Google Scholar] [CrossRef]
- Pavlov, Y.I.; Frahm, C.; Nick McElhinny, S.A.; Niimi, A.; Suzuki, M.; Kunkel, T.A. Evidence that errors made by DNA polymerase α are corrected by DNA polymerase δ. Curr. Biol. 2006, 16, 202–207. [Google Scholar] [CrossRef]
- Miller, M.R.; Ulrich, R.G.; Wang, T.S.; Korn, D. Monoclonal antibodies against human DNA polymerase-α inhibit DNA replication in permeabilized human cells. J. Biol. Chem. 1985, 260, 134–138. [Google Scholar]
- Budd, M.; Campbell, J.L. Temperature-sensitive mutations in the yeast DNA polymerase I gene. Proc. Natl. Acad. Sci. U. S. A. 1987, 84, 2838–2842. [Google Scholar] [CrossRef]
- Eki, T.; Murakami, Y.; Enomoto, T.; Hanaoka, F.; Yamada, M. Characterization of DNA replication at a restrictive temperature in a mouse DNA temperature-sensitive mutant, tsFT20 strain, containing heat-labile DNA polymerase α activity. J. Biol. Chem. 1986, 261, 8888–8893. [Google Scholar]
- Niimi, A.; Limsirichaikul, S.; Yoshida, S.; Iwai, S.; Masutani, C.; Hanaoka, F.; Kool, E.T.; Nishiyama, Y.; Suzuki, M. Palm mutants in DNA polymerases α and η alter DNA replication fidelity and translesion activity. Mol. Cell. Biol. 2004, 24, 2734–2746. [Google Scholar] [CrossRef]
- Mizushina, Y.; Kamisuki, S.; Mizuno, T.; Takemura, M.; Asahara, H.; Linn, S.; Yamaguchi, T.; Matsukage, A.; Hanaoka, F.; Yoshida, S.; Saneyoshi, M.; Sugawara, F.; Sakaguchi, K. Dehydroaltenusin, a mammalian DNA polymerase α inhibitor. J. Biol. Chem. 2000, 275, 33957–3361. [Google Scholar] [CrossRef]
- Koberna, K.; Stanek, D.; Malinsky, J.; Eltsov, M.; Pliss, A.; Ctrnacta, V.; Cermanova, S.; Raska, I. Nuclear organization studied with the help of a hypotonic shift: its use permits hydrophilic molecules to enter into living cells. Chromosoma 1999, 108, 325–335. [Google Scholar] [CrossRef]
- Pedrali-Noy, G.; Spadari, S.; Miller-Faures, A.; Miller, A.O.; Kruppa, J.; Koch, G. Synchronization of HeLa cell cultures by inhibition of DNA polymerase α with aphidicolin. Nucleic Acids Res 1980, 8, 377–387. [Google Scholar] [CrossRef]
- Dimitrova, D.S.; Todorov, I.T.; Melendy, T.; Gilbert, D.M. Mcm2, but not RPA, is a component of the mammalian early G1-phase prereplication complex. J. Cell Biol. 1999, 146, 709–722. [Google Scholar] [CrossRef]
- Liu, J.S.; Kuo, S.R.; Melendy, T. Comparison of checkpoint responses triggered by DNA polymerase inhibition versus DNA damaging agents. Mutat. Res. 2003, 532, 215–226. [Google Scholar] [CrossRef]
- Manthey, K.C.; Opiyo, S.; Glanzer, J.G.; Dimitrova, D.; Elliott, J.; Oakley, G.G. NBS1 mediates ATR-dependent RPA hyperphosphorylation following replication-fork stall and collapse. J. Cell Sci. 2007, 120, 4221–4229. [Google Scholar] [CrossRef]
- Nedelcheva, M.N.; Roguev, A.; Dolapchiev, L.B.; Shevchenko, A.; Taskov, H.B.; Stewart, A.F.; Stoynov, S.S. Uncoupling of unwinding from DNA synthesis implies regulation of MCM helicase by Tof1/Mrc1/Csm3 checkpoint complex. J. Mol. Biol. 2005, 347, 509–521. [Google Scholar] [CrossRef]
- Rossi, R.; Villa, A.; Negri, C.; Scovassi, I.; Ciarrocchi, G.; Biamonti, G.; Montecucco, A. The replication factory targeting sequence/PCNA-binding site is required in G(1) to control the phosphorylation status of DNA ligase I. EMBO J. 1999, 18, 5745–5754. [Google Scholar] [CrossRef]
- Norio, P.; Kosiyatrakul, S.; Yang, Q.; Guan, Z.; Brown, N.M.; Thomas, S.; Riblet, R.; Schildkraut, C.L. Progressive activation of DNA replication initiation in large domains of the immunoglobulin heavy chain locus during B cell development. Mol. Cell 2005, 20, 575–587. [Google Scholar] [CrossRef]
- Jackson, D.A.; Pombo, A. Replicon clusters are stable units of chromosome structure: evidence that nuclear organization contributes to the efficient activation and propagation of S phase in human cells. J. Cell Biol. 1998, 140, 1285–1295. [Google Scholar] [CrossRef]
- Kamisuki, S.; Takahashi, S.; Mizushina, Y.; Hanashima, S.; Kuramochi, K.; Kobayashi, S.; Sakaguchi, K.; Nakata, T.; Sugawara, F. Total synthesis of dehydroaltenusin. Tetrahedron 2004, 60, 5695–5700. [Google Scholar] [CrossRef]
- Takahashi, S.; Kamisuki, S.; Mizushina, Y.; Sakaguchi, K.; Sugawar, F.; Nakata, T. Total synthesis of dehydroaltenusin. Tetrahedron Lett. 2003, 44, 1875–1877. [Google Scholar] [CrossRef]
- Kuriyama, I.; Fukudome, K.; Kamisuki, S.; Kuramochi, K.; Tsubaki, K.; Sakaguchi, K.; Sugawara, F.; Yoshida, H.; Mizushina, Y. The specific inhibitory effect of demethoxydehydroaltenusin, a derivative of dehydroaltenusin, on mammalian DNA polymerase α . Int. J. Mol. Med. 2008, 22, 793–799. [Google Scholar]
- Tamai, K.; Kojima, K.; Hanaichi, T.; Masaki, S.; Suzuki, M.; Umekawa, H.; Yoshida, S. Structural study of immunoaffinity-purified DNA polymerase α-DNA primase complex from calf thymus. Biochim Biophys Acta 1988, 950, 263–273. [Google Scholar] [CrossRef]
- Mizushina, Y.; Yoshida, S.; Matsukage, A.; Sakaguchi, K. The inhibitory action of fatty acids on DNA polymerase β. Biochim. Biophys. Acta 1336, 509–521. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Mizuno, T.; Yamagishi, K.; Miyazawa, H.; Hanaoka, F. Molecular architecture of the mouse DNA polymerase α-primase complex. Mol. Cell. Biol. 1999, 19, 7886–7896. [Google Scholar]
- Sample Availability: Samples of the synthesized dehydroaltenusin and its derivatives are available from Dr. Y. Mizushina (Kobe-Gakuin University).
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kuriyama, I.; Mizuno, T.; Fukudome, K.; Kuramochi, K.; Tsubaki, K.; Usui, T.; Imamoto, N.; Sakaguchi, K.; Sugawara, F.; Yoshida, H.; et al. Effect of Dehydroaltenusin-C12 Derivative, a Selective DNA Polymerase α Inhibitor, on DNA Replication in Cultured Cells. Molecules 2008, 13, 2948-2961. https://doi.org/10.3390/molecules13122948
Kuriyama I, Mizuno T, Fukudome K, Kuramochi K, Tsubaki K, Usui T, Imamoto N, Sakaguchi K, Sugawara F, Yoshida H, et al. Effect of Dehydroaltenusin-C12 Derivative, a Selective DNA Polymerase α Inhibitor, on DNA Replication in Cultured Cells. Molecules. 2008; 13(12):2948-2961. https://doi.org/10.3390/molecules13122948
Chicago/Turabian StyleKuriyama, Isoko, Takeshi Mizuno, Keishi Fukudome, Kouji Kuramochi, Kazunori Tsubaki, Takeo Usui, Naoko Imamoto, Kengo Sakaguchi, Fumio Sugawara, Hiromi Yoshida, and et al. 2008. "Effect of Dehydroaltenusin-C12 Derivative, a Selective DNA Polymerase α Inhibitor, on DNA Replication in Cultured Cells" Molecules 13, no. 12: 2948-2961. https://doi.org/10.3390/molecules13122948
APA StyleKuriyama, I., Mizuno, T., Fukudome, K., Kuramochi, K., Tsubaki, K., Usui, T., Imamoto, N., Sakaguchi, K., Sugawara, F., Yoshida, H., & Mizushina, Y. (2008). Effect of Dehydroaltenusin-C12 Derivative, a Selective DNA Polymerase α Inhibitor, on DNA Replication in Cultured Cells. Molecules, 13(12), 2948-2961. https://doi.org/10.3390/molecules13122948