Vitamin B6: A Long Known Compound of Surprising Complexity


Abstract
:Introduction
The Discovery of Vitamin B6
The Function of VitB6
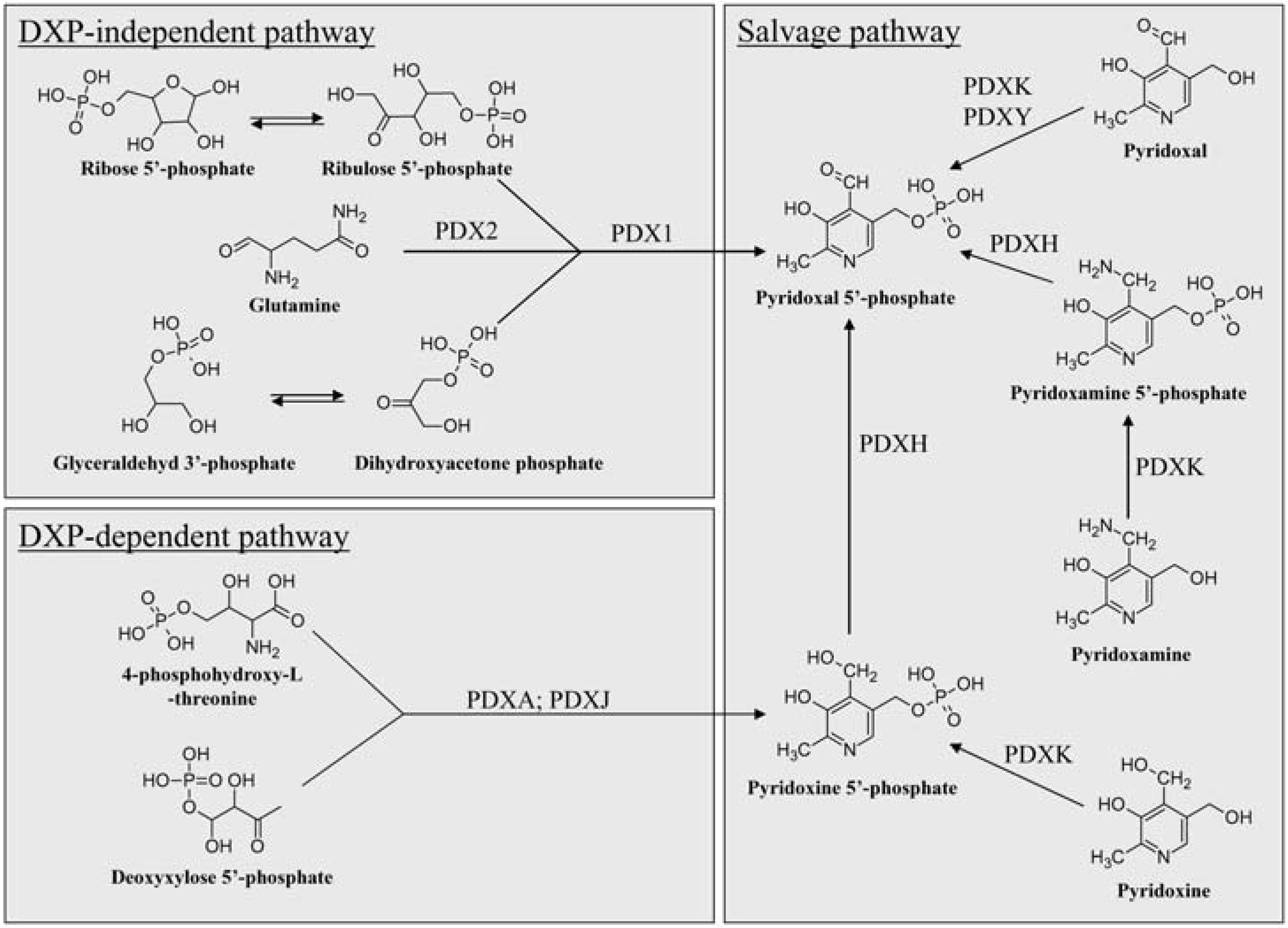
The Known Pathways of VitB6 Anabolism
Deoxyxylose 5’-phosphate-Dependent and –Independent De Novo Biosynthesis of VitB6
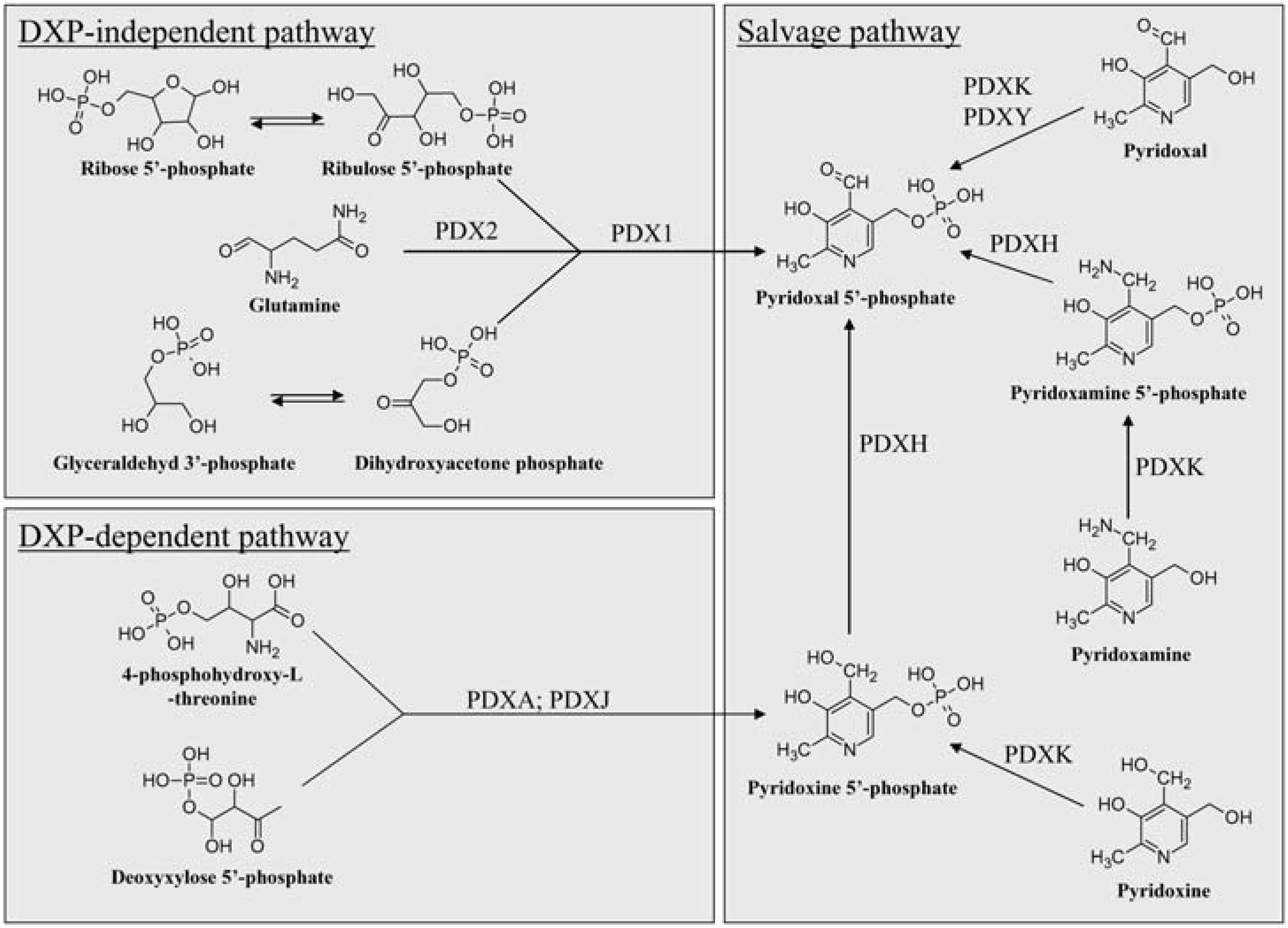
The Salvage Pathway
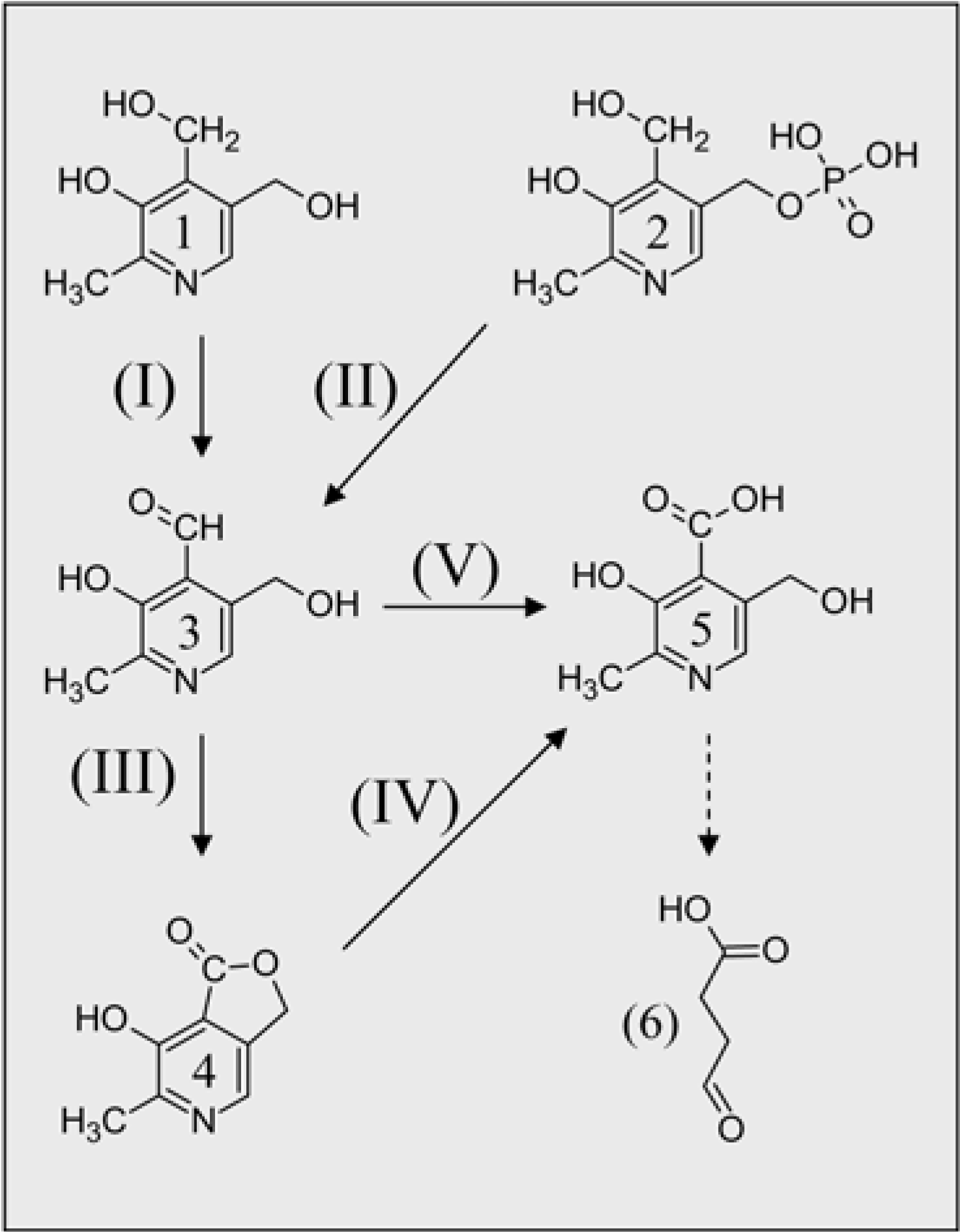
The Other End: VitB6 Catabolism

Allocation of VitB6 in Prokaryotes and Eukaryotes
The Role of VitB6 in Stress Response

{kind=link}
{kind=link}
| Organism | Mutant | Pathway affected | Phenotype | Citation |
|---|---|---|---|---|
| E. coli | ppox/pdxH | Salvage | Reduced growth, aberrant shape | [27] |
| C. nicotianae | sor1/pdx1 | De novo | Increased ROS sensitivity, loss of vitB6 production, increased salt sensitivity, reduced growth | [81] |
| S. cerevisiae | snz1/pdx1 | De novo | Reduced growth in minimal media | [94] |
| S. cerevisiae | sno1/pdx2 | De novo | Reduced growth in minimal media | [94] |
| S. cerevisiae | pdx3 | Salvage | Increased ROS sensitivity | [50] |
| A. thaliana | sos4-1 | Salvage | Increased salt sensitivity | [87, 90] |
| A. thaliana | pdx 1.1, pdx 1.3 | De novo | Increased salt sensitivity | [21, 86] |
| A. thaliana | pdx3/PPOX | Salvage | Reduced aerial & root growth, increased salt sensitivity | [50, 90] |
The Diversity of VitB6 Derivatives
| Derivative | Structure | Function | Organism found | citation |
|---|---|---|---|---|
| Vitamin B6 |  | Antioxidant | ubiquitous | [13] |
| Vitamin B6-phosphate |  | Cofactor | ubiquitous | [13] |
| 4’-O-Methyl-pyridoxine (ginkgotoxin) |  | Unknown; potential inhibitor of PN/PM/PL kinase | Ginkgo biloba, Albizia tanganyicensis | [23, 97, 99] |
| 5’-O-Acetyl-4’-O-methylpyridoxine |  | Unknown | Albizia tanganyicensis | [100] |
| Julibrine I |  | Unknown | Albizia julibrissin | [102] |
| Julibrine II |  | Unknown; has been demonstrated to induce arrhythmia | Albizia julibrissin | [102] |
| 5'-0-(β-D-Glucopyranosyl) pyridoxine |  | Unknown | Various plant products | [111, 112] |
| N-Hexanoyl-pyridoxamine (HAPM) |  | Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [9, 10] |
| N-Nonanedioyl-pyridoxamine monoamide (NDAPM) |  | Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [9, 10] |
| N-Pentanedioyl-pyridoxamine monoamide (PDAPM) |  | Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [104, 105] |
| N-Formyl-pyridoxamine (FAPM) |  | Inhibitor of advanced glycation and lipoxidation reactions | PM treated diabetic and obese rats | [104, 105] |
VitB6 Supplements and Human Health: Is it too much or not enough of a good thing?
Outlook and Perspectives
Acknowledgments
References and Notes
- Ohdake, S. Isolation of "Oryzanin" (Antineuritic Vitamin) from Rice-polishings. Bull. Agri. Chem. Soc. Japan. 1932, 8, 11–46. [Google Scholar] [CrossRef]
- Wiardy, P.W. Crystalline Vitamin B6 (Adermin). Nature 1938, 142, 1158–1158. [Google Scholar] [CrossRef]
- György, P. Vitamin B2 and the Pellagra-like Dermatitis in Rats. Nature 1934, 133, 498–499. [Google Scholar] [CrossRef]
- György, P. Crystalline vitamin B6. J. Am. Chem. Soc. 1938, 60, 983–984. [Google Scholar] [CrossRef]
- Ichiba, A.; Michi, K. Cristalline vitamin B6. Sci. Pap. Inst. Phys. Chem. Res. (Tokyo). 1938, 34, 623–626. [Google Scholar]
- Keresztesy, J.C.; Stevens, J.R. Vitamin B-6. Proc. Soc. Exp. Biol. Med. 1938, 38, 64–65. [Google Scholar] [CrossRef]
- Kuhn, R.; Wendt, G. Über das antidermatitische Vitamin der Hefe. Ber. Chem. Ges. 1938, 71, 780–782. [Google Scholar]
- Lepkovsky. Crystalline factor I. Science 1938, 87, 169–170. [Google Scholar]
- György, P.; Eckardt, R.E. Vitamin B6 and Skin Lesions in Rats. Nature 1939, 144, 512–512. [Google Scholar]
- Harris, S.A.; Folkers, K. Synthetic vitamin B6. Science 1939, 89, 347. [Google Scholar]
- Snell, E.E.; Guirard, B.M.; Williams, R.J. Occurrence in natural products of a physiologically active metabolite of pyridoxine. J. Biol. Chem. 1942, 143, 519–530. [Google Scholar]
- Drewke, C.; Leistner, E. Biosynthesis of vitamin B6 and structurally related derivatives. Vitam. Horm. 2001, 61, 121–155. [Google Scholar]
- Mittenhuber, G. Phylogenetic analyses and comparative genomics of vitamin B6 (pyridoxine) and pyridoxal phosphate biosynthesis pathways. J. Mol. Microbiol.Biotechnol. 2001, 3, 1–20. [Google Scholar]
- Grogan, D.W. Temperature-sensitive murein synthesis in an Escherichia coli pdx mutant and the role of alanine racemase. Arc. Microbiol. 1988, 150, 363–367. [Google Scholar] [CrossRef]
- Nakamura, M.T.; Nara, T.Y. Structure, function, and dietary regulation of delta-6, delta-5, and delta-9 desaturases. Annu. Rev. Nutr. 2004, 24, 345–376. [Google Scholar] [CrossRef]
- Horrobin, D.F. Fatty acid metabolism in health and disease: the role of delta-6-desaturase. Am. J. Clin. Nutr. 1993, 57, 732S-736S; discussion 736S-737S. [Google Scholar]
- Helmreich, E.J. How pyridoxal 5'-phosphate could function in glycogen phosphorylase catalysis. Biofactors 1992, 3, 159–172. [Google Scholar]
- Cheltsov, A.V.; Guida, W.C.; Ferreira, G.C. Circular permutation of 5-aminolevulinate synthase: effect on folding, conformational stability, and structure. J. Biol. Chem. 2003, 278, 27945–27955. [Google Scholar] [CrossRef]
- Rottmann, W.H.; Peter, G.F.; Oeller, P.W.; Keller, J.A.; Shen, N.F.; Nagy, B.P.; Taylor, L.P.; Campbell, A.D.; Theologis, A. 1-aminocyclopropane-1-carboxylate synthase in tomato is encoded by a multigene family whose transcription is induced during fruit and floral senescence. J. Mol. Biol. 1991, 222, 937–961. [Google Scholar] [CrossRef]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (Pyridoxine) and Its Derivatives Are Efficient Singlet Oxygen Quenchers and Potential Fungal Antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Chen, H.; Xiong, L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005, 44, 396–408. [Google Scholar] [CrossRef]
- Hill, R.E.; Himmeldirk, K.; Kennedy, I.A.; Pauloski, R.M.; Sayer, B.G.; Wolf, E.; Spenser, I.D. The biogenetic anatomy of vitamin B6. A 13C NMR investigation of the biosynthesis of pyridoxol in Escherichia coli. J. Biol. Chem. 1996, 271, 30426–30435. [Google Scholar]
- Arenz, A.; Klein, M.; Fiehe, K.; Gross, J.; Drewke, C.; Hemscheidt, T.; Leistner, E. Occurrence of neurotoxic 4'-O-methylpyridoxine in Ginkgo biloba leaves, Ginkgo medications and Japanese Ginkgo food. Planta Med. 1996, 62, 548–551. [Google Scholar] [CrossRef]
- Drewke, C.; Notheis, C.; Hansen, U.; Leistner, E.; Hemscheidt, T.; Hill, R.E.; Spenser, I.D. Growth response to 4-hydroxy-L-threonine of Escherichia coli mutants blocked in vitamin B6 biosynthesis. FEBS Lett. 1993, 318, 125–128. [Google Scholar] [CrossRef]
- Laber, B.; Maurer, W.; Scharf, S.; Stepusin, K.; Schmidt, F.S. Vitamin B6 biosynthesis: formation of pyridoxine 5'-phosphate from 4-(phosphohydroxy)-L-threonine and 1-deoxy-D-xylulose-5-phosphate by PdxA and PdxJ protein. FEBS Lett. 1999, 449, 45–48. [Google Scholar] [CrossRef]
- Cane, D.E.; Hsiung, Y.; Cornish, J.A.; Robinson, J.K.; Spenser, I.D. Biosynthesis of Vitamin B6: The Oxidation of 4-(Phosphohydroxy)-l-threonine by PdxA. J. Am. Chem. Soc. 1998, 120, 1936–1937. [Google Scholar] [CrossRef]
- Lam, H.M.; Winkler, M.E. Characterization of the complex pdxH-tyrS operon of Escherichia coli K-12 and pleiotropic phenotypes caused by pdxH insertion mutations. J. Bacteriol. 1992, 174, 6033–6045. [Google Scholar]
- Zhao, G.; Winkler, M.E. 4-Phospho-hydroxy-L-threonine is an obligatory intermediate in pyridoxal 5'-phosphate coenzyme biosynthesis in Escherichia coli K-12. FEMS Microbiol. Lett. 1996, 135, 275–280. [Google Scholar]
- Sivaraman, J.; Li, Y.; Banks, J.; Cane, D.E.; Matte, A.; Cygler, M. Crystal structure of Escherichia coli PdxA, an enzyme involved in the pyridoxal phosphate biosynthesis pathway. J. Biol. Chem. 2003, 278, 43682–43690. [Google Scholar]
- Notheis, C.; Drewke, C.; Leistner, E. Purification and characterization of the pyridoxol-5'-phosphate: oxygen oxidoreductase (deaminating) from Escherichia coli. Biochim. Biophys. Acta 1995, 1247, 265–271. [Google Scholar] [CrossRef]
- Sprenger, G.A.; Schorken, U.; Wiegert, T.; Grolle, S.; de Graaf, A.A.; Taylor, S.V.; Begley, T.P.; Bringer-Meyer, S.; Sahm, H. Identification of a thiamin-dependent synthase in Escherichia coli required for the formation of the 1-deoxy-D-xylulose 5-phosphate precursor to isoprenoids, thiamin, and pyridoxol. Proc. Natl. Acad. Sci. USA 1997, 94, 12857–12862. [Google Scholar] [CrossRef]
- Franco, M.G.; Laber, B.; Huber, R.; Clausen, T. Structural basis for the function of pyridoxine 5'-phosphate synthase. Structure 2001, 9, 245–253. [Google Scholar] [CrossRef]
- Burns, K.E.; Xiang, Y.; Kinsland, C.L.; McLafferty, F.W.; Begley, T.P. Reconstitution and biochemical characterization of a new pyridoxal-5'-phosphate biosynthetic pathway. J. Am. Chem. Soc. 2005, 127, 3682–3683. [Google Scholar] [CrossRef]
- Tambasco-Studart, M.; Titiz, O.; Raschle, T.; Forster, G.; Amrhein, N.; Fitzpatrick, T.B. Vitamin B6 biosynthesis in higher plants. Proc. Natl. Acad. Sci. USA 2005, 102, 13687–13692. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Bilski, P.; Li, M.Y.; Chignell, C.F.; Daub, M.E. A highly conserved sequence is a novel gene involved in de novo vitamin B6 biosynthesis. Proc. Natl. Acad. Sci. USA 1999, 96, 9374–9378. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Fitzpatrick, T.B.; Raschle, T.; Flicker, K.; Sinning, I.; Muller, S.; Macheroux, P.; Tews, I.; Kappes, B. Vitamin B6 biosynthesis by the malaria parasite Plasmodium falciparum: biochemical and structural insights. J. Biol. Chem. 2006, 281, 3633–3641. [Google Scholar]
- Zhu, J.; Burgner, J.W.; Harms, E.; Belitsky, B.R.; Smith, J.L. A new arrangement of (beta/alpha)8 barrels in the synthase subunit of PLP synthase. J. Biol. Chem. 2005, 280, 27914–27923. [Google Scholar]
- Tambasco-Studart, M.; Tews, I.; Amrhein, N.; Fitzpatrick, T.B. Functional analysis of PDX2 from Arabidopsis, a glutaminase involved in vitamin B6 biosynthesis. Plant Physiol. 2007, 144, 915–925. [Google Scholar] [CrossRef]
- Zein, F.; Zhang, Y.; Kang, Y.N.; Burns, K.; Begley, T.P.; Ealick, S.E. Structural insights into the mechanism of the PLP synthase holoenzyme from Thermotoga maritima. Biochemistry 2006, 45, 14609–14620. [Google Scholar] [CrossRef]
- Flicker, K.; Neuwirth, M.; Strohmeier, M.; Kappes, B.; Tews, I.; Macheroux, P. Structural and thermodynamic insights into the assembly of the heteromeric pyridoxal phosphate synthase from Plasmodium falciparum. J. Mol. Biol. 2007, 374, 732–748. [Google Scholar] [CrossRef]
- Yang, Y.; Tsui, H.C.; Man, T.K.; Winkler, M.E. Identification and function of the pdxY gene, which encodes a novel pyridoxal kinase involved in the salvage pathway of pyridoxal 5'-phosphate biosynthesis in Escherichia coli K-12. J. Bacteriol. 1998, 180, 1814–1821. [Google Scholar]
- Yang, Y.; Zhao, G.; Winkler, M.E. Identification of the pdxK gene that encodes pyridoxine (vitamin B6) kinase in Escherichia coli K-12. FEMS Microbiol. Lett. 1996, 141, 89–95. [Google Scholar] [CrossRef]
- Cao, P.; Gong, Y.; Tang, L.; Leung, Y.C.; Jiang, T. Crystal structure of human pyridoxal kinase. J. Struct. Biol. 2006, 154, 327–332. [Google Scholar] [CrossRef]
- Li, M.H.; Kwok, F.; Chang, W.R.; Lau, C.K.; Zhang, J.P.; Lo, S.C.; Jiang, T.; Liang, D.C. Crystal structure of brain pyridoxal kinase, a novel member of the ribokinase superfamily. J. Biol. Chem. 2002, 277, 46385–46390. [Google Scholar]
- Safo, M.K.; Musayev, F.N.; di Salvo, M.L.; Hunt, S.; Claude, J.B.; Schirch, V. Crystal structure of pyridoxal kinase from the Escherichia coli pdxK gene: implications for the classification of pyridoxal kinases. J. Bacteriol. 2006, 188, 4542–4552. [Google Scholar] [CrossRef]
- Safo, M.K.; Musayev, F.N.; Hunt, S.; di Salvo, M.L.; Scarsdale, N.; Schirch, V. Crystal structure of the PdxY Protein from Escherichia coli. J. Bacteriol. 2004, 186, 8074–8082. [Google Scholar] [CrossRef]
- Musayev, F.N.; di Salvo, M.L.; Ko, T.P.; Gandhi, A.K.; Goswami, A.; Schirch, V.; Safo, M.K. Crystal Structure of human pyridoxal kinase: structural basis of M(+) and M(2+) activation. Protein Sci. 2007, 16, 2184–2194. [Google Scholar] [CrossRef]
- Zhao, G.; Winkler, M.E. Kinetic limitation and cellular amount of pyridoxine (pyridoxamine) 5'-phosphate oxidase of Escherichia coli K-12. J. Bacteriol. 1995, 177, 883–891. [Google Scholar]
- Loubbardi, A.; Marcireau, C.; Karst, F.; Guilloton, M. Sterol uptake induced by an impairment of pyridoxal phosphate synthesis in Saccharomyces cerevisiae: cloning and sequencing of the PDX3 gene encoding pyridoxine (pyridoxamine) phosphate oxidase. J. Bacteriol. 1995, 177, 1817–1823. [Google Scholar]
- Sang, Y.; Barbosa, J.M.; Wu, H.; Locy, R.D.; Singh, N.K. Identification of a pyridoxine (pyridoxamine) 5'-phosphate oxidase from Arabidopsis thaliana. FEBS Lett. 2007, 581, 344–348. [Google Scholar] [CrossRef]
- Pedelacq, J.D.; Rho, B.S.; Kim, C.Y.; Waldo, G.S.; Lekin, T.P.; Segelke, B.W.; Rupp, B.; Hung, L.W.; Kim, S.I.; Terwilliger, T.C. Crystal structure of a putative pyridoxine 5'-phosphate oxidase (Rv2607) from Mycobacterium tuberculosis. Proteins 2006, 62, 563–569. [Google Scholar]
- Biswal, B.K.; Au, K.; Cherney, M.M.; Garen, C.; James, M.N. The molecular structure of Rv2074, a probable pyridoxine 5'-phosphate oxidase from Mycobacterium tuberculosis, at 1.6 angstroms resolution. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 735–742. [Google Scholar] [CrossRef]
- di Salvo, M.L.; Safo, M.K.; Musayev, F.N.; Bossa, F.; Schirch, V. Structure and mechanism of Escherichia coli pyridoxine 5'-phosphate oxidase. Biochim. Biophys. Acta 2003, 1647, 76–82. [Google Scholar]
- Fonda, M.L.; Trauss, C.; Guempel, U.M. The binding of pyridoxal 5'-phosphate to human serum albumin. Arch. Biochem. Biophys. 1991, 288, 79–86. [Google Scholar] [CrossRef]
- Li, T.K.; Lumeng, L.; Veitch, R.L. Regulation of pyridoxal 5'-phosphate metabolism in liver. Biochem. Biophys. Res. Commun. 1974, 61, 677–684. [Google Scholar] [CrossRef]
- Lumeng, L.; Brashear, R.E.; Li, T.K. Pyridoxal 5'-phosphate in plasma: source, protein-binding, and cellular transport. J. Lab. Clin. Med. 1974, 84, 334–343. [Google Scholar]
- Harris, H. The human alkaline phosphatases: what we know and what we don't know. Clin. Chim. Acta 1990, 186, 133–150. [Google Scholar] [CrossRef]
- Bull, H.; Murray, P.G.; Thomas, D.; Fraser, A.M.; Nelson, P.N. Acid phosphatases. Mol. Pathol. 2002, 55, 65–72. [Google Scholar] [CrossRef]
- Lee, Y.P.; Kim, D.W.; Lee, M.J.; Jeong, M.S.; Kim, S.Y.; Lee, S.H.; Jang, S.H.; Park, J.; Kang, T.C.; Won, M.H.; Cho, S.W.; Kwon, O.S.; Eum, W.S.; Choi, S.Y. Human brain pyridoxal-5'-phosphate phosphatase (PLPP):protein transduction of PEP-1-PLPP into PC12 cells. BMB Rep. 2008, 41, 408–413. [Google Scholar] [CrossRef]
- Jang, Y.M.; Kim, D.W.; Kang, T.C.; Won, M.H.; Baek, N.I.; Moon, B.J.; Choi, S.Y.; Kwon, O.S. Human pyridoxal phosphatase. Molecular cloning, functional expression, and tissue distribution. J. Biol. Chem. 2003, 278, 50040–50046. [Google Scholar]
- Fonda, M.L. Purification and characterization of vitamin B6-phosphate phosphatase from human erythrocytes. J. Biol. Chem. 1992, 267, 15978–15983. [Google Scholar]
- Mukherjee, T.; Kinsland, C.; Begley, T.P. PLP catabolism: identification of the 4-pyridoxic acid dehydrogenase gene in Mesorhizobium loti MAFF303099. Bioorg. Chem. 2007, 35, 458–464. [Google Scholar] [CrossRef]
- Yagi, T.; Kishore, G.M.; Snell, E.E. The bacterial oxidation of vitamin B6. 4-Pyridoxic acid dehydrogenase: a membrane-bound enzyme from Pseudomonas MA-1. J. Biol. Chem. 1983, 258, 9419–9425. [Google Scholar]
- Burg, R.W.; Rodwell, V.W.; Snell, E.E. Bacterial oxidation of vitamin B6. II. Metabolites of pyridoxamine. J. Biol. Chem. 1960, 235, 1164–1169. [Google Scholar]
- Fukuwatari, T.; Wada, H.; Shibata, K. Age-related alterations of B-group vitamin contents in urine, blood and liver from rats. J. Nutr. Sci. Vitaminol. (Tokyo) 2008, 54, 357–362. [Google Scholar] [CrossRef]
- Rybak, M.E.; Pfeiffer, C.M. Clinical analysis of vitamin B(6): determination of pyridoxal 5'-phosphate and 4-pyridoxic acid in human serum by reversed-phase high-performance liquid chromatography with chlorite postcolumn derivatization. Anal. Biochem. 2004, 333, 336–344. [Google Scholar] [CrossRef]
- Pearl, P.L.; Novotny, E.J.; Acosta, M.T.; Jakobs, C.; Gibson, K.M. Succinic semialdehyde dehydrogenase deficiency in children and adults. Ann. Neurol. 2003, 54 Suppl. 6, S73–S80. [Google Scholar]
- Mulligan, J.H.; Snell, E.E. Transport and metabolism of vitamin B6 in Salmonella typhimurium LT2. J. Biol. Chem. 1976, 251, 1052–1056. [Google Scholar]
- Shane, B.; Snell, E.E. Transport and metabolism of vitamin B6 in the yeast Saccharomyces carlsbergensis 4228. J. Biol. Chem. 1976, 251, 1042–1051. [Google Scholar]
- Zhang, Z.M.; McCormick, D.B. Uptake of N-(4'-pyridoxyl) amines and release of amines by renal cells: a model for transporter-enhanced delivery of bioactive compounds. Proc. Natl. Acad. Sci. USA 1991, 88, 10407–10410. [Google Scholar] [CrossRef]
- Said, Z.M.; Subramanian, V.S.; Vaziri, N.D.; ad Said, H.M. Pyridoxine uptake by colonocytes: a specific and regulated carrier-mediated process. Am. J. Physiol. Cell. Physiol. 2008, 294, C1192–C1197. [Google Scholar] [CrossRef]
- Said, H.M.; Ortiz, A.; Ma, T.Y. A carrier-mediated mechanism for pyridoxine uptake by human intestinal epithelial Caco-2 cells: regulation by a PKA-mediated pathway. Am. J. Physiol. Cell. Physiol. 2003, 285, C1219–C1225. [Google Scholar] [CrossRef]
- Schenker, S.; Johnson, R.F.; Mahuren, J.D.; Henderson, G.I.; Coburn, S.P. Human placental vitamin B6 (pyridoxal) transport: normal characteristics and effects of ethanol. Am. J. Physiol. 1992, 262, R966–R974. [Google Scholar]
- Hirose, K.; Chumnantana, R.; Nakashima, T.; Ashiuchi, M.; Yagi, T. Efflux system for pyridoxine in Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem. 2000, 64, 2675–2679. [Google Scholar] [CrossRef]
- Yagi, T.; Tanouchi, A.; Hiraoka, Y. Growth phase-dependent active transport of pyridoxine in a fission yeast, Schizosaccharomyces pombe. FEMS Microbiol. Lett. 1998, 161, 145–150. [Google Scholar] [CrossRef]
- Morita, T.; Takegawa, K.; Yagi, T. Disruption of the plr1+ gene encoding pyridoxal reductase of Schizosaccharomyces pombe. J. Biochem. 2004, 135, 225–230. [Google Scholar] [CrossRef]
- Stolz, J.; Vielreicher, M. Tpn1p, the plasma membrane vitamin B6 transporter of Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 18990–18996. [Google Scholar] [CrossRef]
- Stolz, J.; Wohrmann, H.J.; Vogl, C. Amiloride uptake and toxicity in fission yeast are caused by the pyridoxine transporter encoded by bsu1+ (car1+). Eukaryot. Cell. 2005, 4, 319–326. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Hebbeln, P.; Eudes, A.; Ter Beek, J.; Rodionova, I.A.; Erkens, G.B.; Slotboom, D.J.; Gelfand, M.S.; Osterman, A.L.; Hanson, A.D.; Eitinger, T. A Novel Class of Modular Transporters for Vitamins in Prokaryotes. J. Bacteriol. 2009, 191, 42–51. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Jenns, A.E.; Chung, K.R.; Daub, M.E. SOR1, a gene required for photosensitizer and singlet oxygen resistance in Cercospora fungi, is highly conserved in divergent organisms. Mol. Cell. 1998, 1, 603–609. [Google Scholar] [CrossRef]
- Ehrenshaft, M.; Chung, K.R.; Jenns, A.E.; Daub, M.E. Functional characterization of SOR1, a gene required for resistance to photosensitizing toxins in the fungus Cercospora nicotianae. Curr. Genet. 1999, 34, 478–485. [Google Scholar] [CrossRef]
- Bilski, P.; Li, M.Y.; Ehrenshaft, M.; Daub, M.E.; Chignell, C.F. Vitamin B6 (pyridoxine) and its derivatives are efficient singlet oxygen quenchers and potential fungal antioxidants. Photochem. Photobiol. 2000, 71, 129–134. [Google Scholar] [CrossRef]
- Bilski, P.; Daub, M.E.; Chignell, C.F. Direct detection of singlet oxygen via its phosphorescence from cellular and fungal cultures. Meth. Enzymol. 2002, 352, 41–52. [Google Scholar]
- Graham, C.M.; Ehrenshaft, M.; Hausner, G.; Reid, D.M. A highly conserved gene for vitamin B biosynthesis may have consequences for stress and hormone responses in plants. Physiol. Plant. 2004, 121, 8–14. [Google Scholar] [CrossRef]
- Chen, H.; Xiong, L. Pyridoxine is required for post-embryonic root development and tolerance to osmotic and oxidative stresses. Plant J. 2005, 44, 396–408. [Google Scholar] [CrossRef]
- Titiz, O.; Tambasco-Studart, M.; Warzych, E.; Apel, K.; Amrhein, N.; Laloi, C.; Fitzpatrick, T.B. PDX1 is essential for vitamin B6 biosynthesis, development and stress tolerance in Arabidopsis. Plant J. 2006, 48, 933–946. [Google Scholar] [CrossRef]
- Shi, H.; Xiong, L.; Stevenson, B.; Lu, T.; Zhu, J.K. The Arabidopsis salt overly sensitive 4 mutants uncover a critical role for vitamin B6 in plant salt tolerance. Plant Cell 2002, 14, 575–588. [Google Scholar] [CrossRef]
- Wagner, S.; Bernhardt, A.; Leuendorf, J.E.; Drewke, C.; Lytovchenko, A.; Mujahed, N.; Gurgui, C.; Frommer, W.B.; Leistner, E.; Fernie, A.R.; Hellmann, H. Analysis of the Arabidopsis rsr4-1/pdx1-3 mutant reveals the critical function of the PDX1 protein family in metabolism, development, and vitamin B6 biosynthesis. Plant Cell 2006, 18, 1722–1735. [Google Scholar] [CrossRef]
- Leuendorf, J.E.; Genau, A.; Szewczyk, A.; Mooney, S.; Drewke, C.; Leistner, E.; Hellmann, H. The Pdx1 family is structurally and functionally conserved between Arabidopsis thaliana and Ginkgo biloba. Febs J. 2008, 275, 960–969. [Google Scholar] [CrossRef]
- Gonzalez, E.; Danehower, D.; Daub, M.E. Vitamer levels, stress response, enzyme activity, and gene regulation of Arabidopsis lines mutant in the pyridoxine/pyridoxamine 5'-phosphate oxidase (PDX3) and the pyridoxal kinase (SOS4) genes involved in the vitamin B6 salvage pathway. Plant Physiol. 2007, 145, 985–996. [Google Scholar] [CrossRef]
- Antelmann, H.; Bernhardt, J.; Schmid, R.; Mach, H.; Volker, U.; Hecker, M. First steps from a two-dimensional protein index towards a response-regulation map for Bacillus subtilis. Electrophoresis 1997, 18, 1451–1463. [Google Scholar] [CrossRef]
- Savenstrand, H.; Brosche, M.; Strid, A. Ultraviolet-B signalling: Arabidopsis brassinosteroid mutants are defective in UV-B regulated defence gene expression. Plant Physiol. Biochem. 2004, 42, 687–694. [Google Scholar] [CrossRef]
- Osmani, A.H.; May, G.S.; Osmani, S.A. The extremely conserved pyroA gene of Aspergillus nidulans is required for pyridoxine synthesis and is required indirectly for resistance to photosensitizers. J. Biol. Chem. 1999, 274, 23565–23569. [Google Scholar] [CrossRef]
- Padilla, P.A.; Fuge, E.K.; Crawford, M.E.; Errett, A.; Werner-Washburne, M. The highly conserved, coregulated SNO and SNZ gene families in Saccharomyces cerevisiae respond to nutrient limitation. J. Bacteriol. 1998, 180, 5718–5726. [Google Scholar]
- Braun, E.L.; Fuge, E.K.; Padilla, P.A.; Werner-Washburne, M. A stationary-phase gene in Saccharomyces cerevisiae is a member of a novel, highly conserved gene family. J. Bacteriol. 1996, 178, 6865–6872. [Google Scholar]
- Denslow, S.A.; Rueschhoff, E.E.; Daub, M.E. Regulation of the Arabidopsis thaliana vitamin B6 biosynthesis genes by abiotic stress. Plant Physiol. Biochem. 2007, 45, 152–161. [Google Scholar] [CrossRef]
- Kästner, U.; Hallmen, C.; Wiese, M.; Leistner, E.; Drewke, C. The human pyridoxal kinase, a plausible target for ginkgotoxin from Ginkgo biloba. Febs J. 2007, 274, 1036–1045. [Google Scholar] [CrossRef]
- Arenz, A.; Klein, M.; Fiehe, K.; Gross, J.; Drewke, C.; Hemscheidt, T.; Leistner, E. Occurrence of Neurotoxic 4'-O-Methylpyridoxine in Ginkgo biloba Leaves, Ginkgo Medications and Japanese Ginkgo Food. Planta Med. 1996, 62, 548–551. [Google Scholar] [CrossRef]
- Fiehe, K.; Arenz, A.; Drewke, C.; Hemscheidt, T.; Williamson, R.T.; Leistner, E. Biosynthesis of 4'-O-methylpyridoxine (Ginkgotoxin) from primary precursors. J. Nat. Prod. 2000, 63, 185–189. [Google Scholar] [CrossRef]
- Wada, K.; Ishigaki, S.; Ueda, K.; Sakata, M.; Haga, M. An antivitamin B6, 4'-methoxypyridoxine, from the seed of Ginkgo biloba L. Chem. Pharm. Bull. (Tokyo) 1985, 33, 3555–3557. [Google Scholar]
- Buss, K.; Drewke, C.; Lohmann, S.; Piwonska, A.; Leistner, E. Properties and interaction of heterologously expressed glutamate decarboxylase isoenzymes GAD(65kDa) and GAD(67kDa) from human brain with ginkgotoxin and its 5'-phosphate. J. Med. Chem. 2001, 44, 3166–3174. [Google Scholar] [CrossRef]
- Higuchi, O.; Nakagawa, K.; Tsuzuki, T.; Suzuki, T.; Oikawa, S.; Miyazawa, T. Aminophospholipid glycation and its inhibitor screening system: a new role of pyridoxal 5'-phosphate as the inhibitor. J. Lipid Res. 2006, 47, 964–974. [Google Scholar] [CrossRef]
- Negre-Salvayre, A.; Coatrieux, C.; Ingueneau, C.; Salvayre, R. Advanced lipid peroxidation end products in oxidative damage to proteins. Potential role in diseases and therapeutic prospects for the inhibitors. Br. J. Pharmacol. 2008, 153, 6–20. [Google Scholar]
- Metz, T.O.; Alderson, N.L.; Thorpe, S.R.; Baynes, J.W. Pyridoxamine, an inhibitor of advanced glycation and lipoxidation reactions: a novel therapy for treatment of diabetic complications. Arch. Biochem. Biophys. 2003, 419, 41–49. [Google Scholar] [CrossRef]
- Metz, T.O.; Alderson, N.L.; Chachich, M.E.; Thorpe, S.R.; Baynes, J.W. Pyridoxamine traps intermediates in lipid peroxidation reactions in vivo: evidence on the role of lipids in chemical modification of protein and development of diabetic complications. J. Biol. Chem. 2003, 278, 42012–42019. [Google Scholar]
- Voziyan, P.A.; Metz, T.O.; Baynes, J.W.; Hudson, B.G. A post-Amadori inhibitor pyridoxamine also inhibits chemical modification of proteins by scavenging carbonyl intermediates of carbohydrate and lipid degradation. J. Biol. Chem. 2002, 277, 3397–3403. [Google Scholar] [CrossRef]
- Booth, A.A.; Khalifah, R.G.; Todd, P.; Hudson, B.G. In vitro kinetic studies of formation of antigenic advanced glycation end products (AGEs). Novel inhibition of post-Amadori glycation pathways. J. Biol. Chem. 1997, 272, 5430–5437. [Google Scholar] [CrossRef]
- Booth, A.A.; Khalifah, R.G.; Hudson, B.G. Thiamine pyrophosphate and pyridoxamine inhibit the formation of antigenic advanced glycation end-products: comparison with aminoguanidine. Biochem. Biophys. Res. Commun. 1996, 220, 113–119. [Google Scholar] [CrossRef]
- Mackey, A.D.; McMahon, R.J.; Townsend, J.H.; Gregory, J.F., 3rd. Uptake, hydrolysis, and metabolism of pyridoxine-5'-beta-D-glucoside in Caco-2 cells. J. Nutr. 2004, 134, 842–846. [Google Scholar]
- Gregory, J.F.; Ink, S.L. Identification and quantification of pyridoxine-β-glucoside as a major form of vitamin B-6 in plant-derived foods. J. Agric. Food Chem. 1987, 35, 76–82. [Google Scholar] [CrossRef]
- Opassiri, R.; Hua, Y.; Wara-Aswapati, O.; Akiyama, T.; Svasti, J.; Esen, A.; Ketudat Cairns, J.R. Beta-glucosidase, exo-beta-glucanase and pyridoxine transglucosylase activities of rice BGlu1. Biochem. J. 2004, 379, 125–131. [Google Scholar] [CrossRef]
- Suzuki, Y.; Uchida, K. Formation of 5'-0-(β-D-Glucopyranosyl) pyridoxine in soybean seedlings and suspension cells cultured in the presence of pyridoxine. Bull. Res. Inst. Bioresour. Okayama Univ. 1998, 5, 107–120. [Google Scholar]
- Scott, P.M.; Lau, B.P.; Lawrence, G.A.; Lewis, D.A. Analysis of Ginkgo biloba for the presence of ginkgotoxin and ginkgotoxin 5'-glucoside. J. AOAC Int. 2000, 83, 1313–1320. [Google Scholar]
- Suzuki, Y.; Uchida, K. Formation of beta-galactosides of pyridoxine using Sporobolomyces singularis. Methods Enzymol. 1997, 280, 71–77. [Google Scholar]
- Suzuki, Y.; Uchida, K. Formation of beta-fructosyl compounds of pyridoxine in growing culture of Aspergillus niger. Biosci. Biotechnol. Biochem. 1993, 57, 875–880. [Google Scholar] [CrossRef]
- Opassiri, R.; Pomthong, B.; Onkoksoong, T.; Akiyama, T.; Esen, A.; Ketudat Cairns, J.R. Analysis of rice glycosyl hydrolase family 1 and expression of Os4bglu12 beta-glucosidase. BMC Plant Biol. 2006, 6, 33. [Google Scholar] [CrossRef]
- Nakano, H.; Gregory, J.F., 3rd. Pyridoxine and pyridoxine-5'-beta-D-glucoside exert different effects on tissue B-6 vitamers but similar effects on beta-glucosidase activity in rats. J. Nutr. 1995, 125, 2751–2762. [Google Scholar]
- Trumbo, P.R.; Banks, M.A.; Gregory, J.F., 3rd. Hydrolysis of pyridoxine-5'-beta-D-glucoside by a broad-specificity beta-glucosidase from mammalian tissues. Proc. Soc. Exp. Biol. Med. 1990, 195, 240–246. [Google Scholar] [CrossRef]
- Perry, T.; Holloway, H.W.; Weerasuriya, A.; Mouton, P.R.; Duffy, K.; Mattison, J.A.; Greig, N.H. Evidence of GLP-1-mediated neuroprotection in an animal model of pyridoxine-induced peripheral sensory neuropathy. Exp. Neurol. 2007, 203, 293–301. [Google Scholar] [CrossRef]
- Albin, R.L.; Albers, J.W.; Greenberg, H.S.; Townsend, J.B.; Lynn, R.B.; Burke, J.M., Jr.; Alessi, A.G. Acute sensory neuropathy-neuronopathy from pyridoxine overdose. Neurology 1987, 37, 1729–1732. [Google Scholar] [CrossRef]
- Lu, T.; Xu, Y.; Monttinen, E.S.; Kato, N. Supplementing vitamin B6 to a low vitamin B6 diet exaggerates UVB-Induced skin tumorigenesis in DMBA-treated hairless mice. J. Nutr. Sci. Vitaminol. (Tokyo) 2008, 54, 262–265. [Google Scholar] [CrossRef]
- Wondrak, G.T.; Roberts, M.J.; Jacobson, M.K.; Jacobson, E.L. 3-hydroxypyridine chromophores are endogenous sensitizers of photooxidative stress in human skin cells. J. Biol. Chem. 2004, 279, 30009–30020. [Google Scholar]
- Maeda, T.; Taguchi, H.; Minami, H.; Sato, K.; Shiga, T.; Kosaka, H.; Yoshikawa, K. Vitamin B6 phototoxicity induced by UVA radiation. Arch. Dermatol. Res. 2000, 292, 562–567. [Google Scholar] [CrossRef]
- Theodoratou, E.; Farrington, S.M.; Tenesa, A.; McNeill, G.; Cetnarskyj, R.; Barnetson, R.A.; Porteous, M.E.; Dunlop, M.G.; Campbell, H. Dietary vitamin B6 intake and the risk of colorectal cancer. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 171–182. [Google Scholar] [CrossRef]
- Tsao, S.M.; Yin, M.C.; Liu, W.H. Oxidant stress and B vitamins status in patients with non-small cell lung cancer. Nutr. Cancer. 2007, 59, 8–13. [Google Scholar] [CrossRef]
- van Uffelen, J.G.; Chinapaw, M.J.; van Mechelen, W.; Hopman-Rock, M. Walking or vitamin B for cognition in older adults with mild cognitive impairment? A randomised controlled trial. Br. J. Sports Med. 2008, 42, 344–351. [Google Scholar] [CrossRef] [Green Version]
- van Uffelen, J.G.; Chin, A.P.M.J.; Hopman-Rock, M.; van Mechelen, W. The effect of walking and vitamin B supplementation on quality of life in community-dwelling adults with mild cognitive impairment: a randomized, controlled trial. Qual.Life Res. 2007, 16, 1137–1146. [Google Scholar]
- Luchsinger, J.A.; Tang, M.X.; Miller, J.; Green, R.; Mayeux, R. Relation of higher folate intake to lower risk of Alzheimer disease in the elderly. Arch. Neurol. 2007, 64, 86–92. [Google Scholar] [CrossRef]
- Balk, E.M.; Raman, G.; Tatsioni, A.; Chung, M.; Lau, J.; Rosenberg, I.H. Vitamin B6, B12, and folic acid supplementation and cognitive function: a systematic review of randomized trials. Arch. Intern. Med. 2007, 167, 21–30. [Google Scholar] [CrossRef]
- Ford, A.H.; Flicker, L.; Thomas, J.; Norman, P.; Jamrozik, K.; Almeida, O.P. Vitamins B12, B6, and folic acid for onset of depressive symptoms in older men: results from a 2-year placebo-controlled randomized trial. J. Clin. Psychiatry 2008, 69, 1203–1209. [Google Scholar] [CrossRef]
- Merete, C.; Falcon, L.M.; Tucker, K.L. Vitamin B6 is associated with depressive symptomatology in Massachusetts elders. J. Am. Coll. Nutr. 2008, 27, 421–427. [Google Scholar] [CrossRef]
- Miodownik, C.; Meoded, A.; Libov, I.; Bersudsky, Y.; Sela, B.A.; Lerner, V. Pyridoxal plasma level in schizophrenic and schizoaffective patients with and without tardive dyskinesia. Clin. Neuropharmacol. 2008, 31, 197–203. [Google Scholar] [CrossRef]
- Lerner, V.; Miodownik, C.; Kaptsan, A.; Bersudsky, Y.; Libov, I.; Sela, B.A.; Witztum, E. Vitamin B6 treatment for tardive dyskinesia: a randomized, double-blind, placebo-controlled, crossover study. J. Clin. Psychiatry 2007, 68, 1648–1654. [Google Scholar] [CrossRef]
- Miodownik, C.; Lerner, V.; Statsenko, N.; Dwolatzky, T.; Nemets, B.; Berzak, E.; Bergman, J. Vitamin B6 versus mianserin and placebo in acute neuroleptic-induced akathisia: a randomized, double-blind, controlled study. Clin. Neuropharmacol. 2006, 29, 68–72. [Google Scholar] [CrossRef]
- Rimland, B.; Edelson, S.M. Parent ratings of behavior effects of biomedical interventions; Autism Research Institute: San Diego, 2005; p. 34. [Google Scholar]
- Audhya, T. Laboratory indices of vitamin and mineral deficiency in autism. In Presented at Defeat Autism Now! San Diego, California, USA, 2002.
- Lonn, E.; Yusuf, S.; Arnold, M.J.; Sheridan, P.; Pogue, J.; Micks, M.; McQueen, M.J.; Probstfield, J.; Fodor, G.; Held, C.; Genest, J., Jr. Homocysteine lowering with folic acid and B vitamins in vascular disease. N. Engl. J. Med. 2006, 354, 1567–1577. [Google Scholar] [CrossRef]
- Ishihara, J.; Iso, H.; Inoue, M.; Iwasaki, M.; Okada, K.; Kita, Y.; Kokubo, Y.; Okayama, A.; Tsugane, S. Intake of folate, vitamin B6 and vitamin B12 and the risk of CHD: the Japan Public Health Center-Based Prospective Study Cohort I. J. Am. Coll. Nutr. 2008, 27, 127–136. [Google Scholar] [CrossRef]
- Albert, C.M.; Cook, N.R.; Gaziano, J.M.; Zaharris, E.; MacFadyen, J.; Danielson, E.; Buring, J.E.; Manson, J.E. Effect of folic acid and B vitamins on risk of cardiovascular events and total mortality among women at high risk for cardiovascular disease: a randomized trial. JAMA 2008, 299, 2027–2036. [Google Scholar] [CrossRef]
- Herrmann, M.; Schmidt, J.; Umanskaya, N.; Colaianni, G.; Al Marrawi, F.; Widmann, T.; Zallone, A.; Wildemann, B.; Herrmann, W. Stimulation of osteoclast activity by low B-vitamin concentrations. Bone 2007, 41, 584–591. [Google Scholar] [CrossRef]
- Toyota, T.; Kai, Y.; Kakizaki, M.; Ohtsuka, H.; Shibata, Y.; Goto, Y. The endocrine pancreas in pyridoxine deficient rats. Tohoku J. Exp. Med. 1981, 134, 331–336. [Google Scholar] [CrossRef]
- Jain, S.K. Vitamin B6 (pyridoxamine) supplementation and complications of diabetes. Metabolism 2007, 56, 168–171. [Google Scholar] [CrossRef]
- MacKenzie, K.E.; Wiltshire, E.J.; Gent, R.; Hirte, C.; Piotto, L.; Couper, J.J. Folate and vitamin B6 rapidly normalize endothelial dysfunction in children with type 1 diabetes mellitus. Pediatrics 2006, 118, 242–253. [Google Scholar] [CrossRef]
- Denslow, S.A.; Rueschhoff, E.E.; Daub, M.E. Regulation of the Arabidopsis thaliana vitamin B(6) biosynthesis genes by abiotic stress. Plant Physiol. Biochem. 2007, 2, 152–161. [Google Scholar] [CrossRef]
- Musayev, F.N.; Di Salvo, M.L.; Ko, T.P.; Schirch, V.; Safo, M.K. Structure and properties of recombinant human pyridoxine 5'-phosphate oxidase. Protein Sci. 2003, 12, 1455–1463. [Google Scholar] [CrossRef]
- Zhao, G.; Winkler, M.E. Kinetic limitation and cellular amount of pyridoxine (pyridoxamine) 5'-phosphate oxidase of Escherichia coli K-12. J. Bacteriol. 1995, 177, 883–891. [Google Scholar]
- Laine-Cessac, P.; Cailleux, A.; Allain, P. Mechanisms of the inhibition of human erythrocyte pyridoxal kinase by drugs. Biochem. Pharmacol. 1997, 54, 863–870. [Google Scholar]
- Hanna, M.C.; Turner, A.J.; Kirkness, E.F. Human pyridoxal kinase. cDNA cloning, expression, and modulation by ligands of the benzodiazepine receptor. J. Biol. Chem. 1997, 272, 10756–10760. [Google Scholar] [CrossRef]
- Li, M.H.; Kwok, F.; An, X.M.; Chang, W.R.; Lau, C.K.; Zhang, J.P.; Liu, S.Q.; Leung, Y.C.; Jiang, T.; Liang, D.C. Crystallization and preliminary crystallographic studies of pyridoxal kinase from sheep brain. Acta Crystallogr. D. Biol. Crystallogr. 2002, 58, 1479–1481. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mooney, S.; Leuendorf, J.-E.; Hendrickson, C.; Hellmann, H. Vitamin B6: A Long Known Compound of Surprising Complexity. Molecules 2009, 14, 329-351. https://doi.org/10.3390/molecules14010329
Mooney S, Leuendorf J-E, Hendrickson C, Hellmann H. Vitamin B6: A Long Known Compound of Surprising Complexity. Molecules. 2009; 14(1):329-351. https://doi.org/10.3390/molecules14010329
Chicago/Turabian StyleMooney, Sutton, Jan-Erik Leuendorf, Christopher Hendrickson, and Hanjo Hellmann. 2009. "Vitamin B6: A Long Known Compound of Surprising Complexity" Molecules 14, no. 1: 329-351. https://doi.org/10.3390/molecules14010329
APA StyleMooney, S., Leuendorf, J.-E., Hendrickson, C., & Hellmann, H. (2009). Vitamin B6: A Long Known Compound of Surprising Complexity. Molecules, 14(1), 329-351. https://doi.org/10.3390/molecules14010329