Substituted Pyrazinecarboxamides as Abiotic Elicitors of Flavolignan Production in Silybum marianum (L.) Gaertn Cultures in Vitro

Abstract
:1. Introduction
2. Results and Discussion
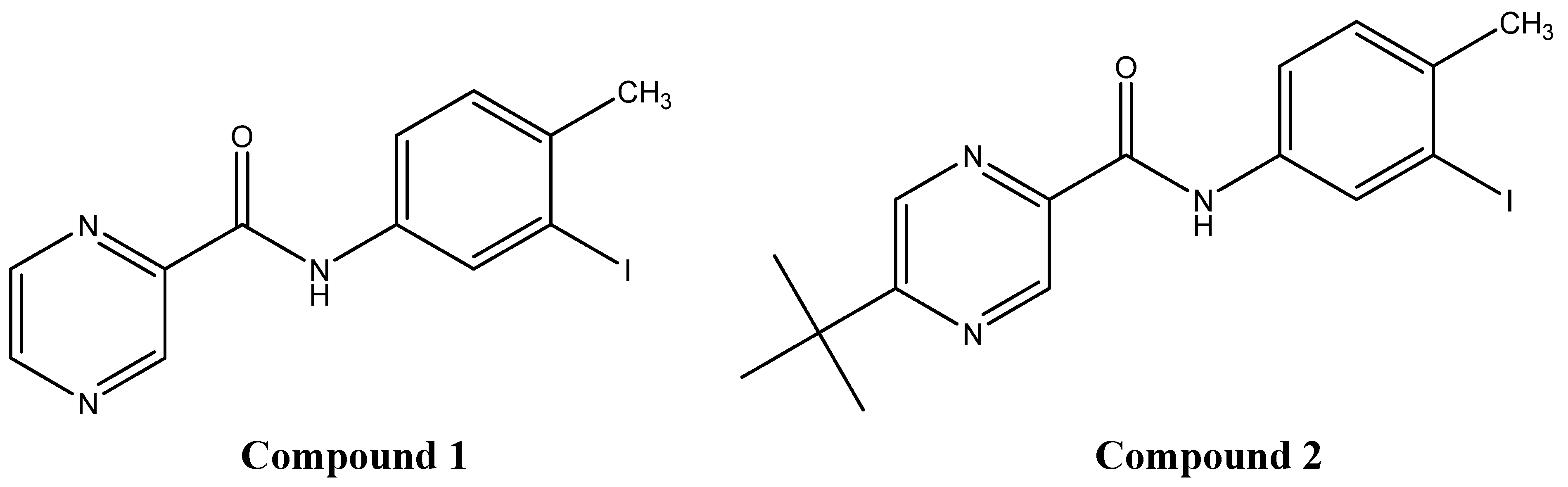
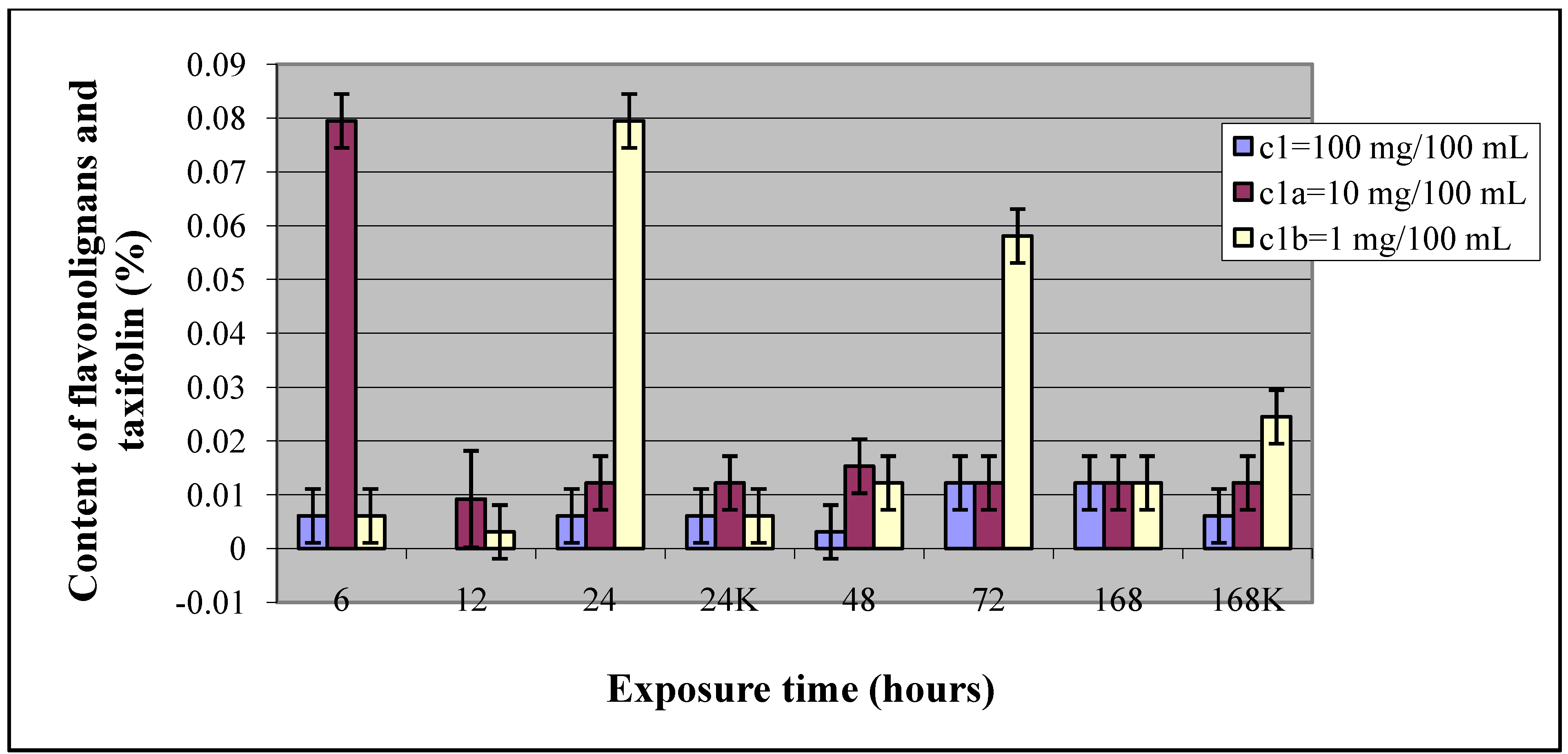
2.1. Elicitation with N-(3-iodo-4-methylphenyl)pyrazine-2-carboxamide (1)

{kind=link}
{kind=link}
{kind=link}
| Elicitor concentration (mg/100 mL) | Exposure time (hours) | Content of flavonolignans and taxifolin (%) | SD | Statistic value - t |
|---|---|---|---|---|
| 6 | 0.006 | 0.005 | 0.00 | |
| 12 | 0.000 | 0.000 | 1.63 | |
| 24 | 0.006 | 0.005 | 0.00 | |
| 100 (c1) | 24K | 0.006 | 0.005 | - |
| 48 | 0.003 | 0.005 | 0.577 | |
| 72 | 0.012 | 0.005 | 1.16 | |
| 168 | 0.012 | 0.005 | 1.16 | |
| 168K | 0.006 | 0.005 | - | |
| 6 | 0.080 | 0.005 | 12.7 | |
| 12 | 0.009 | 0.009 | 0.408 | |
| 24 | 0,012 | 0.005 | 0,00 | |
| 10 (c1a) | 24K | 0.012 | 0.005 | - |
| 48 | 0.015 | 0.005 | 0.577 | |
| 72 | 0.012 | 0.005 | 0.00 | |
| 168 | 0.012 | 0.005 | 0.00 | |
| 168K | 0.012 | 0.005 | - | |
| 6 | 0.006 | 0.005 | 0.00 | |
| 12 | 0.003 | 0.005 | 0.577 | |
| 24 | 0.080 | 0.005 | 13.9 | |
| 1 (c1b) | 24K | 0.006 | 0.005 | - |
| 48 | 0.012 | 0.005 | 1.16 | |
| 72 | 0.058 | 0.005 | 9.82 | |
| 168 | 0.012 | 0.005 | 231 | |
| 168K | 0.025 | 0.005 | - |

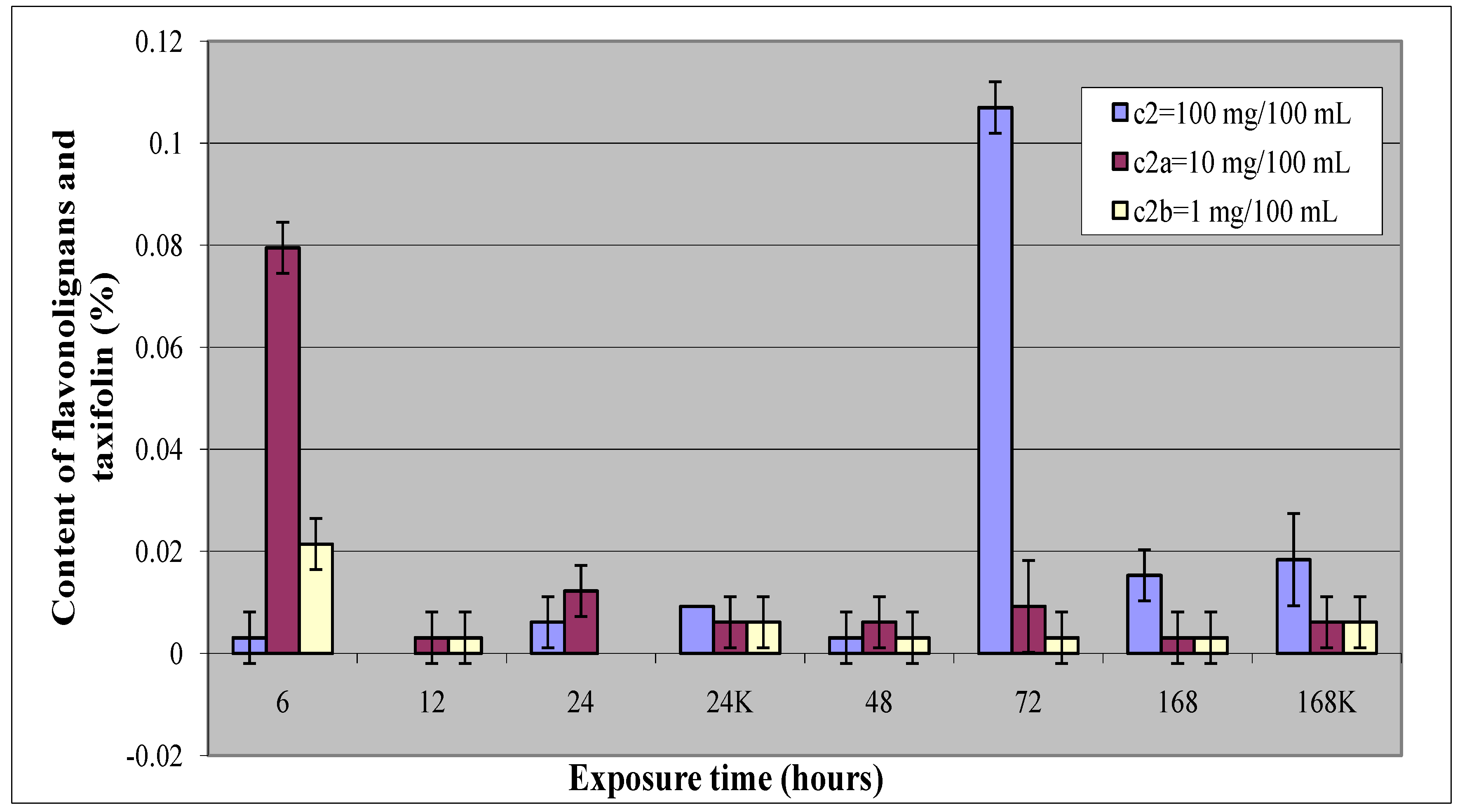
2.2. Elicitation with N-(3-iodo-4-methylphenyl)-5-tert-butylpyrazine-2-carboxamide (2)
| Elicitor concentration (mg/100 mL) | Exposure time (hours) | Content of flavonolignans and taxifolin (%) | SD | Statistical value - t |
|---|---|---|---|---|
| 6 | 0.003 | 0.005 | 1.63 | |
| 12 | 0.000 | 0.000 | - | |
| 24 | 0.006 | 0.005 | 0.816 | |
| 100 (c2) | 24K | 0.009 | 0.000 | - |
| 48 | 0.003 | 0.005 | 1.63 | |
| 72 | 0.107 | 0.005 | 26.1 | |
| 168 | 0.015 | 0.005 | 0.408 | |
| 168K | 0.018 | 0.009 | - | |
| 6 | 0.080 | 0.005 | 13.9 | |
| 12 | 0.003 | 0.005 | 0.577 | |
| 24 | 0.012 | 0.005 | 1.16 | |
| 10 (c2a) | 24K | 0.006 | 0.005 | - |
| 48 | 0.006 | 0.005 | 0.00 | |
| 72 | 0.009 | 0.009 | 0.408 | |
| 168 | 0.003 | 0.005 | 0.577 | |
| 168K | 0.006 | 0.005 | - | |
| 6 | 0.021 | 0.005 | 2.89 | |
| 12 | 0.003 | 0.005 | 0.577 | |
| 24 | 0,000 | 0.000 | 1.63 | |
| 1 (c2b) | 24K | 0.006 | 0.005 | - |
| 48 | 0.003 | 0.005 | 0.577 | |
| 72 | 0.003 | 0.005 | 0.577 | |
| 168 | 0.003 | 0.005 | 0.577 | |
| 168K | 0.006 | 0.005 | - |

3. Experimental
3.1. Abiotic elicitors in Silybum marianum L. callus culture
3.2. Material and methods

3.3. Flavonolignan analysis
4. Conclusions
Acknowledgements
References
- Doležal, M. Biologicky aktivní pyraziny přírodního a syntetického původu. Chem. Listy 2006, 100, 959–966. [Google Scholar]
- Doležal, M.; Miletín, M.; Kuneš, J.; Kráľová, K. Substituted amides of pyrazine-2-carboxylic acids, their synthesis and biological activity. Molecules 2002, 7, 363–373. [Google Scholar] [CrossRef]
- Ricci, D.; Maggiali, C.A.; Ronchini, F.; Tirillini, B.; Fraternale, D. Auxin activity of diazine carboxylic acids. Phytochemistry 1991, 30, 2821–2824. [Google Scholar] [CrossRef]
- Doležal, M.; Čmedlová, P.; Palek, L.; Vinšová, J.; Kuneš, J.; Buchta, V.; Jampílek, J.; Kráľová, K. Synthesis and biological evaluation of pyrazinecarboxamides. Eur. J. Med. Chem. 2008, 43, 1105–1113. [Google Scholar] [CrossRef]
- Tůmová, L.; Ostrožlík, P. Ononis arvensis in vitro - abiotická elicitace. Czech. Slov. Pharm. 2002, 4, 173–176. [Google Scholar]
- Doležal, M.; Tůmová, L.; Kešetovičová, D.; Tůma, J.; Kráľová, K. Substituted N-phenylpyrazine-2-carboxamides, their synthesis and evaluation as herbicides and abiotic elicitors. Molecules 2007, 12, 2589–2598. [Google Scholar] [CrossRef]
- Arvind, S.; Negi, J.K.; Kumar, S.L.; Karuna, S.; Gupta, M.M.; Khanuja, S.P.S. Recent advances in plant hepatoprotectives: A chemical and biological profile of some important leads. Med. Res. Rev. 2008, 28, 746–772. [Google Scholar] [CrossRef]
- Gažák, R.; Walterová, D.; Kren, V. Silybin and silymarin – new and emerging application in medicine. Curr. Med. Chem. 2007, 14, 315–338. [Google Scholar] [CrossRef]
- Jung, H.J.; Park, J.W.; Lee, J.S.; Lee, S.R.; Jang, B.C.; Suh, S.I.; Suh, M.H.; Baek, W.K. Silibinin inhibits expression of HIF-1 alpha through suppression of protein translation in prostate cancer cells. Biochem. Biophys. Res. Com. 2009, 390, 71–76. [Google Scholar] [CrossRef]
- Kim, S.; Choi, M.G.; Lee, H.S.; Lee, S.K.; Kim, S.H.; Kim, W.W.; Hur, S.M.; Kim, J.H.; Choe, J.H.; Nam, S.J.; Yang, J.H.; Kim, S.; Lee, J.E.; Kim, J.S. Silibinin suppresses TNF-α-induced MMP-9 expression in gastric cancer cells through inhibition of the MAPK pathway. Molecules 2009, 14, 4300–4311. [Google Scholar] [CrossRef]
- Singh, R.P.; Agarwal, R. Cosmeceuticals and silibinin. Clinic. Dermatol. 2009, 27, 479–484. [Google Scholar]
- Tůmová, L.; Gallová, K.; Řimáková, J.; Doležal, M.; Tůma, J. The effect of substituted amides of pyrazine-2-carboxylic acids on flavonolignan production in Silybum marianum culture in vitro. Acta Physiol. Plant. 2005, 27, 357–362. [Google Scholar] [CrossRef]
- Geneva, M.; Stancheva, I.; Sichanova, M.; Boychinova, M.; Georgiev, G.; Doležal, M. Improvement of milk thistle (Silybum marianum L.) seed yield and quality with foliar fertilization and growth effector MD 148/II. Gen. Appl. Plant Physiol. 2008, 34, 309–317. [Google Scholar]
- Stancheva, I.; Georgiev, G.; Geneva, M.; Ivanova, A.; Doležal, M.; Tůmová, L. Influence of foliar fertilization and growth effector 5-tert-butyl-N-m-tolylpyrazine-2-carboxamide (MD 148/II) on the milk thistle (Silybum marianum L.) seed yield and quality. J. Plant Nutr. 2010, 33. in press. [Google Scholar]
- Doležal, M.; Zitko, J.; Kešetovičová, D.; Kuneš, J.; Svobodová, M. Substituted N-phenylpyrazine-2-carboxamides: Synthesis and antimycobacterial evaluation. Molecules 2009, 14, 4180–4189. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 437–497. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds 1 and 2 are available from the authors.
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tůmová, L.; Tůma, J.; Megušar, K.; Doležal, M. Substituted Pyrazinecarboxamides as Abiotic Elicitors of Flavolignan Production in Silybum marianum (L.) Gaertn Cultures in Vitro. Molecules 2010, 15, 331-340. https://doi.org/10.3390/molecules15010331
Tůmová L, Tůma J, Megušar K, Doležal M. Substituted Pyrazinecarboxamides as Abiotic Elicitors of Flavolignan Production in Silybum marianum (L.) Gaertn Cultures in Vitro. Molecules. 2010; 15(1):331-340. https://doi.org/10.3390/molecules15010331
Chicago/Turabian StyleTůmová, Lenka, Jiří Tůma, Klara Megušar, and Martin Doležal. 2010. "Substituted Pyrazinecarboxamides as Abiotic Elicitors of Flavolignan Production in Silybum marianum (L.) Gaertn Cultures in Vitro" Molecules 15, no. 1: 331-340. https://doi.org/10.3390/molecules15010331
APA StyleTůmová, L., Tůma, J., Megušar, K., & Doležal, M. (2010). Substituted Pyrazinecarboxamides as Abiotic Elicitors of Flavolignan Production in Silybum marianum (L.) Gaertn Cultures in Vitro. Molecules, 15(1), 331-340. https://doi.org/10.3390/molecules15010331