Quercetin as a Potential Modulator of P-Glycoprotein Expression and Function in Cells of Human Pancreatic Carcinoma Line Resistant to Daunorubicin

Abstract
:Introduction
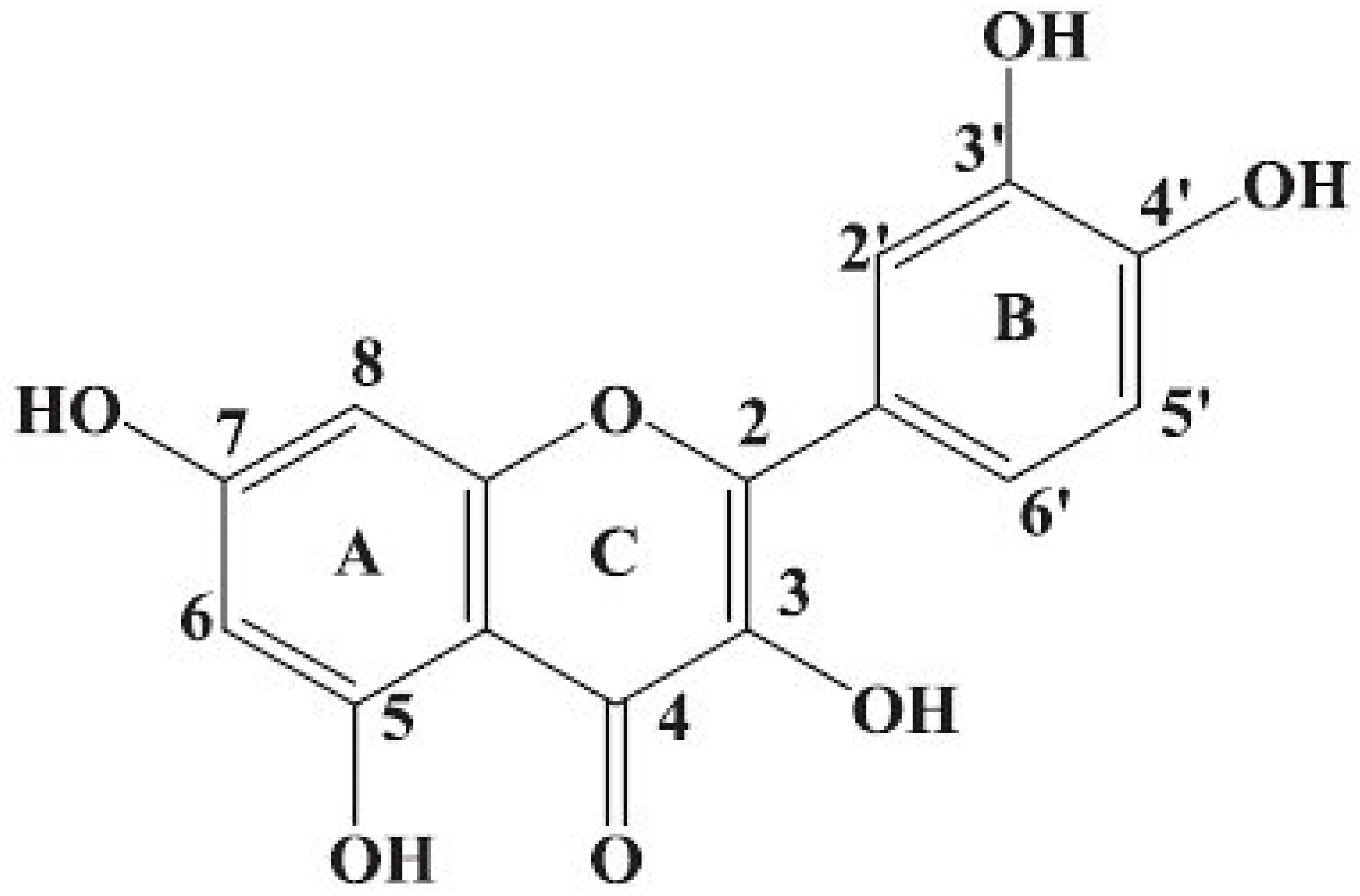
Results and Discussion

{kind=link}
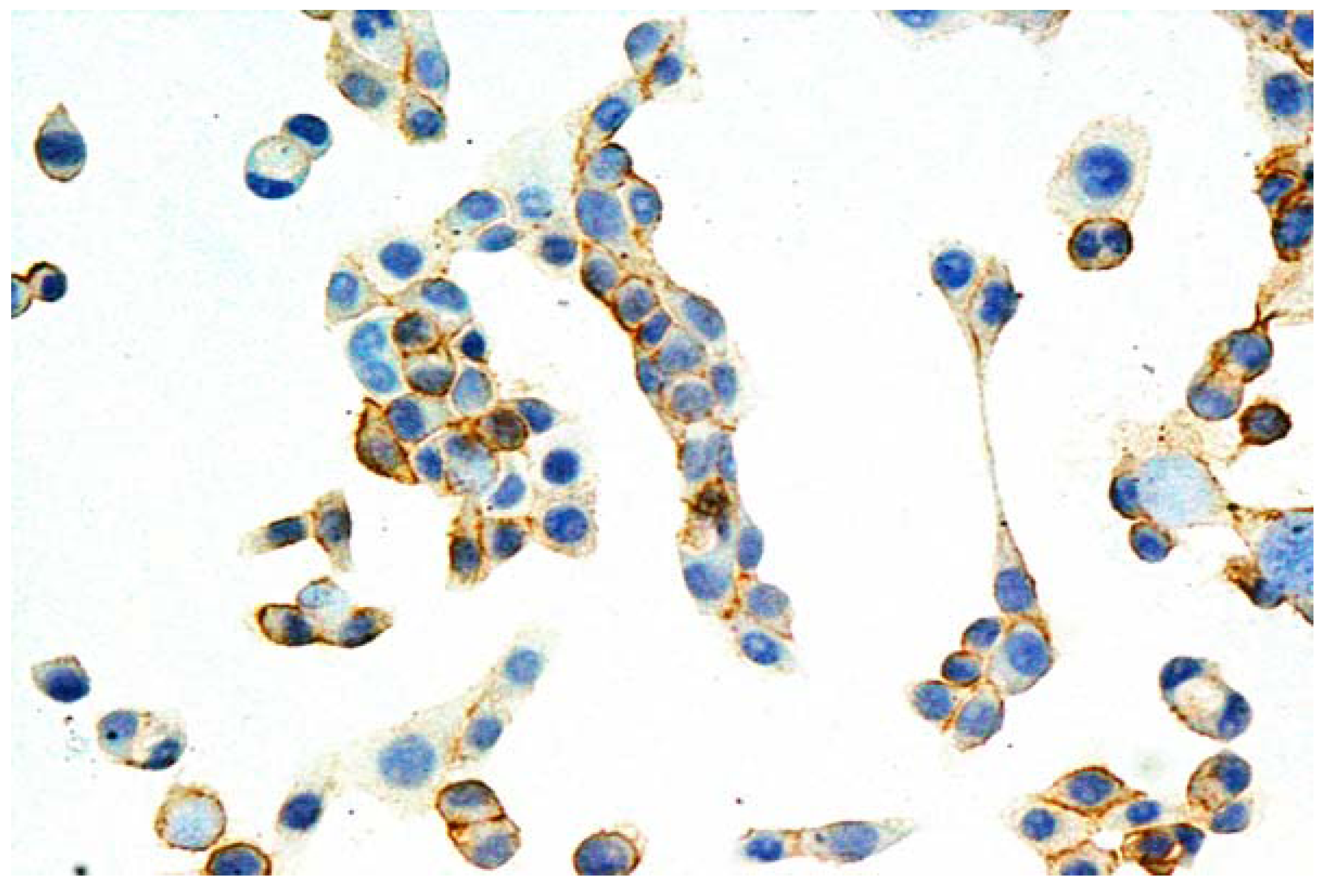{kind=link}
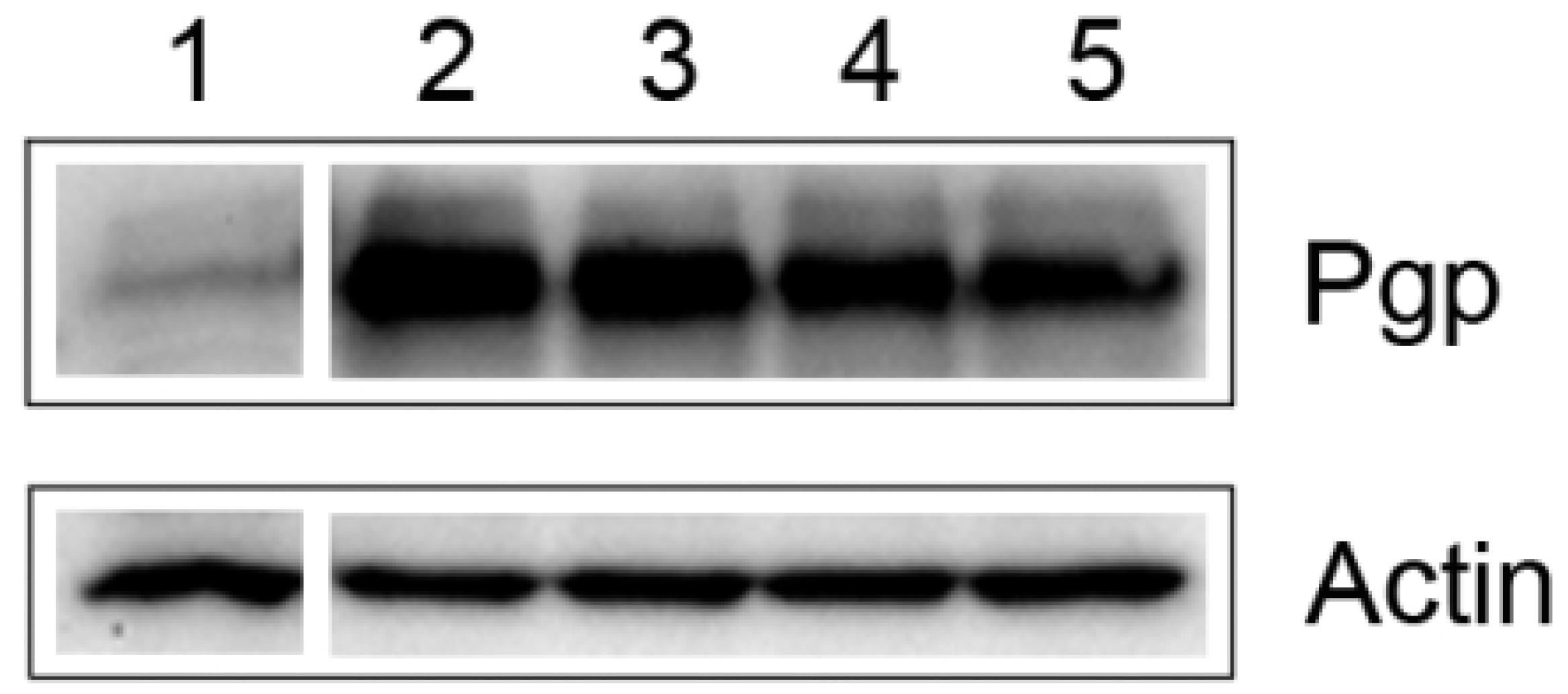{kind=link}
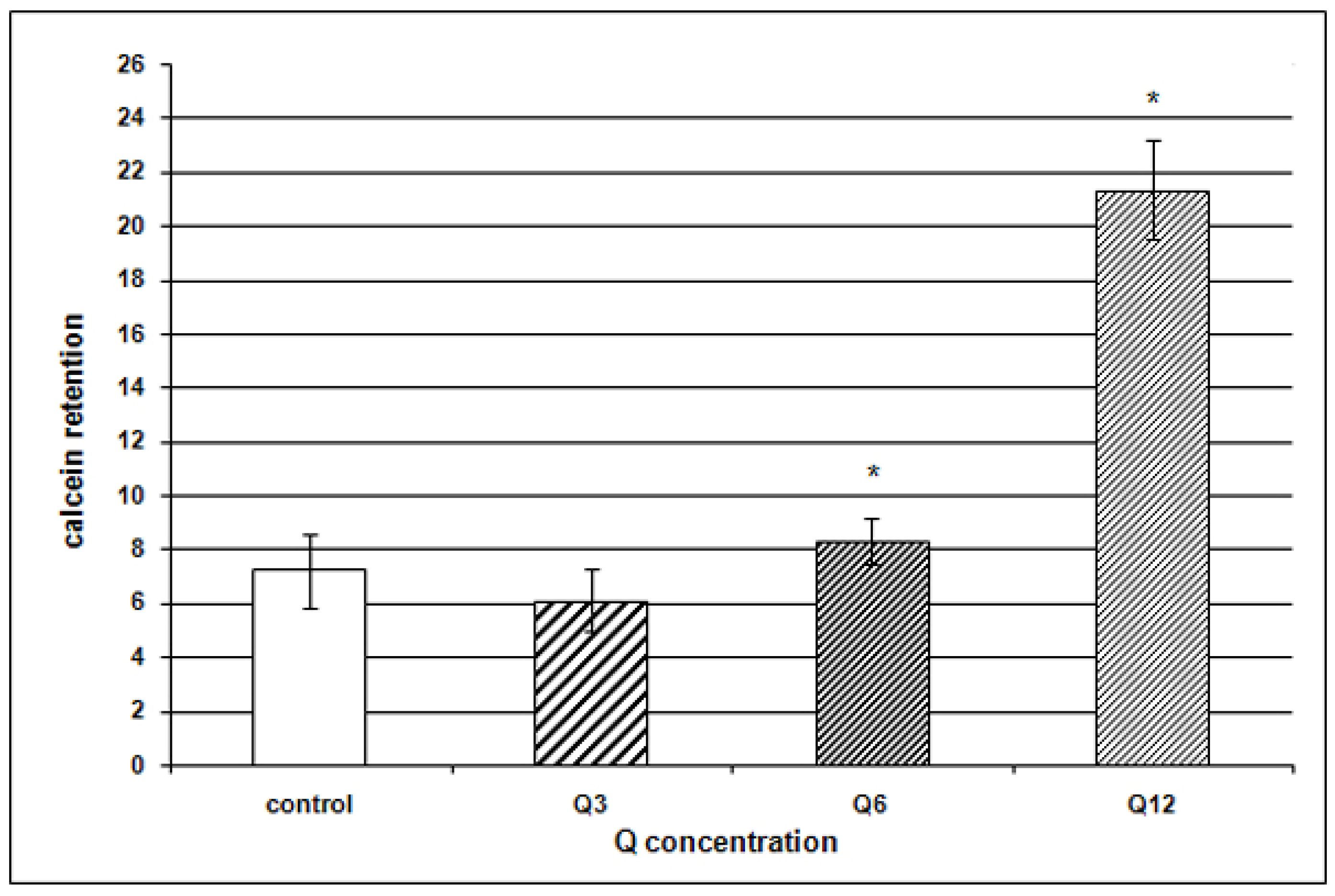{kind=link}
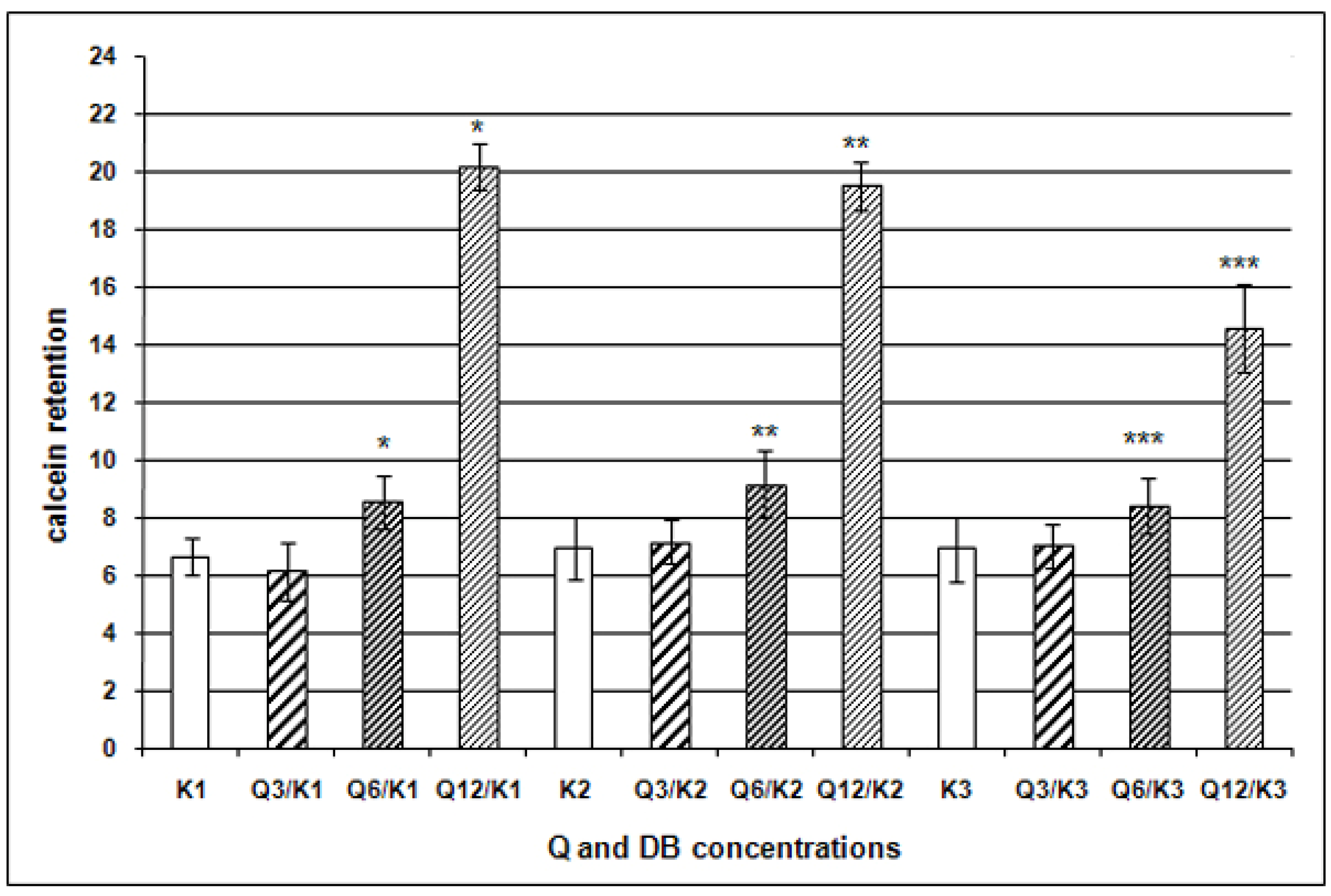{kind=link}
{kind=link}
{kind=link}
| Q concentration [µM] | C (0) | 1 | 2 | 3 | 6 | 12 | 25 | 50 |
|---|---|---|---|---|---|---|---|---|
| OD | 2.472 | 2.363 | 2.175 | 1.930 | 1.596 | 0.650 | 0.351 | 0.273 |
| SD | 0.024 | 0.033 | 0.033 | 0.033 | 0.048 | 0.030 | 0.030 | 0.026 |
| % of C value | 100 | 95.584 | 87.999 | 78.074 | 64.556 | 26.281 | 14.186 | 11.057 |
| Q [µM] | C (0) | 1 | 2 | 3 | 6 | 12 | 25 | 50 |
|---|---|---|---|---|---|---|---|---|
| OD | 2.363 | 2.317 | 2.277 | 2.254 | 1.809 | 1.192 | 1.022 | 0.649 |
| SD | 0.035 | 0.119 | 0.021 | 0.059 | 0.013 | 0.008 | 0.058 | 0.039 |
| % of C value | 100 | 98.053 | 96.360 | 95.380 | 76.552 | 50.444 | 43.235 | 27.483 |




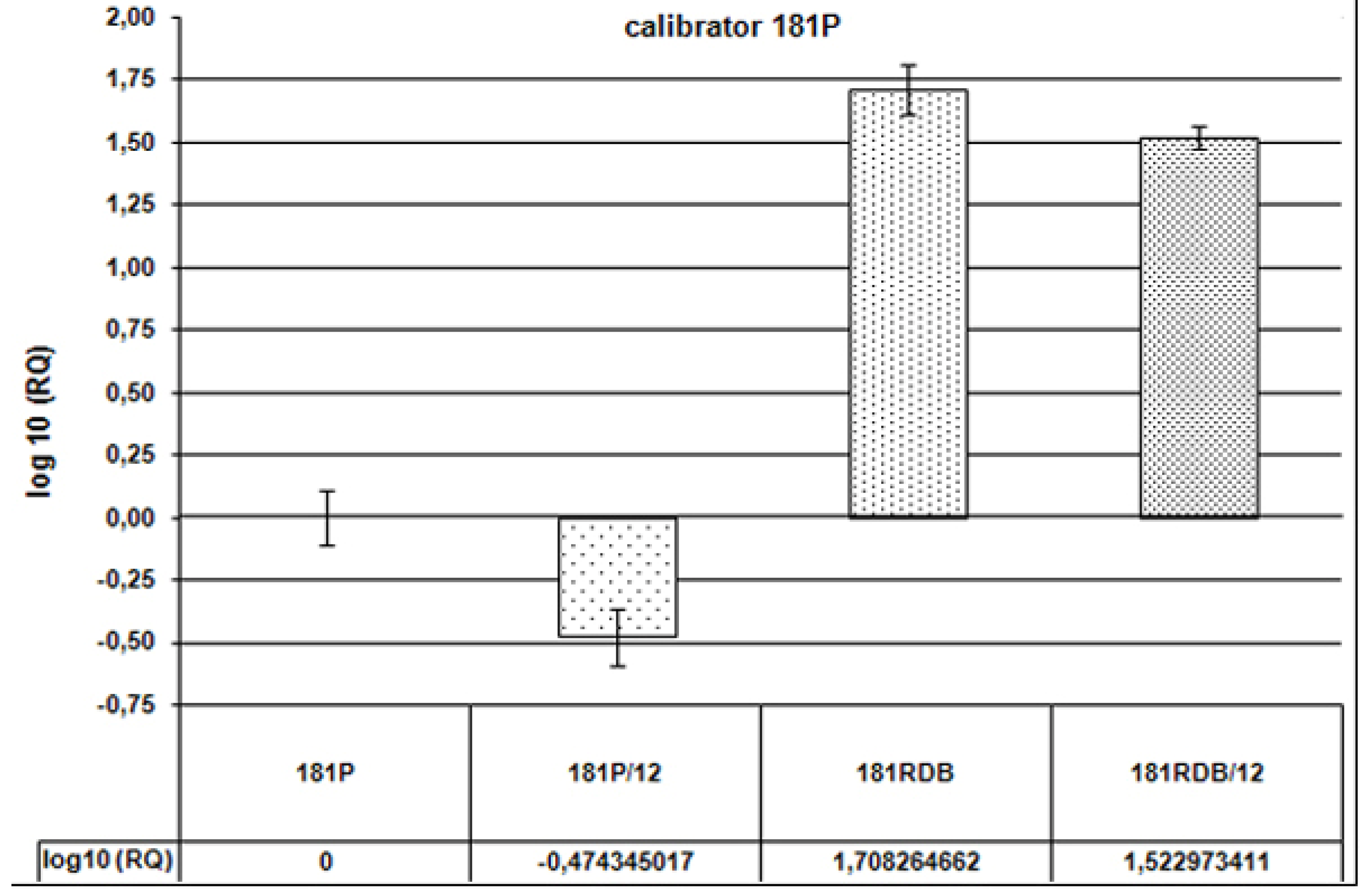
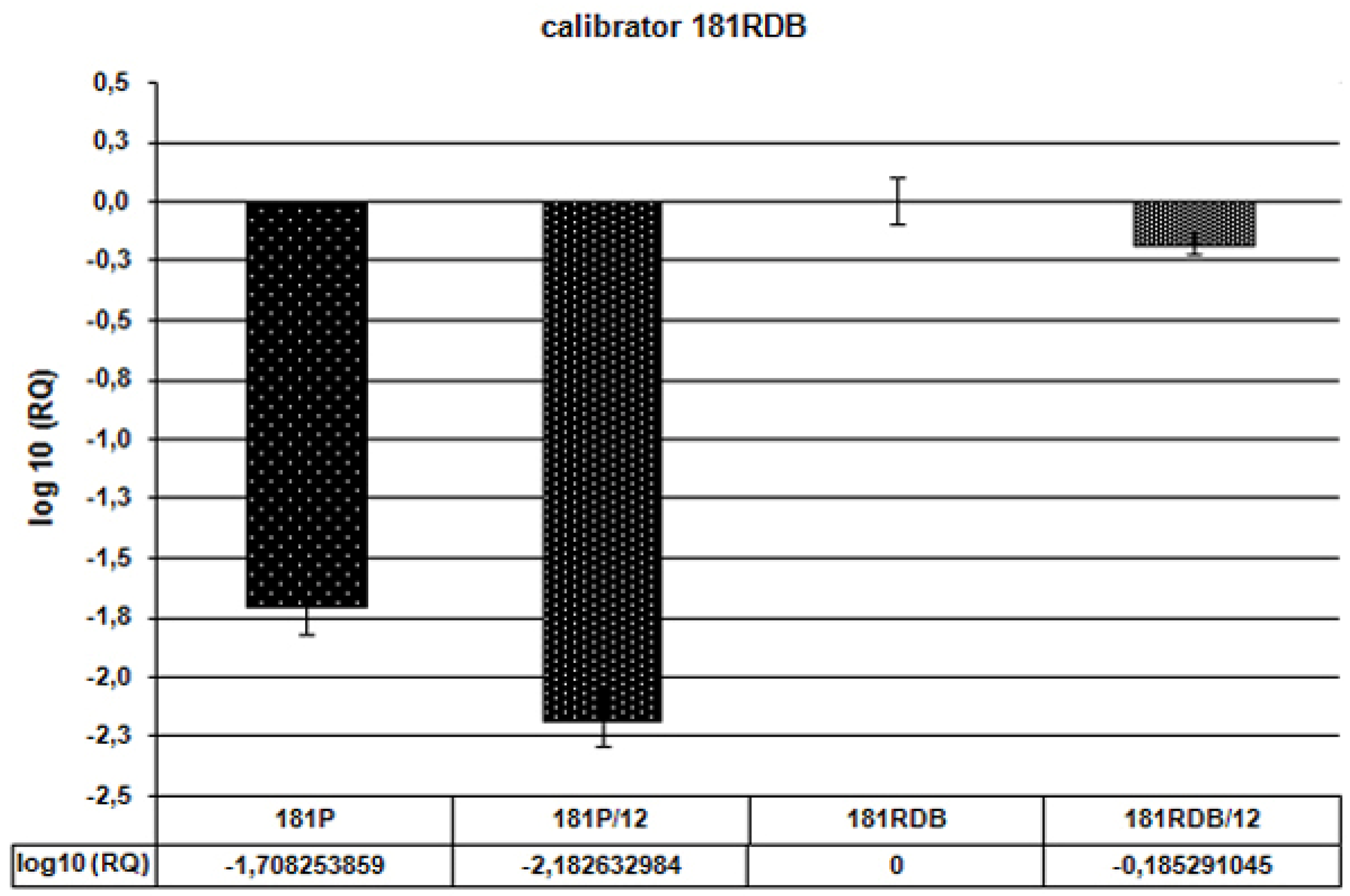
Conclusions
Experimental
Cell lines
Calculation of resistance index for Q on the basis of cytotoxic tests
Immunocytochemical analysis of P-gp expression
Western blotting analysis of P-gp
Transport function of P-gp
Examination of ABCB1 gene expression
Statistical analysis
Acknowledgements
- Sample Availability: Samples of the compounds are available from the authors.
References
- Gottesman, M.M.; Ling, V. The molecular basis of multidrug resistance in cancer: The early years of P-glycoprotein research. FEBS Lett. 2006, 580, 998–1009. [Google Scholar] [CrossRef]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Shapiro, A.B.; Ling, V. Positively cooperative sites for drug transport by P-glycoprotein with distinct drug specificities. Eur. J. Biochem. 1997, 250, 130–137. [Google Scholar]
- Sharom, F.J.; Liu, R.; Romsicki, Y.; Lu, P. Insights into the structure and substrate interactions of the P-glycoprotein multidrug transporter from spectroscopic studies. Biochim. Biophys. Acta 1999, 1461, 327–345. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP-dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef]
- Laurent, G.; Jaffrezou, J.P. Signaling pathways activated by daunorubicin. Blood 2001, 98, 913–924. [Google Scholar] [CrossRef]
- Szulawska, A.; Czyz, M. Molecular mechanisms of anthracyclines action. Postepy Hig. Med. Dosw. 2006, 60, 78–100. [Google Scholar]
- Kim, Y.; Ma, A.G.; Kitta, K.; Fitch, S.N.; Ikeda, T.; Ihara, Y.; Simon, A.R.; Evans, T.; Suzuki, Y.J. Anthracycline-induced suppression of GATA-4 transcription factor: Implication in the regulation of cardiac myocyte apoptosis. Mol. Pharmacol. 2003, 63, 368–377. [Google Scholar] [CrossRef]
- Varma, M.V.; Ashokraj, Y.; Dey, C.S.; Panchagnula, R. P-glycoprotein inhibitors and their screening: A perspective from bioavailability enhancement. Pharmacol. Res. 2003, 48, 347–359. [Google Scholar] [CrossRef]
- Teodori, E.; Dei, S.; Scapecchi, S.; Gualtieri, F. The medicinal chemistry of multidrug resistance (MDR) reversing drugs. Farmaco 2002, 57, 385–415. [Google Scholar] [CrossRef]
- Formica, J.V.; Regelson, W. Review of the biology of Quercetin and related bioflavonoids. Food. Chem. Toxicol. 1995, 33, 1061–1080. [Google Scholar] [CrossRef]
- Erlund, I. Review of flavonoids quercetin, hesperetin, and naringenin. Dietary sources, bioactivities, bioavailability, and epidemiology. Nutr. Res. 2004, 24, 551–874. [Google Scholar]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar]
- Simic, A.; Manojlovic, D.; Segan, D.; Todorovic, M. Electrochemical behavior and antioxidant and prooxidant activity of natural phenolics. Molecules 2007, 12, 2327–2340. [Google Scholar] [CrossRef]
- Bast, A.; Haenen, G.R.; Bruynzeel, A.M.; Van der Vijgh, W.J. Protection by flavonoids against anthracycline cardiotoxicity: From chemistry to clinical trials. Cardiovasc. Toxicol. 2007, 7, 154–159. [Google Scholar] [CrossRef]
- Le Good, J.A.; Ziegler, W.H.; Parekh, D.B.; Alessi, D.R.; Cohen, P.; Parker, P.J. Protein kinase C isotypes controlled by phosphoinositide 3-kinase through the protein kinase PDK1. Science 1998, 281, 2042–2045. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Tran, E.; Nguyen, T.H.; Do, P.T.; Huynh, T.H.; Huynh, H. The role of activated MEK-ERK pathway in quercetin-induced growth inhibition and apoptosis in A549 lung cancer cells. Carcinogenesis 2004, 25, 647–659. [Google Scholar]
- Chen, D.; Dou, Q.P. Tea polyphenols and their roles in cancer prevention and chemotherapy. Int. J. Mol. Sci. 2008, 9, 1196–1206. [Google Scholar] [CrossRef]
- Yang, H.; Landis-Piwowar, K.R.; Chen, D.; Milacic, V.; Dou, Q.P. Natural compounds with proteasome inhibitory activity for cancer prevention and treatment. Curr. Protein Pept. Sci. 2008, 9, 227–239. [Google Scholar] [CrossRef]
- Di Pietro, A.; Conseil, G.; Perez-Victoria, J.M.; Dayan, G.; Baubichon-Cortay, H.; Trompier, D.; Steinfels, E.; Jault, J.M.; de Wet, H.; Maitrejean, M.; Comte, G.; Boumendjel, A.; Mariotte, A.M.; Dumontet, C.; McIntosh, D.B.; Goffeau, A.; Castanys, S.; Gamarro, F.; Barron, D. Modulation by flavonoids of cell multidrug resistance mediated by P-glycoprotein and related ABC transporters. Cell. Mol. Life Sci. 2002, 59, 307–322. [Google Scholar] [CrossRef]
- Kioka, N.; Hosokawa, N.; Komano, T.; Hirayoshi, K.; Nagata, K.; Ueda, K. Quercetin, a bioflavonoid, inhibits the increase of human multidrug resistance gene (MDR1) expression caused by arsenite. FEBS Lett. 1992, 301, 307–309. [Google Scholar] [CrossRef]
- Ozben, T. Mechanisms and strategies to overcome multiple drug resistance in cancer. FEBS Lett. 2006, 580, 2903–2909. [Google Scholar] [CrossRef]
- Bansal, T.; Awasthi, A.; Jaggi, M.; Khar, R.K.; Talegaonkar, S. Pre-clinical evidence for altered absorption and biliary excretion of irinotecan (CPT-11) in combination with quercetin: Possible contribution of P-glycoprotein. Life Sci. 2008, 83, 250–259. [Google Scholar] [CrossRef]
- Choi, Y.J.; Jeong, Y.J.; Lee, Y.J.; Kwon, H.M.; Kang, Y.H. (-)Epigallocatechin gallate and quercetin enhance survival signaling in response to oxidant-induced human endothelial apoptosis. J. Nutr. 2005, 135, 707–713. [Google Scholar]
- van Erk, M.J.; Roepman, P.; van der Lende, T.R.; Stierum, R.H.; Aarts, J.M.; van Bladeren, P.J.; van Ommen, B. Integrated assessment by multiple gene expression analysis of quercetin bioactivity on anticancer-related mechanisms in colon cancer cells in vitro. Eur. J. Nutr. 2005, 44, 143–156. [Google Scholar] [CrossRef]
- Borska, S.; Gebarowska, E.; Wysocka, T.; Drag-Zalesinska, M.; Zabel, M. The effects of quercetin vs cisplatin on proliferation and the apoptotic process in A549 and SW1271 cell lines in in vitro conditions. Folia Morphol. (Warsz) 2004, 63, 103–105. [Google Scholar]
- Sarkadi, B.; Muller, M. Search for specific inhibitors of multidrug resistance in cancer. Semin. Cancer Biol. 1997, 8, 171–182. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Zeng, S. Establishment of a P-glycoprotein substrate screening model and its preliminary application. World J. Gastroenterol. 2004, 10, 1365–1368. [Google Scholar]
- Chieli, E.; Romiti, N.; Rodeiro-Guerra, I.; Garrido-Garrido, G. In vitro effects of Mangifera indica and polyphenols derived on ABCB1/P-glycoprotein activity. Food Chem. Toxicol. 2009. [Google Scholar] [CrossRef]
- Hayeshi, R.; Masimirembwa, C.; Mukanganyama, S.; Ungell, A.L. The potential inhibitory effect of antiparasitic drugs and natural products on P-glycoprotein mediated efflux. Eur. J. Pharm. Sci. 2006, 29, 70–81. [Google Scholar] [CrossRef]
- Khantramat, O.; Chaiwangyen, W.; Limtrakul, P. Screening of flavonoids for their potential inhibitory effect on P-glycoprotein activity in human cervical carcinoma KB cells. Chiang Mai Med. Bull. 2004, 43, 45–56. [Google Scholar]
- O'Leary, K.A.; de Pascual-Tereasa, S.; Needs, P.W.; Bao, Y.P.; O'Brien, N.M.; Williamson, G. Effect of flavonoids and vitamin E on cyclooxygenase-2 (COX-2) transcription. Mutat. Res. 2004, 551, 245–254. [Google Scholar] [CrossRef]
- Badhan, R.; Penny, J. In silico modelling of the interaction of flavonoids with human P-glycoprotein nucleotide-binding domain. Eur. J. Med.Chem. 2006, 41, 285–295. [Google Scholar] [CrossRef]
- Mitsunaga, Y.; Takanaga, H.; Matsuo, H.; Naito, M.; Tsuruo, T.; Ohtani, H.; Sawada, Y. Effect of bioflavonoids on vincristine transport across blood-brain barrier. Eur. J. Pharmacol. 2000, 395, 193–201. [Google Scholar] [CrossRef]
- Kothan, S.; Dechsupa, S.; Leger, G.; Moretti, J.L.; Vergote, J.; Mankhetkorn, S. Spontaneous mitochondrial membrane potential change during apoptotic induction by quercetin in K562 and K562/adr cells. Can. J. Physiol. Pharmacol. 2004, 82, 1084–1090. [Google Scholar] [CrossRef]
- Nabekura, T.; Kamiyama, S.; Kitagawa, S. Effects of dietary chemopreventive phytochemicals on P-glycoprotein function. Biochem. Biophys. Res. Commun. 2005, 327, 866–870. [Google Scholar] [CrossRef]
- Critchfield, J.W.; Welsh, C.J.; Phang, J.M.; Yeh, G.C. Modulation of adriamycin accumulation and efflux by flavonoids in HCT-15 colon cells. Activation of P-glycoprotein as a putative mechanism. Biochem. Pharmacol. 1994, 48, 1437–1445. [Google Scholar]
- Morris, M.E.; Zhang, S. Flavonoid-drug interactions: Effects of flavonoids on ABC transporters. Life Sci. 2006, 78, 2116–2130. [Google Scholar] [CrossRef]
- O'Prey, J.; Brown, J.; Fleming, J.; Harrison, P.R. Effects of dietary flavonoids on major signal transduction pathways in human epithelial cells. Biochem. Pharmacol. 2003, 66, 2075–2088. [Google Scholar]
- Rosner, K.; Ropke, C.; Pless, V.; Skovgaard, G.L. Late type apoptosis and apoptosis free lethal effect of quercetin in human melanoma. Biosci. Biotechnol. Biochem. 2006, 70, 2169–2177. [Google Scholar] [CrossRef]
- Shapiro, A.B.; Ling, V. Effect of quercetin on Hoechst 33342 transport by purified and reconstituted P-glycoprotein. Biochem. Pharmacol. 1997, 53, 587–596. [Google Scholar]
- Shapiro, A.B.; Ling, V. Using purified P-glycoprotein to understand multidrug resistance. J. Bioenerg. Biomembr. 1995, 27, 7–13. [Google Scholar] [CrossRef]
- Son, Y.O.; Lee, K.Y.; Kook, S.H.; Lee, J.C.; Kim, J.G.; Jeon, Y.M.; Jang, Y.S. Selective effects of quercetin on the cell growth and antioxidant defense system in normal versus transformed mouse hepatic cell lines. Eur. J. Pharmacol. 2004, 502, 195–204. [Google Scholar] [CrossRef]
- Nagy, H.; Goda, K.; Fenyvesi, F.; Bacso, Z.; Szilasi, M.; Kappelmayer, J.; Lustyik, G.; Cianfriglia, M.; Szabo, G., Jr. Distinct groups of multidrug resistance modulating agents are distinguished by competition of P-glycoprotein-specific antibodies. Biochem. Biophys. Res. Commun. 2004, 315, 942–949. [Google Scholar] [CrossRef]
- Germann, U.A. P-glycoprotein--a mediator of multidrug resistance in tumour cells. Eur. J. Cancer 1996, 32A, 927–944. [Google Scholar] [CrossRef]
- Miura, T.; Muraoka, S.; Fujimoto, Y. Inactivation of creatine kinase induced by quercetin with horseradish peroxidase and hydrogen peroxide. pro-oxidative and anti-oxidative actions of quercetin. Food Chem. Toxicol. 2003, 41, 759–765. [Google Scholar] [CrossRef]
- Moon, Y.J.; Wang, X.; Morris, M.E. Dietary flavonoids: Effects on xenobiotic and carcinogen metabolism. Toxicol. In Vitro 2006, 20, 187–210. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Dunnick, J.K.; Hailey, J.R. Toxicity and carcinogenicity studies of quercetin, a natural component of foods. Fundam. Appl. Toxicol. 1992, 19, 423–431. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef]
- Okamoto, T. Safety of quercetin for clinical application (Review). Int. J. Mol. Med. 2005, 16, 275–278. [Google Scholar]
- Gupta, C.; Vikram, A.; Tripathi, D.N.; Ramarao, P.; Jena, G.B. Antioxidant and antimutagenic effect of quercetin against DEN induced hepatotoxicity in rat. Phytother. Res. 2009. [Google Scholar] [CrossRef]
- Lage, H.; Jordan, A.; Scholz, R.; Dietel, M. Thermosensitivity of multidrug-resistant human gastric and pancreatic carcinoma cells. Int. J. Hyperthermia 2000, 16, 291–303. [Google Scholar] [CrossRef]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Tiberghien, F.; Loor, F. Ranking of P-glycoprotein substrates and inhibitors by a calcein-AM fluorometry screening assay. Anticancer Drugs 1996, 7, 568–578. [Google Scholar] [CrossRef]
© 2010 by the authors;
Share and Cite
Borska, S.; Sopel, M.; Chmielewska, M.; Zabel, M.; Dziegiel, P. Quercetin as a Potential Modulator of P-Glycoprotein Expression and Function in Cells of Human Pancreatic Carcinoma Line Resistant to Daunorubicin. Molecules 2010, 15, 857-870. https://doi.org/10.3390/molecules15020857
Borska S, Sopel M, Chmielewska M, Zabel M, Dziegiel P. Quercetin as a Potential Modulator of P-Glycoprotein Expression and Function in Cells of Human Pancreatic Carcinoma Line Resistant to Daunorubicin. Molecules. 2010; 15(2):857-870. https://doi.org/10.3390/molecules15020857
Chicago/Turabian StyleBorska, Sylwia, Miroslaw Sopel, Magdalena Chmielewska, Maciej Zabel, and Piotr Dziegiel. 2010. "Quercetin as a Potential Modulator of P-Glycoprotein Expression and Function in Cells of Human Pancreatic Carcinoma Line Resistant to Daunorubicin" Molecules 15, no. 2: 857-870. https://doi.org/10.3390/molecules15020857
APA StyleBorska, S., Sopel, M., Chmielewska, M., Zabel, M., & Dziegiel, P. (2010). Quercetin as a Potential Modulator of P-Glycoprotein Expression and Function in Cells of Human Pancreatic Carcinoma Line Resistant to Daunorubicin. Molecules, 15(2), 857-870. https://doi.org/10.3390/molecules15020857