Effects of Polysaccharide Elicitors from Endophytic Fusarium oxysporium Dzf17 on Growth and Diosgenin Production in Cell Suspension Culture of Dioscorea zingiberensis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Time Courses of Cell Growth and Diosgenin Production

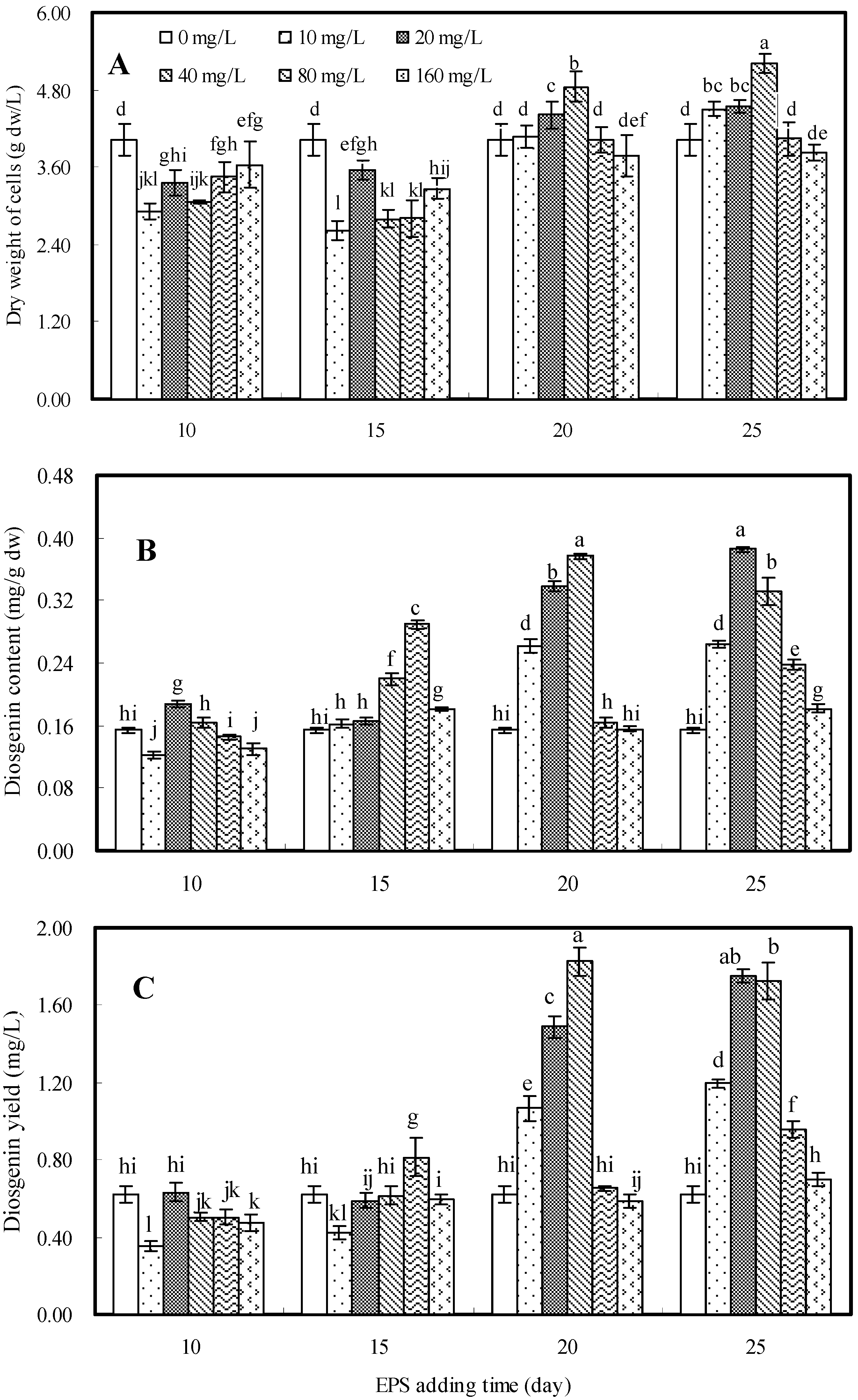
2.2. Effects of EPS on Cell Growth and Diosgenin Production

2.3. Effects of WPS on Cell Growth and Diosgenin Production

2.4. Effects of SPS on Cell Growth and Diosgenin Production

2.5. Dynamics of Cell Growth and Diosgenin Production after Addition of WPS

3. Experimental
3.1. Cell Culture
3.2. Cultivation of Endophytic Fungus Fusarium Oxysporum Dzf17
3.3. Preparation of Exopolysaccharide
3.4. Preparation of Water-Extracted Mycelial Polysaccharide and Sodium Hydroxide-Extracted Mycelial Polysaccharide
3.5. Elicitation Treatment of Suspension Cells
3.6. Cell Biomass Determination
3.7. Extraction and Determination of Diosgenin
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Li, H.; Ni, J. Treatment of wastewater from Dioscorea zingiberensis tubers used for producing steroid hormones in a microbial fuel cell. Bioresour. Technol. 2011, 102, 2731–2735. [Google Scholar] [CrossRef]
- Chen, Y.; Fan, J.; Luo, Z.; Fu, Y. Rapid clonal propagation of Dioscorea zingiberensis. Plant Cell Tissue Organ Cult. 2003, 73, 75–80. [Google Scholar] [CrossRef]
- Liu, W.; Huang, W.; Sun, W.L.; Zhu, Y.L.; Ni, J.R. Production of diosgenin from yellow ginger (Dioscorea zingiberensis C. H. Wright) saponins by commercial cellulase. World J. Microbiol. Biotechnol. 2010, 26, 1171–1180. [Google Scholar] [CrossRef]
- Qin, H.; Tian, W.; Lin, C. A highly efficient synthesis of 22-deoxy-OSW-1 by utilizing the intact skeleton of diosgenin. Tetrahedron Lett. 2006, 47, 3217–3219. [Google Scholar] [CrossRef]
- Zhou, L.; Wu, J. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep. 2006, 23, 789–810. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Farkya, S.; Srivastava, A.K.; Bisaria, V.S. Bioprocess considerations for production of secondary metabolites by plant cell suspension cultures. Biotechnol. Bioproc. Eng. 2002, 7, 138–149. [Google Scholar] [CrossRef]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plant Res. 2009, 3, 1222–1239. [Google Scholar]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Georgiev, M.I.; Weber, J.; Maciuk, A. Bioprocessing of plant cell cultures for mass production of targeted compounds. Appl. Microbiol. Biotechnol. 2009, 83, 809–823. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, C.; Li, J.; Wang, S.; Wu, J. Heptasaccharide and octasaccharide isolated from Paris polyphylla var. yunnanensis and their plant growth-regulatory activity. Plant Sci. 2003, 165, 571–575. [Google Scholar] [CrossRef]
- Zhou, L.; Cao, X.; Zhang, R.; Peng, Y.; Zhao, S.; Wu, J. Stimulation of saponin production in Panax ginseng hairy roots by two oligosaccharides from Paris polyphylla var. yunnanensis. Biotechnol. Lett. 2007, 29, 631–634. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, L.; Wu, J. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures. Appl. Microbiol. Biotechnol. 2010, 87, 137–144. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, L.; Wu, J. Promotion of Salvia miltiorrhiza hairy root growth and tanshinone production by polysaccharide-protein fractions of plant growth-promoting rhizobacterium Bacillus cereus. Process Biochem. 2010, 45, 1517–1522. [Google Scholar] [CrossRef]
- Baldi, A.; Srivastava, A.K.; Bisaria, V.S. Fungal elicitors for enhanced production of secondary metabolites in plant cell suspension cultures. Soil Biol. 2009, 18, 373–380. [Google Scholar] [CrossRef]
- Zhao, J.; Zhu, W.-H.; Hu, Q. Selection of fungal elicitors to increase indole alkaloid accumulation in Catharanthus roseus suspension cell culture. Enzym. Microb. Tech. 2001, 28, 666–672. [Google Scholar] [CrossRef]
- Chen, H.; Chen, F. Effects of yeast elicitor on the growth and secondary metabolism of a high-tanshinone-producing line of the Ti transformed Salvia miltiorrhiza cells in suspension culture. Process Biochem. 2000, 35, 837–840. [Google Scholar] [CrossRef]
- Zhang, C.; Wu, J. Ethylene inhibitors enhance elicitor-induced paclitaxel production in suspension cultures of Taxus spp. cells. Enzym. Microb. Tech. 2003, 32, 71–77. [Google Scholar] [CrossRef]
- Singh, G.; Gavrieli, J.; Oakey, J.S.; Curtis, W.R. Interaction of methyl jasmonate, wounding and fungal elicitation during sesquiterpene induction in Hyoscyamus muticus in root cultures. Plant Cell Rep. 1998, 17, 391–395. [Google Scholar] [CrossRef]
- Lu, M.B.; Wong, H.L.; Teng, W.L. Effects of elicitation on the production of saponin in cell culture of Panax ginseng. Plant Cell Rep. 2001, 20, 674–677. [Google Scholar]
- Chong, T.M.; Abdullah, M.A.; Lai, Q.M.; Nor'Aini, F.M.; Lajis, N.H. Effective elicitation factors in Morinda elliptica cell suspension culture. Process Biochem. 2005, 40, 3397–3405. [Google Scholar] [CrossRef]
- Rojas, R.; Alba, J.; Magana-Plaza, I.; Cruz, F.; Ramos-Valdivia, A.C. Stimulated production of diosgenin in Dioscorea galeottiana cell suspension cultures by abiotic and biotic factors. Biotechnol. Lett. 1999, 21, 907–911. [Google Scholar] [CrossRef]
- Flores-Sanchez, I.J.; Pec, J.; Fei, J.; Choi, Y.H.; Dusek, J.; Verpoorte, R. Elicitation studies in cell suspension cultures of Cannabis sativa L. J. Biotechnol. 2009, 143, 157–168. [Google Scholar]
- Fukui, H.; Yoshikawa, N.; Tabata, M. Induction of shikonin formation by agar in Lithospermum erythrorhizon cell suspension cultures. Phytochemistry 1983, 22, 2451–2453. [Google Scholar]
- Bi, F.; Iqbal, S.; Arman, M.; Ali, A.; Hassan, M. Carrageenan as an elicitor of induced secondary metabolites and its effects on various growth characters of chickpea and maize plants. J. Saudi Chem. Soc. 2011, 15, 269–273. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef]
- Gimenez, C.; Cabrera, R.; Reina, M.; Gonzalez-Coloma, A. Fungal endophytes and their role in plant protection. Curr. Org. Chem. 2007, 11, 707–720. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, J.; Shan, T.; Cai, X.; Peng, Y. Spirobisnaphthalenes from fungi and their biological activities. Mini-Rev. Med. Chem. 2010, 10, 977–989. [Google Scholar] [CrossRef]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini-Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Zhang, R.; Li, P.; Xu, L.; Chen, Y.; Sui, P.; Zhou, L.; Li, J. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860. [Google Scholar] [CrossRef]
- Zhang, R.; Li, P.; Xu, L.; Chen, Y.; Sui, P.; Zhou, L.; Li, J. Enhancenment of diosgenin productiong in Dioscorea zingiberensis cell culture by oligosaccharide elicitor from its endophytic fungus Fusarium oxysporum. Nat. Prod. Commun. 2009, 4, 1459–1462. [Google Scholar]
- Fang, F.; Dai, C.; Wang, Y. Role of nitric oxide and hydrogen peroxide in the essential oil increasing of suspension cells from Atractylodes lancea induced by endophytic fungal Cunninghamella sp. AL4 elicitor. Chin. J. Biotechnol. 2009, 25, 1490–1496. [Google Scholar]
- Wang, Y.; Dai, C.; Zhao, Y.; Peng, Y. Fungal endophyte-induced volatile oil accumulation in Atractylodes lancea plantlets is mediated by nitric oxide, salicylic acid and hydrogen peroxide. Process Biochem. 2011, 46, 730–735. [Google Scholar] [CrossRef]
- De, D.; De, B. Elicitation of diosgenin production in Dioscorea floribunda by ethylene-generating agent. Fitoterapia 2005, 76, 153–156. [Google Scholar] [CrossRef]
- Wang, Z.; Luo, D.; Ena, C. Optimization of polysaccharides extraction from Gynostemma pentaphyllum Makino using uniform design. Carbohydr. Polym. 2007, 69, 311–317. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, W.; Ni, J. A promising clean process for production of diosgenin from Dioscorea zingiberensis C. H. Wright. J. Clean. Prod. 2010, 18, 242–247. [Google Scholar] [CrossRef]
- John, M.; Rohrig, H.; Schmidt, J.; Walden, R.; Shell, J. Cell signaling by oligosaccharides. Trends Plant Sci. 1997, 2, 111–115. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the polysaccharide elicitors are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, P.; Mou, Y.; Shan, T.; Xu, J.; Li, Y.; Lu, S.; Zhou, L. Effects of Polysaccharide Elicitors from Endophytic Fusarium oxysporium Dzf17 on Growth and Diosgenin Production in Cell Suspension Culture of Dioscorea zingiberensis. Molecules 2011, 16, 9003-9016. https://doi.org/10.3390/molecules16119003
Li P, Mou Y, Shan T, Xu J, Li Y, Lu S, Zhou L. Effects of Polysaccharide Elicitors from Endophytic Fusarium oxysporium Dzf17 on Growth and Diosgenin Production in Cell Suspension Culture of Dioscorea zingiberensis. Molecules. 2011; 16(11):9003-9016. https://doi.org/10.3390/molecules16119003
Chicago/Turabian StyleLi, Peiqin, Yan Mou, Tijiang Shan, Jianmei Xu, Yan Li, Shiqiong Lu, and Ligang Zhou. 2011. "Effects of Polysaccharide Elicitors from Endophytic Fusarium oxysporium Dzf17 on Growth and Diosgenin Production in Cell Suspension Culture of Dioscorea zingiberensis" Molecules 16, no. 11: 9003-9016. https://doi.org/10.3390/molecules16119003