Keeping an Eye on Myocilin: A Complex Molecule Associated with Primary Open-Angle Glaucoma Susceptibility

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
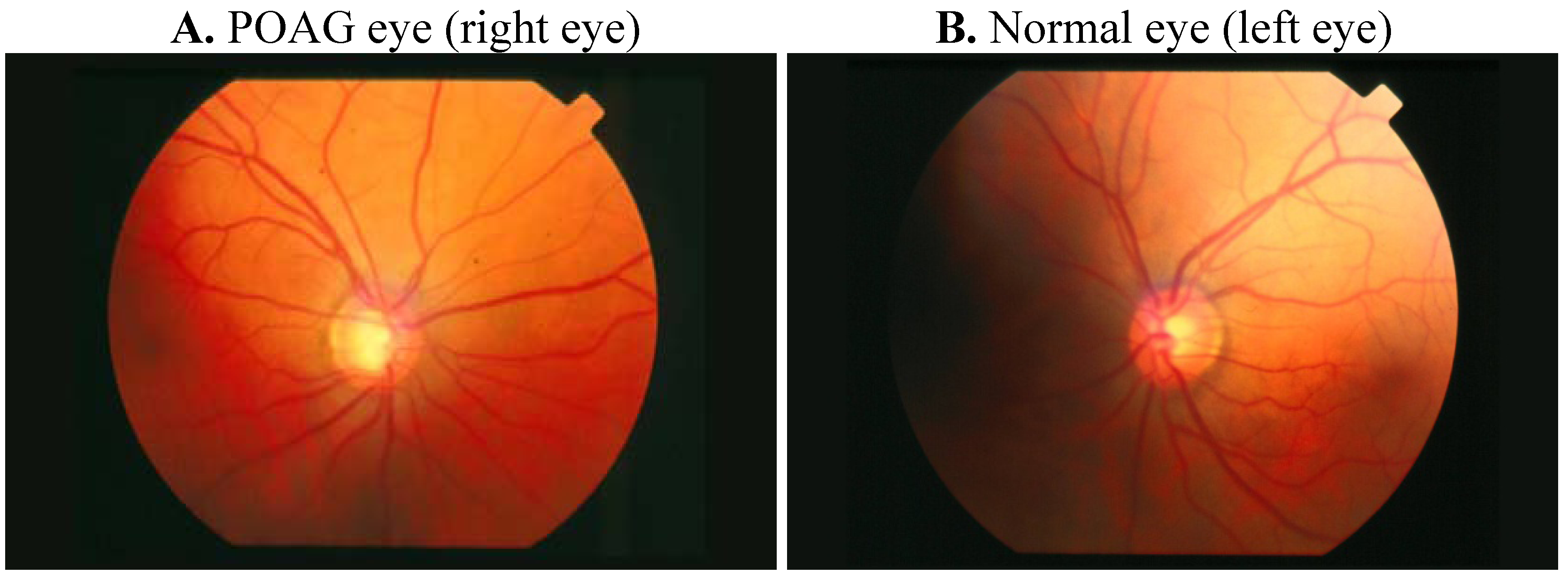
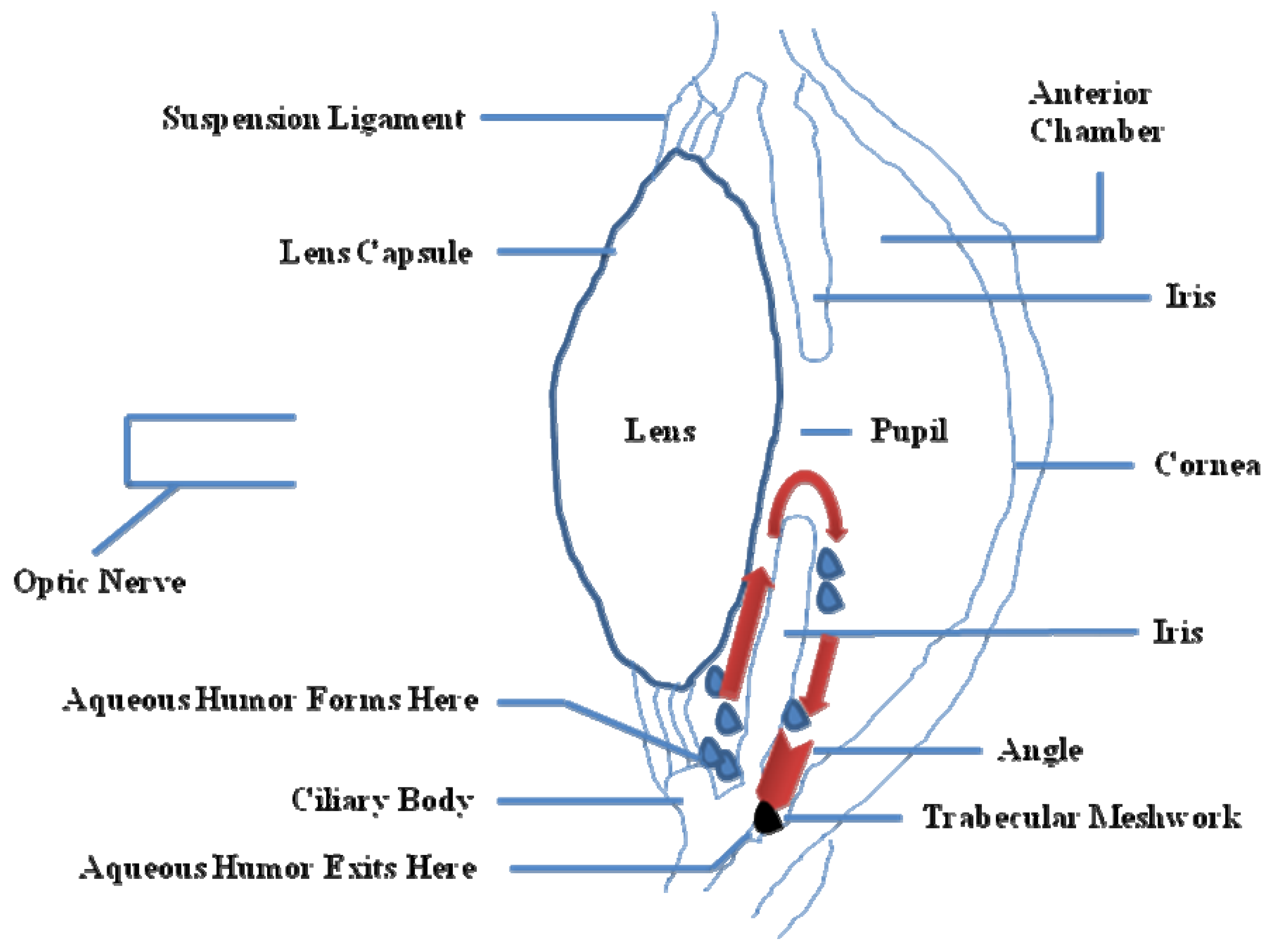
1.1. Physiopathology of POAG


1.2. Gene Mapping of MYOC
1.3. Expression and Regulation of MYOC
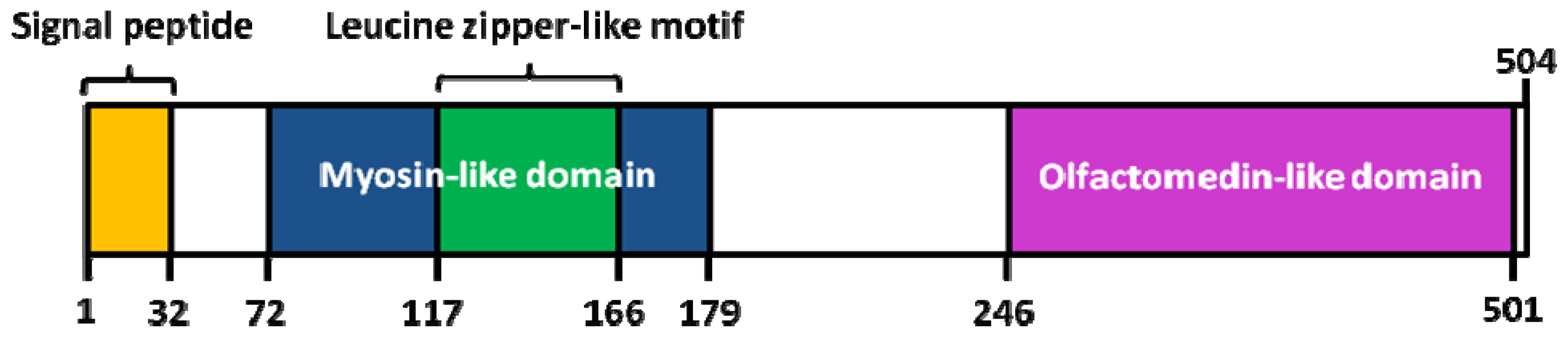
1.4. Structural Characterization of MYOC

1.5. Functions of MYOC
2. Results and Discussion
2.1. Myocilin-Molecules Interactions in POAG Etiology
2.2. MYOC Variants in POAG Etiology
2.3. Misfolding and Aggregation of Myocilin in POAG Etiology
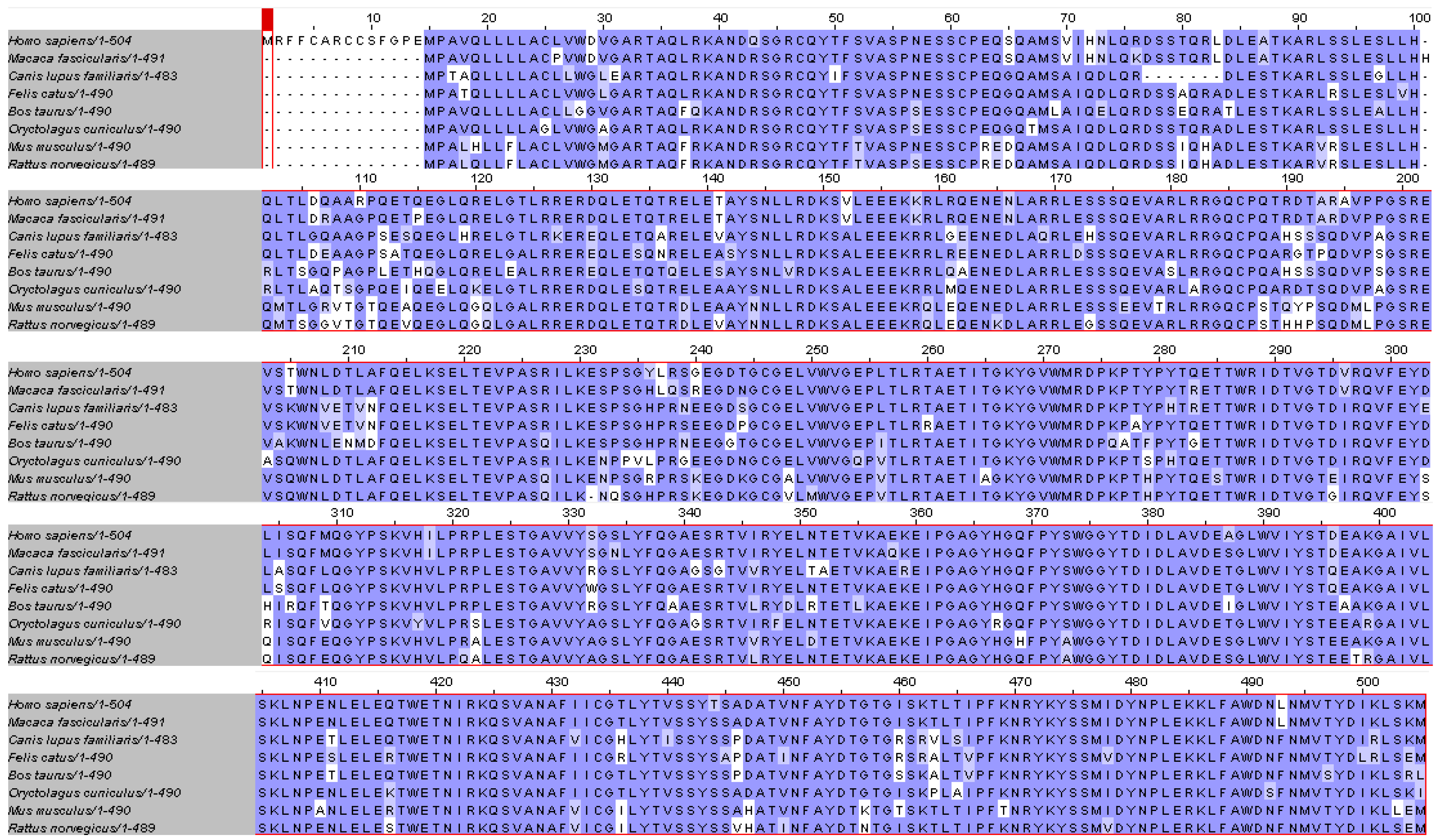
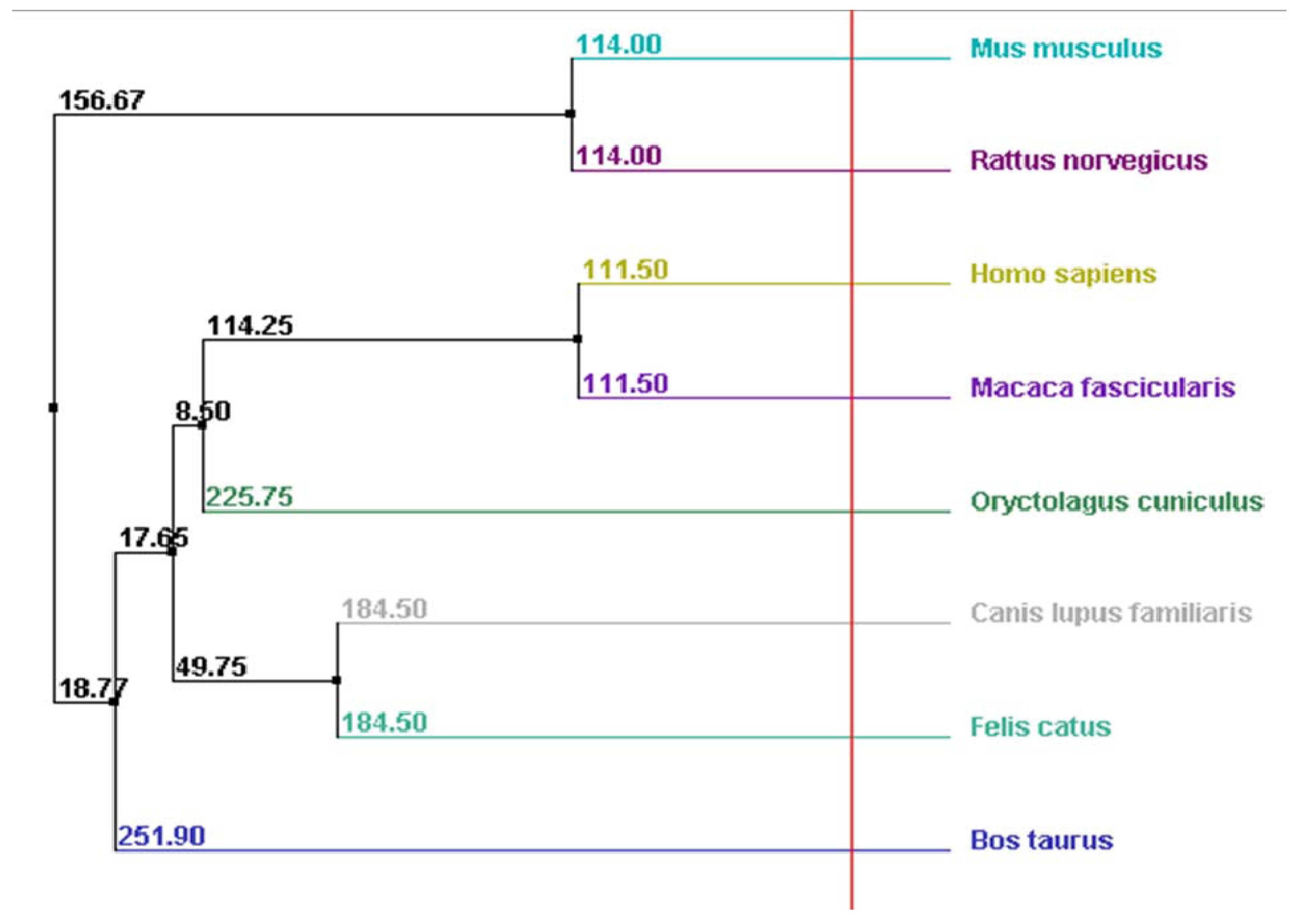
2.4. Myocilin Is Phylogenetically Well Conserved


2.5. Putative 3D-Structure of the Myocilin

3. Conclusions
Acknowledgements
References and Note
- Flammer, J.; Mozaffarieh, M. What is the present pathogenetic concept of glaucomatous optic neuropathy? Surv. Ophthalmol. 2007, 52, 162–173. [Google Scholar] [CrossRef]
- Morrison, J.C.; Johnson, E.C.; Cepurna, W.; Jia, L. Understanding mechanisms of pressure-induced optic nerve damage. Prog. Retin. Eye Res. 2005, 24, 217–240. [Google Scholar] [CrossRef]
- Quigley, H.A. Neuronal death in glaucoma. Prog. Retin. Eye Res. 1999, 18, 39–57. [Google Scholar] [CrossRef]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef]
- Leske, M.C. Open-angle glaucoma—An epidemiologic overview. Ophthalmic. Epidemiol. 2007, 14, 166–172. [Google Scholar] [CrossRef]
- Libby, R.T.; Gould, D.B.; Anderson, M.G.; John, S.W.M. Complex genetics of glaucoma susceptibility. Rev. Genomics Hum. Genet. 2005, 6, 15–44. [Google Scholar] [CrossRef]
- Leske, M.C. Ocular perfusion pressure and glaucoma: clinical trial and epidemiologic findings. Curr. Opin. Ophthalmol. 2009, 20, 73–78. [Google Scholar] [CrossRef]
- Tamm, E.R. The trabecular meshwork outflow pathways: Structural and functional aspects. Exp. Eye Res. 2009, 88, 648–655. [Google Scholar] [CrossRef]
- Tektas, O.Y.; Lütjen-Drecoll, E. Structural changes of the trabecular meshwork in different kinds of glaucoma. Exp. Eye Res. 2009, 88, 769–775. [Google Scholar] [CrossRef]
- Gasiorowski, J.Z.; Russell, P. Biological properties of trabecular meshwork cells. Exp. Eye Res. 2009, 88, 671–675. [Google Scholar] [CrossRef]
- Polansky, J.R.; Fauss, D.J.; Chen, P.; Chen, H.; Lütjen-Drecoll, E.; Johnson, D.; Kurtz, R.M.; Ma, Z.D.; Bloom, E.; Nguyen, T.D. Cellular pharmacology and molecular biology of the trabecular meshwork inducible glucocorticoid response gene product. Ophthalmologica 1997, 211, 126–139. [Google Scholar] [CrossRef]
- Stone, E.M.; Fingert, J.H.; Alward, W.L.M.; Nguyen, T.D.; Polansky, J.R.; Sunden, S.L.F.; Nishimura, D.; Clark, A.F.; Nystuen, A.; Nichols, B.E.; et al. Identification of a gene that causes primary open angle glaucoma. Science 1997, 275, 668–670. [Google Scholar] [CrossRef]
- Detry-Morel, M. Currents on target intraocular pressure and intraocular pressure fluctuations in glaucoma management. Bull. Soc. Belge Ophtalmol. 2008, 308, 35–43. [Google Scholar]
- Clement, C.I.; Goldberg, I. The management of complicated glaucoma. Indian J. Ophthalmol. 2011, 59, 141–147. [Google Scholar] [CrossRef]
- Fan, B.J.; Wiggs, J.L. Glaucoma: Genes, phenotypes, and new directions for therapy. J. Clin. Invest. 2010, 120, 3064–3072. [Google Scholar] [CrossRef]
- Shimizu, S.; Lichter, P.R.; Johnson, A.T.; Zhou, Z.; Higashi, M.; Gottfredsdottir, M.; Othman, M.; Moroi, S.E.; Rozsa, F.W.; Schertzer, R.M.; et al. Age-dependent prevalence of mutations at the GLC1A locus in primary open-angle glaucoma. Am. J. Ophthalmol. 2000, 130, 165–177. [Google Scholar] [CrossRef]
- Gobeil, S.; Rodrigue, M.A.; Moisan, S.; Nguyen, T.D.; Polansky, J.R.; Morissette, J.; Raymond, V. Intracellular sequestration of hetero-oligomers formed by wild-type and glaucoma-causing myocilin mutants. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3560–3567. [Google Scholar]
- Jacobson, N.; Andrews, M.; Shepard, A.R.; Nishimura, D.; Searby, C.; Fingert, J.H.; Hageman, G.; Mullins, R.; Davidson, B.L.; Kwon, Y.H.; et al. Non-secretion of mutant proteins of the glaucoma gene myocilin in cultured trabecular meshwork cells and in aqueous humor. Hum. Mol. Genet. 2001, 10, 117–125. [Google Scholar] [CrossRef]
- Malyukova, I.; Lee, H.S.; Fariss, R.N.; Tomarev, S.I. Mutated mouse and human myocilins have similar properties and do not block general secretory pathway. Investig. Ophthalmol. Vis. Sci. 2006, 47, 206–212. [Google Scholar] [CrossRef]
- Joe, M.K.; Sohn, S.; Hur, W.; Moon, Y.; Choi, Y.R.; Kee, C. Accumulation of mutant myocilins in ER leads to ER stress and potential cytotoxicity in human trabecular meshwork cells. Biochem. Biophys. Res. Commun. 2003, 312, 592–600. [Google Scholar] [CrossRef]
- Liu, Y.; Vollrath, D. Reversal of mutant myocilin non-secretion and cell killing: Implications for glaucoma. Hum. Mol. Genet. 2004, 13, 1193–1204. [Google Scholar] [CrossRef]
- Sheffield, V.C.; Stone, E.M.; Alward, W.L.; Drack, A.V.; Johnson, A.T.; Streb, L.M.; Nichols, B.E. Genetic linkage of familial open angle glaucoma to chromosome 1q21-q31. Nat. Genet. 1993, 4, 47–50. [Google Scholar] [CrossRef]
- Ortego, J.; Escribano, J.; Coca-Prados, M. Cloning and characterization of subtracted cDNAs from a human ciliary body library encoding TIGR, a protein involved in juvenile open angle glaucoma with homology to myosin and olfactomedin. FEBS Lett. 1997, 413, 349–353. [Google Scholar] [CrossRef]
- Kubota, R.; Noda, S.; Wang, Y.; Minoshima, S.; Asakawa, S.; Kudoh, J.; Mashima, Y.; Oguchi, Y.; Shimizu, N.A. novel myosin-like protein (myocilin) expressed in the connecting cilium of the photoreceptor: Molecular cloning, tissue expression, and chromosomal mapping. Genomics 1997, 41, 360–369. [Google Scholar]
- Nguyen, T.D.; Chen, P.; Huang, W.D.; Chen, H.; Johnson, D.; Polansky, J.R. Gene structure and properties of TIGR, an olfactomedin-related glycoprotein cloned from glucocorticoid-induced trabecular meshwork cells. J. Biol. Chem. 1998, 273, 6341–6350. [Google Scholar]
- Francois, J. Cortisone et tension ocularie. Ann. Ocul. (Paris) 1954, 187, 805–816. [Google Scholar]
- Fingert, J.H.; Clark, A.F.; Craig, J.E.; Alward, W.L.; Snibson, G.R.; McLaughlin, M.; Tuttle, L.; Mackey, D.A.; Sheffield, V.C.; Stone, E.M. Evaluation of the myocilin (MYOC) glaucoma gene in monkey and human steroid-induced ocular hypertension. Invest. Ophthalmol. Vis. Sci. 2001, 42, 145–152. [Google Scholar]
- Tomarev, S.I.; Wistow, G.; Raymond, V.; Dubois, S.; Malyukova, I. Gene expression profile of the human trabecular meshwork: NEIBank sequence tag analysis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2588–2596. [Google Scholar] [CrossRef]
- Karali, A.; Russell, P.; Stefani, F.H.; Tamm, E.R. Localization of myocilin/trabecular meshwork-inducible glucocorticoid response protein in the human eye. Investig. Ophthalmol. Vis. Sci. 2000, 41, 729–740. [Google Scholar]
- Wistow, G.; Berstein, S.L.; Ray, S.; Wyatt, M.K.; Behal, A.; Touchman, J.W.; Bouffard, G.; Smith, D.; Peterson, K. Expressed sequence tag analysis of adult human iris for the NEIBank Project: Steroid-response factors and similarities with retinal pigment epithelium. Mol. Vis. 2002, 8, 185–195. [Google Scholar]
- Wistow, G.; Berstein, S.L.; Wyatt, M.K.; Fariss, R.N.; Behal, A.; Touchman, J.W.; Bouffard, G.; Smith, D.; Peterson, K. Expressed sequence tag analysis of human RPE/choroid for the NEIBank Project: Over 6000 non-redundant transcripts, novel genes and splice variants. Mol. Vis. 2002, 8, 205–220. [Google Scholar]
- Swiderski, R.E.; Ross, J.L.; Fingert, J.H.; Clark, A.F.; Alward, W.L.; Stone, E.M.; Sheffield, V.C. Localization of MYOC transcripts in human eye and optic nerve by in situ hybridization. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3420–3428. [Google Scholar]
- Rao, P.V.; Allingham, R.R.; Epstein, D.L. TIGR/myocilin in human aqueous humor. Exp. Eye Res. 2000, 71, 637–641. [Google Scholar] [CrossRef]
- Russell, P.; Tamm, E.R.; Grehn, F.J.; Picht, G.; Johnson, M. The presence and properties of myocilin in the aqueous humor. Investig. Ophthalmol. Vis. Sci. 2001, 42, 983–986. [Google Scholar]
- Kong, T.H. Post-transcriptional modification of the gene genetically linked to juvenile open-angle glaucoma: novel transcripts in human ocular tissues. Gene 2001, 280, 115–122. [Google Scholar] [CrossRef]
- Joe, M.K.; Sohn, S.; Kim, T.E.; Im, J.E.; Choi, Y.R.; Kee, C. Analysis of glucocorticoid-induced MYOC expression in human trabecular meshwork cells. Vis. Res. 2011, 51, 1033–1038. [Google Scholar] [CrossRef]
- Park, B.C.; Tibudan, M.; Samaraweera, M.; Shen, X.; Yue, B.Y. Interaction between two glaucoma genes, optineurin and myocilin. Genes Cells 2007, 12, 969–979. [Google Scholar] [CrossRef]
- Fingert, J.H.; Ying, L.; Swiderski, R.E.; Nystuen, A.M.; Arbour, N.C.; Alward, W.L.M.; Sheffield, V.C.; Stone, E.M. Characterization and comparison of the human and mouse GLC1A glaucoma genes. Genome Res. 1998, 8, 377–384. [Google Scholar]
- Tomarev, S.I.; Tamm, E.R.; Chang, B. Characterization of the mouse Myoc/Tigr gene. Biochem. Biophys. Res. Commun. 1998, 245, 887–893. [Google Scholar] [CrossRef]
- Kubota, R.; Kudoh, J.; Mashima, Y.; Asakawa, S.; Minoshima, S.; Hejtmancik, J.F.; Oguchi, Y.; Shimizu, N. Genomic organization of the human myocilin gene (MYOC) responsible for primary open angle glaucoma (GLC1A). Biochem. Biophys. Res. Commun. 1998, 242, 396–400. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Talukdar, S.; Bhattacharjee, A.; Ray, K. Bioinformatic approaches for identification and characterization of olfactomedin related genes with a potential role in pathogenesis of ocular disorders. Mol. Vis. 2004, 10, 304–314. [Google Scholar]
- Yokoe, H.; Anholt, R.R. Molecular cloning of olfactomedin, an extracellular matrix protein specific to olfactory neuroepithelium. Proc. Natl. Acad. Sci. USA 1993, 90, 4655–4659. [Google Scholar] [CrossRef]
- Zeng, L.C.; Han, Z.G.; Ma, W.J. Elucidation of subfamily segregation and intramolecular coevolution of the olfactomedin-like proteins by comprehensive phylogenetic analysis and gene expression pattern assessment. FEBS Lett. 2005, 579, 5443–5453. [Google Scholar] [CrossRef]
- Snyder, D.A.; Rivers, A.M.; Yokoe, H.; Menco, B.P.; Anholt, R.R. Olfactomedin: Purification, characterization, and localization of a novel olfactory glycoprotein. Biochemistry 1991, 30, 9143–9153. [Google Scholar] [CrossRef]
- Barembaum, M.; Moreno, T.A.; LaBonne, C.; Sechrist, J.; Bronner-Fraser, M. Noelin-1 is a secreted glycoprotein involved in generation of the neural crest. Nat. Cell Biol. 2000, 2, 219–225. [Google Scholar] [CrossRef]
- Danielson, P.E.; Forss-Petter, S.; Battenberg, E.L.; deLecea, L.; Bloom, F.E.; Sutcliffe, J.G. Four structurally distinct neuron-specific olfactomedin-related glycoproteins produced by differential promoter utilization and alternative mRNA splicing from a single gene. J. Neurosci. Res. 1994, 38, 468–478. [Google Scholar] [CrossRef]
- Graveel, C.R.; Harkins-Perry, S.R.; Acevedo, L.G.; Farnham, P.J. Identification and characterization of CRG-L2, a new marker for liver tumor development. Oncogene 2003, 22, 1730–1736. [Google Scholar] [CrossRef]
- Hillier, B.J.; Vacquier, V.D. Amassin, an olfactomedin protein, mediates the massive intercellular adhesion of sea urchin coelomocytes. J. Cell Biol. 2003, 160, 597–604. [Google Scholar] [CrossRef]
- Kulkarni, N.H.; Karavanich, C.A.; Atchley, W.R.; Anholt, R.R. Characterization and differential expression of a human gene family of olfactomedin-related proteins. Genet. Res. 2000, 76, 41–50. [Google Scholar] [CrossRef]
- Loria, P.M.; Hodgkin, J.; Hobert, O. A conserved postsynaptic transmembrane protein affecting neuromuscular signaling in Caenorhabditis elegans. J. Neurosci. 2004, 24, 2191–2201. [Google Scholar] [CrossRef]
- Nakaya, N.; Lee, H.S.; Takada, Y.; Tzchori, I.; Tomarev, S.I. Zebrafish olfactomedin 1 regulates retinal axon elongation in vivo and is a modulator of Wnt signaling pathway. J. Neurosci. 2008, 28, 7900–7910. [Google Scholar] [CrossRef]
- Torrado, M.; Trivedi, R.; Zinovieva, R.; Karavanova, I.; Tomarev, S.I. Optimedin: A novel olfactomedin-related protein that interacts with myocilin. Hum. Mol. Genet. 2002, 11, 1291–1301. [Google Scholar] [CrossRef]
- Tsuda, H.; Sasai, N.; Matsuo-Takasaki, M.; Sakuragi, M.; Murakami, Y.; Sasai, Y. Dorsalization of the neural tube by Xenopus tiarin, a novel patterning factor secreted by the flanking nonneural head ectoderm. Neuron 2002, 33, 515–528. [Google Scholar] [CrossRef]
- Aroca-Aguilar, J.D.; Sanchez-Sanchez, F.; Ghosh, S.; Coca-Prados, M.; Escribano, J. Myocilin mutations causing glaucoma inhibit the intracellular endoproteolytic cleavage of myocilin between amino acids arg226 and ile227. J. Biol. Chem. 2005, 280, 21043–21051. [Google Scholar]
- Tamm, E.R. Myocilin and glaucoma: Facts and ideas. Prog. Retin. Eye Res. 2002, 21, 395–428. [Google Scholar] [CrossRef]
- Ortego, J.; Escribano, J.; Coca-Prados, M. Cloning and characterization of subtracted cDNAs from a human ciliary body library encoding TIGR, a protein involved in juvenile open angle glaucoma with homology to myosin and olfactomedin. FEBS Lett. 1997, 413, 349–353. [Google Scholar] [CrossRef]
- Gobeil, S.; Letartre, L.; Raymond, V. Functional analysis of the glaucoma-causing TIGR/myocilin protein: integrity of amino-terminal coiled-coil regions and olfactomedin homology domain is essential for extracellular adhesion and secretion. Exp. Eye Res. 2006, 82, 1017–1029. [Google Scholar] [CrossRef]
- Burns, J.N.; Orwig, S.D.; Harris, J.L.; Watkins, J.D.; Vollrath, D.; Lieberman, R.L. Rescue of glaucoma-causing mutant myocilin thermal stability by chemical chaperones. ACS Chem. Biol. 2010, 5, 477–487. [Google Scholar] [CrossRef]
- Fingert, J.H.; Stone, E.M.; Sheffield, V.C.; Alward, W.L. Myocilin glaucoma. Surv. Ophthalmol. 2002, 47, 547–561. [Google Scholar] [CrossRef]
- Kim, B.S.; Savinova, O.V.; Reedy, M.V.; Martin, J.; Lun, Y.; Gan, L.; Smith, R.S.; Tomarev, S.I.; John, S.W.M.; Johnson, R.L. Targeted disruption of the myocilin gene (Myoc) suggests that human glaucoma-causing mutations are gain of function. Mol. Cell. Biol. 2001, 21, 7707–7713. [Google Scholar] [CrossRef]
- Gould, D.B.; Miceli-Libby, L.; Savinova, O.V.; Torrado, M.; Tomarev, S.I.; Smith, R.S.; John, S.W.M. Genetically increasing Myoc expression supports a necessary pathologic role of abnormal proteins in glaucoma. Mol. Cell. Biol. 2004, 24, 9019–9025. [Google Scholar] [CrossRef]
- Wiggs, J.L.; Vollrath, D. Molecular and clinical evaluation of a patient hemizygous for TIGR/MYOC. Arch. Ophthal. 2001, 119, 1674–1678. [Google Scholar] [CrossRef]
- Lam, D.S.; Leung, Y.F.; Chua, J.K.; Baum, L.; Fan, D.S.; Choy, K.W.; Pang, C.P. Truncations in the TIGR gene in individuals with and without primary open-angle glaucoma. Investig. Ophthalmol. Vis. Sci. 2000, 4, 1386–1391. [Google Scholar]
- Paper, W.; Kroeber, M.; Heersink, S.; Stephan, D.A.; Fuchshofer, R.; Russell, P.; Tamm, E.R. Elevated amounts of myocilin in the aqueous humor of transgenic mice cause significant changes in ocular gene expression. Exp. Eye Res. 2008, 87, 257–267. [Google Scholar] [CrossRef]
- Fautsch, M.P.; Vrabel, A.M.; Johnson, D.H. The identification of myocilin-associated proteins in the human trabecular meshwork. Exp. Eye Res. 2006, 82, 1046–1052. [Google Scholar] [CrossRef]
- Joe, M.K.; Sohn, S.; Choi, Y.R.; Park, H.; Kee, C. Identification of flotillin-1 as a protein interacting with myocilin: Implications for the pathogenesis of primary open-angle glaucoma. Biochem. Biophys. Res. Commun. 2005, 336, 1201–1206. [Google Scholar] [CrossRef]
- Li, Y.; Aroca-Aguilar, J.D.; Ghosh, S.; Sánchez-Sánchez, F.; Escribano, J.; Coca-Prados, M. Interaction of myocilin with the C-terminal region of hevin. Biochem. Biophys. Res. Commun. 2006, 339, 797–804. [Google Scholar] [CrossRef]
- Peters, D.M.; Herbert, K.; Biddick, B.; Peterson, J.A. Myocilin binding to Hep II domain of fibronectin inhibits cell spreading and incorporation of paxillin into focal adhesions. Exp. Cell Res. 2005, 303, 218–228. [Google Scholar] [CrossRef]
- Surgucheva, I.; Park, B.C.; Yue, B.Y.; Tomarev, S.; Surguchov, A. Interaction of myocilin with gamma-synuclein affects its secretion and aggregation. Cell. Mol. Neurobiol. 2005, 25, 1009–1033. [Google Scholar] [CrossRef]
- Kwon, H.S.; Lee, H.S.; Ji, Y.; Rubin, J.S.; Tomarev, S.I. Myocilin is a modulator of Wnt signaling. Mol. Cell. Biol. 2009, 29, 2139–2154. [Google Scholar] [CrossRef]
- Wentz-Hunter, K.; Ueda, J.; Yue, B.Y. Protein interactions with myocilin. Invest. Ophthalmol. Vis. Sci. 2002, 43, 176–182. [Google Scholar]
- Copin, B.; Brézin, A.P.; Valtot, F.; Dascotte, J.C.; Béchetoille, A.; Garchon, H.J. Apolipoprotein E-promoter single-nucleotide polymorphisms affect the phenotype of primary open-angle glaucoma and demonstrate interaction with the myocilin gene. Am. J. Hum. Genet. 2002, 70, 1575–1581. [Google Scholar] [CrossRef]
- Ueda, J.; Yue, B.Y. Distribution of myocilin and extracellular matrix components in the corneoscleral meshwork of human eyes. Invest. Ophthalmol. Vis. Sci. 2003, 44, 4772–4779. [Google Scholar] [CrossRef]
- Fautsch, M.P.; Johnson, D.H. Characterization of myocilin-myocilin interactions. Invest. Ophthalmol. Vis. Sci. 2001, 42, 2324–2331. [Google Scholar]
- Lo, W.R.; Rowlette, L.L.; Caballero, M.; Yang, P.; Hernandez, M.R.; Borras, T. Tissue differential microarray analysis of dexamethasone induction reveals potential mechanisms of steroid glaucoma. Invest. Ophthal. Vis. Sci. 2003, 44, 473–485. [Google Scholar]
- Liton, P.B.; Luna, C.; Challa, P.; Epstein, D.L.; Gonzalez, P. Genome-wide expression profile of human trabecular meshwork cultured cells, non glaucomatous and primary open angle glaucoma tissue. Mol. Vis. 2006, 12, 774–790. [Google Scholar]
- Tomarev, S.I.; Nakaya, N. Olfactomedin domain-containing proteins: Possible mechanisms of action and functions in normal development and pathology. Mol. Neurobiol. 2009, 40, 122–138. [Google Scholar] [CrossRef]
- Kwon, Y.H.; Fingert, J.H.; Kuehn, M.H.; Alward, W.L.M. Primary Open-Angle Glaucoma. N. Engl. J. Med. 2009, 360, 1113–1124. [Google Scholar] [CrossRef]
- Zhou, Z.; Vollrath, D. A cellular assay distinguishes normal and mutant TIGR/myocilin protein. Hum. Mol. Genet. 1999, 8, 2221–2228. [Google Scholar] [CrossRef]
- Rangachari, K.; Dhivya, M.; Eswari Pandaranayaka, P.J.; Prasanthi, N.; Sundaresan, P.; Krishnadas, S.R.; Krishnaswamy, S. Glaucoma database. Bioinformation 2011, 5, 398–399. [Google Scholar] [CrossRef]
- Adam, M.F.; Belmouden, A.; Binisti, P.; Brezin, A.P.; Valtot, F.; Bechetoille, A.; Dascot, J.-C.; Copin, B.; Gomez, L.; Chaventre, A.; et al. Recurrent mutations in a single exon encoding the evolutionarily conserved olfactomedin-homology domain of TIGR in familial open-angle glaucoma. Hum. Molec. Genet. 1997, 6, 2091–2097. [Google Scholar] [CrossRef]
- Colomb, E.; Nguyen, T.D.; Bechetoille, A.; Dascotte, J.-C.; Valtot, F.; Brezin, A.P.; Berkani, M.; Copin, B.; Gomez, L.; Polansky, J.R.; et al. Association of a single nucleotide polymorphism in the TIGR/MYOCILIN gene promoter with the severity of primary open-angle glaucoma. Clin. Genet. 2001, 60, 220–225. [Google Scholar]
- Polansky, J.R.; Juster, R.P.; Spaeth, G.L. Association of the myocilin mt.1 promoter variant with the worsening of glaucomatous disease over time. Clin. Genet. 2003, 64, 18–27. [Google Scholar] [CrossRef]
- Saura, M.; Cabana, M.; Ayuso, C.; Valverde, D. Mutations including the promoter region of myocilin/TIGR gene. Eur. J. Hum. Genet. 2005, 13, 384–387. [Google Scholar] [CrossRef]
- Kasahara, N.; Caixeta-Umbelino, C.; Paolera, M.D.; Rocha, M.N.; Richeti, F.; Vasconcellos, J.P.; Cohen, R.; Costa, V.P.; Longui, C.A.; Melo, M.R.; et al. Myocilin mt.1 gene promoter single nucleotide polymorphism (-1000C > G) in Brazilian patients with primary open angle glaucoma. Ophthal. Genet. 2011, 32, 18–23. [Google Scholar] [CrossRef]
- Faucher, M.; Anctil, J.-L.; Rodrigue, M.-A.; Duchesne, A.; Bergeron, D.; Blondeau, P.; Cote, G.; Dubois, S.; Bergeron, J.; Arseneault, R.; et al. Founder TIGR/myocilin mutations for glaucoma in the Quebec population. Hum. Mol. Genet. 2002, 11, 2077–2090. [Google Scholar] [CrossRef]
- Fingert, J.H.; Heon, E.; Liebmann, J.M.; Yamamoto, T.; Craig, J.E.; Rait, J.; Kawase, K.; Hoh, S.-T.; Buys, Y.M.; Dickinson, J.; et al. Analysis of myocilin mutations in 1703 glaucoma patients from five different populations. Hum. Mol. Genet. 1999, 8, 899–905. [Google Scholar] [CrossRef]
- De Vasconcellos, J.P.; de Melo, M.B.; Schimiti, R.; Costa, F.F.; Costa, V.P. Penetrance and phenotype of the Cys433Arg myocilin mutation in a family pedigree with primary open-angle glaucoma. J. Glaucoma 2003, 12, 104–107. [Google Scholar] [CrossRef]
- Vincent, A.L.; Billingsley, G.; Buys, Y.; Levin, A.V.; Priston, M.; Trope, G.; Williams-Lyn, D.; Heon, E. Digenic inheritance of early-onset glaucoma: CYP1B1, a potential modifier gene. Am. J. Hum. Genet. 2002, 70, 448–460. [Google Scholar] [CrossRef]
- Yam, G.H.-F.; Zuber, C.; Roth, J. A synthetic chaperone corrects the trafficking defect and disease phenotype in a protein misfolding disorder. FASEB J. 2005, 19, 12–18. [Google Scholar] [CrossRef]
- Yam, G.H.; Gaplovska-Kysela, K.; Zuber, C.; Roth, J. Aggregated myocilin induces russell bodies and causes apoptosis: implications for the pathogenesis of myocilin-caused primary open-angle glaucoma. Am. J. Pathol. 2007, 170, 100–109. [Google Scholar] [CrossRef]
- Aroca-Aguilar, J.D.; Sánchez-Sánchez, F.; Ghosh, S.; Fernández-Navarro, A.; Coca-Prados, M.; Escribano, J. Interaction of Recombinant Myocilin with the Matricellular Protein SPARC: Functional Implications. Invest. Ophthalmol. Vis. Sci. 2011, 52, 179–189. [Google Scholar] [CrossRef]
- Sánchez-Sánchez, F.; Martínez-Redondo, F.; Aroca-Aguilar, J.D.; Coca-Prados, M.; Escribano, J. Characterization of the intracellular proteolytic cleavage of myocilin and identification of calpain II as a myocilin-processing protease. J. Biol. Chem. 2007, 282, 27810–27824. [Google Scholar]
- Aroca-Aguilar, J.D.; Martínez-Redondo, F.; Sánchez-Sánchez, F.; Coca-Prados, M.; Escribano, J. Functional role of proteolytic processing of recombinant myocilin in self-aggregation. Invest. Ophthalmol. Vis. Sci. 2010, 51, 72–78. [Google Scholar] [CrossRef]
- Jia, L.-Y.; Gong, B.; Pang, C.-P.; Huang, Y.; Lam, D.S.-C.; Wang, N.; Yam, G.H.-F. Correction of the disease phenotype of myocilin-causing glaucoma by a natural osmolyte. Invest. Ophthalmol. Vis. Sci. 2009, 50, 3743–3749. [Google Scholar]
- Yam, G.H.-F.; Gaplovska-Kysela, K.; Zuber, C.; Roth, J. Sodium 4-phenylbutyrate acts as a chemical chaperone on misfolded myocilin to rescue cells from endoplasmic reticulum stress and apoptosis. Invest. Ophthalmol. Vis. Sci. 2007, 48, 1683–1690. [Google Scholar] [CrossRef]
- Menaa, B.; Menaa, F.; Aiolfi-Guimarães, C.; Sharts, O. Silica-based nanoporous sol-gel glasses: From bioencapsulation to protein folding studies. Int. J. Nanotechnol. 2010, 7, 1–45. [Google Scholar] [CrossRef]
- Menaa, B.; Montoneri, C.; Menaa, F.; Montoneri, E.; Boffa, V.; Sharts, N. Protein helical structure enhancement in biocompatible fluoro-phosphonate-based nanoporous silica glasses assessed by circular dichroism spectroscopy. Int. J. Nanotechnol. 2011, 8, 471–491. [Google Scholar]
- Menaa, B.; Miyagawa, Y.; Takahashi, M.; Herrero, M.; Rives, V.; Menaa, F.; Eggers, D.K. Bioencapsulation of apomyoglobin in nanoporous organosilica sol-gel glasses: Influence of the siloxane network on the conformation and stability of a model protein. Biopolymers 2009, 91, 895–906. [Google Scholar] [CrossRef]
- Orwig, S.D.; Lieberman, R.L. Biophysical characterization of the olfactomedin domain of myocilin, an extracellular matrix protein implicated in inherited forms of glaucoma. PLoS One 2011, 6, e16347. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acid. Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Higgins, D.G.; Thompson, J.D.; Gibson, T.J. Using CLUSTAL for multiple sequence alignments. Methods Enzymol. 1996, 266, 383–402. [Google Scholar]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER: Fully automated protein structure prediction in CASP8. Proteins 2009, S9, 100–113. [Google Scholar] [CrossRef]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef]
- Bluhm, W.F.; Beran, B.; Bi, C.; Dimitropoulos, D.; Prlic, A.; Quinn, G.B.; Rose, P.W.; Shah, C.; Young, J.; Yukich, B.; et al. Quality assurance for the query and distribution systems of the RCSB Protein Data Bank. Database (Oxford) 2011, 2011. Article ID bar003. [Google Scholar]
- Rose, P.W.; Beran, B.; Bi, C.; Bluhm, W.F.; Dimitropoulos, D.; Goodsell, D.S.; Prlic, A.; Quesada, M.; Quinn, G.B.; Westbrook, J.D.; et al. The RCSB Protein Data Bank: redesigned web site and web services. Nucl. Acid. Res. 2011, Database, D392–D401. [Google Scholar]
- Suzuki, Y.; Noma, A.; Suzuki, T.; Ishitani, R.; Nureki, O. Structural basis of tRNA modification with CO2 fixation and methylation by wybutosine synthesizing enzyme TYW4. Nucl. Acid. Res. 2009, 37, 2910–2925. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Menaa, F.; Braghini, C.A.; Vasconcellos, J.P.C.D.; Menaa, B.; Costa, V.P.; Figueiredo, E.S.D.; Melo, M.B.D. Keeping an Eye on Myocilin: A Complex Molecule Associated with Primary Open-Angle Glaucoma Susceptibility. Molecules 2011, 16, 5402-5421. https://doi.org/10.3390/molecules16075402
Menaa F, Braghini CA, Vasconcellos JPCD, Menaa B, Costa VP, Figueiredo ESD, Melo MBD. Keeping an Eye on Myocilin: A Complex Molecule Associated with Primary Open-Angle Glaucoma Susceptibility. Molecules. 2011; 16(7):5402-5421. https://doi.org/10.3390/molecules16075402
Chicago/Turabian StyleMenaa, Farid, Carolina Ayumi Braghini, Jose Paulo Cabral De Vasconcellos, Bouzid Menaa, Vital Paulino Costa, Eugênio Santana De Figueiredo, and Monica Barbosa De Melo. 2011. "Keeping an Eye on Myocilin: A Complex Molecule Associated with Primary Open-Angle Glaucoma Susceptibility" Molecules 16, no. 7: 5402-5421. https://doi.org/10.3390/molecules16075402
APA StyleMenaa, F., Braghini, C. A., Vasconcellos, J. P. C. D., Menaa, B., Costa, V. P., Figueiredo, E. S. D., & Melo, M. B. D. (2011). Keeping an Eye on Myocilin: A Complex Molecule Associated with Primary Open-Angle Glaucoma Susceptibility. Molecules, 16(7), 5402-5421. https://doi.org/10.3390/molecules16075402