The Use of Bio-Guided Fractionation to Explore the Use of Leftover Biomass in Dutch Flower Bulb Production as Allelochemicals against Weeds


 and
and
Abstract
:
1. Introduction
2. Results and Discussion

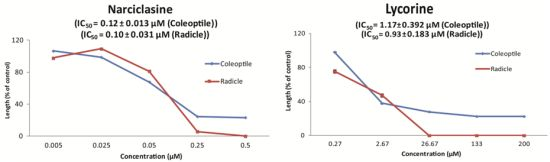{kind=link}
{kind=link}
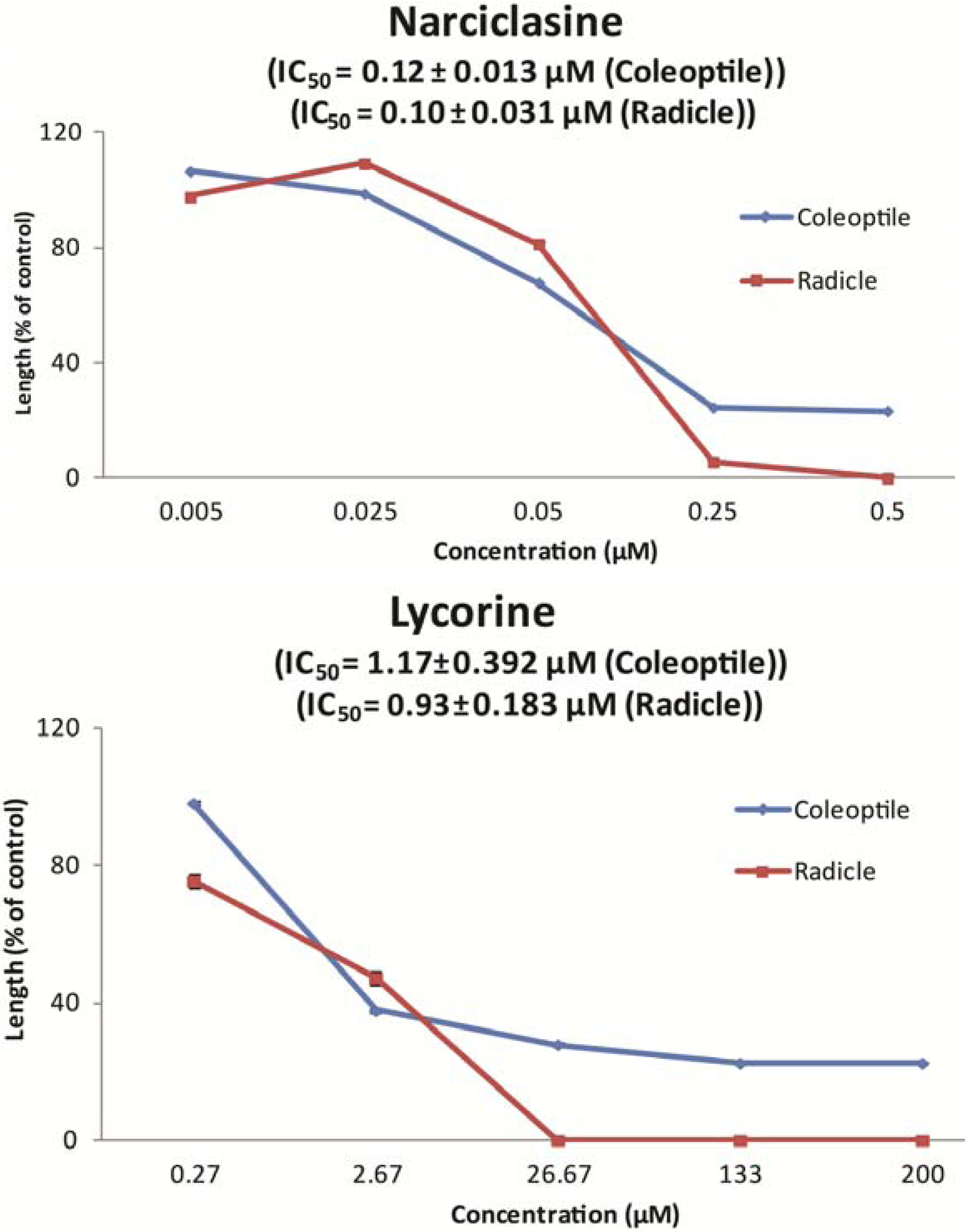{kind=link}
{kind=link}
{kind=link}
| No. | Scientific Name | Family | Part of plant | Germination inhibition of L. perenne(%) | |||
|---|---|---|---|---|---|---|---|
| Methanol Extracts | Chloroform Extracts | ||||||
| 1 mg/mL | 0.5 mg/mL | 1 mg/mL | 0.5 mg/mL | ||||
| 1. | Narcissus pseudonarcissus cv. Carlton | Amaryllidaceae | Bulb | 96.7 *** | 96.7 *** | 100.0 *** | 100.0 *** |
| 2. | Narcissus tazetta cv. Grand soleil d’or | Amaryllidaceae | Bulb | 86.7 *** | 56.7 *** | 16.7 | 16.7 |
| 3. | Narcissus tazetta cv. Avalanche | Amaryllidaceae | Bulb | 86.7 *** | 53.3 *** | 23.3 | 6.7 |
| 4. | Hippeastrum Hybrid cv. Misty | Amaryllidaceae | Bulb | 76.7 *** | 90.0 *** | 6.7 | 10.0 |
| 5. | Narcissus poeticus cv. Recurvus | Amaryllidaceae | Bulb | 99.7 *** | 86.7 *** | 20.0 | 3.3 |
| 6. | Hippeastrum cybister cv. Chico | Amaryllidaceae | Bulb | 86.7 *** | 53.3 *** | 3.3 | 16.7 |
| 7. | Crinum asiaticum cv. Sandarium | Amaryllidaceae | Bulb | 20.0 | 3.3 | 93.3 *** | 30.0 |
| 8. | Amaryllis belladonna | Amaryllidaceae | Bulb | 30.0 | 10.0 | 00.0 | 3.3 |
| 9. | Nerine sarniensis | Amaryllidaceae | Bulb | 26.7 | 16.7 | 3.3 | 3.3 |
| 10. | Hymenocallis festalis cv. Zwanenburg | Amaryllidaceae | Leaf | 10.0 | 10.0 | 6.7 | 10.0 |
| 11. | Gladiolus cv. Rose Supreme | Iridaceae | Flower | 20.0 | 6.7 | 3.3 | 6.7 |
| 12. | Arisaema cv. Tortuosum | Araceae | Bulb | 20.0 | 6.7 | 3.3 | 3.3 |
| 13. | Anemone Coronaria cv. De Caen | Ranunculaceae | Leaf | 6.7 | 6.7 | 16.7 | 13.3 |
| 14. | Allium spherocephalon | Amaryllidaceae | Flower | 6.7 | 3.3 | 16.7 | 3.3 |
| 15. | Oxalis triangularis | Oxalidaceae | Flower | 13.3 | 10.0 | 10.0 | 6.7 |
| 16. | Scilla siberica cv. Spring Beauty | Asparagaceae | Flower | 3.3 | 10.0 | 3.3 | 13.3 |
| 17. | Ornithogalum thyrsoides cv. Mount Fuji | Asparagaceae | Leaf and stem | 6.7 | 20.0 | 10.0 | 10.0 |
| 18. | Hyacinthus orientalis cv. White Pearl | Asparagaceae | Leaf | 10.0 | 3.3 | 6.7 | 00.0 |
| 19. | Babiana stricta | Iridaceae | Leaf | 10.0 | 6.7 | 3.3 | 00.0 |
| 20. | Amarine tubergenii cv. Zwanenburg | Asparagaceae | Flower | 16.7 | 16.7 | 10.0 | 3.3 |
| 21. | Zephyranthes robusta | Amaryllidaceae | Flower | 10.0 | 10.0 | 0.0 | 0.0 |
| 22. | Ixia paniculata cv. Yellow Emperor | Iridaceae | Flower | 10.0 | 6.7 | 6.7 | 30.0 |
| 23. | Caladium cv. Postman Joyner | Araceae | Bulb | 3.3 | 3.3 | 6.7 | 3.3 |
| 24. | Begonia cv. Compacta | Begoniaceae | Bulb | 3.3 | 3.3 | 6.7 | 6.7 |
| 25. | Dahlia cv. Deca Split Pink Bells | Iridaceae | Bulb | 3.3 | 10.0 | 13.3 | 3.3 |
| 26. | Eucomis comosa | Liliaceae | Bulb | 3.3 | 13.3 | 6.7 | 6.7 |
| 27. | Iris X hollandica cv. Apollo | Iridaceae | Bulb | 10.0 | 3.3 | 3.3 | 3.3 |
| 28. | Incarvillea delavayi | Bignoniaceae | Leaf | 13.3 | 10.0 | 3.3 | 13.3 |
| 29. | Arisaema galeatum | Araceae | Leaf | 6.7 | 10.0 | 6.7 | 6.7 |
| 30. | Dahlia cv. Apricot Star | Iridaceae | Leaf | 3.3 | 16.7 | 13.3 | 13.3 |
| 31. | Lilium candidum | Liliaceae | Bulb | 10.0 | 13.3 | 13.3 | 3.3 |
| 32. | Hyacinthus orientalis cv. Delft Blue | Asparagaceae | Bulb | 3.3 | 10.0 | 6.7 | 16.7 |
| 33. | Fritallaria | Liliaceae | Bulb | 10.0 | 3.3 | 0.0 | 0.0 |
| No. | Scientific Name | Family | Part of plant | Germination inhibition of S. vulgaris (%) | |||
|---|---|---|---|---|---|---|---|
| Methanol Extracts | Chloroform Extracts | ||||||
| 1 mg/mL | 0.5 mg/mL | 1 mg/mL | 0.5 mg/mL | ||||
| 1. | Narcissus pseudonarcissus cv. Carlton | Amaryllidaceae | Bulb | 100.0 *** | 100.0 *** | 100.0 *** | 100.0 *** |
| 2. | Narcissus tazetta cv. Grand soleil d’or | Amaryllidaceae | Bulb | 100.0 *** | 100.0 *** | 100.0 *** | 100.0 *** |
| 3. | Narcissus tazetta cv. Avalanche | Amaryllidaceae | Bulb | 100.0 *** | 100.0 *** | 100.0 *** | 16.7 *** |
| 4. | Hippeastrum Hybrid cv. Misty | Amaryllidaceae | Bulb | 100.0 *** | 96.7 *** | 0.0 | 0.0 |
| 5. | Narcissus poeticus cv. Recurvus | Amaryllidaceae | Bulb | 100.0 *** | 90.0 *** | 3.3 | 0.0 |
| 6. | Hippeastrum cybister cv. Chico | Amaryllidaceae | Bulb | 100.0 *** | 86.7 *** | 0.0 | 0.0 |
| 7. | Crinum asiaticum cv. Sandarium | Amaryllidaceae | Bulb | 93.3 *** | 30.0 | 0.0 | 0.0 |
| 8. | Amaryllis belladonna | Amaryllidaceae | Bulb | 90.0 *** | 6.7 | 6.7 | 0.0 |
| 9. | Nerine sarniensis | Amaryllidaceae | Bulb | 83.3 *** | 0.0 | 16.7 | 0.0 |
| 10. | Hymenocallis festalis cv. Zwanenburg | Amaryllidaceae | Leaf | 43.3 ** | 0.0 | 0.0 | 0.0 |
| 11. | Gladiolus cv. Rose Supreme | Iridaceae | Flower | 0.0 | 0.0 | 0.0 | 0.0 |
| 12. | Arisaema cv.Tortuosum | Araceae | Bulb | 0.0 | 3.3 | 0.0 | 0.0 |
| 13. | Anemone Coronaria cv. De Caen | Ranunculaceae | Leave | 0.0 | 0.0 | 0.0 | 0.0 |
| 14. | Allium spherocephalon | Amaryllidaceae | Flower | 0.0 | 0.0 | 3.3 | 0.0 |
| 15. | Oxalis triangularis | Oxalidaceae | Flower | 0.0 | 0.0 | 0.0 | 0.0 |
| 16. | Scilla siberica cv. Spring Beauty | Asparagaceae | Flower | 0.0 | 0.0 | 0.0 | 0.0 |
| 17. | Ornithogalum thyrsoides cv. Mount Fuji | Asparagaceae | Leaf and stem | 0.0 | 0.0 | 0.0 | 0.0 |
| 18. | Hyacinthus orientalis cv. White Pearl | Asparagaceae | Leaf | 0.0 | 0.0 | 0.0 | 0.0 |
| 19. | Babiana stricta | Iridaceae | Leaf | 0.0 | 0.0 | 0.0 | 0.0 |
| 20. | Amarine tubergenii cv. Zwanenburg | Asparagaceae | Flower | 0.0 | 0.0 | 0.0 | 0.0 |
| 21. | Zephyranthes robusta | Amaryllidaceae | Flower | 0.0 | 0.0 | 0.0 | 0.0 |
| 22. | Ixia paniculata cv.Yellow Emperor | Iridaceae | Flower | 0.0 | 0.0 | 0.0 | 0.0 |
| 23. | Caladium cv. Postman Joyner | Araceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 24. | Begonia cv. Compacta | Begoniaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 25. | Dahlia cv. Deca Split Pink Bells | Iridaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 26. | Eucomis comosa | Liliaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 27. | Iris X hollandica cv. Apollo | Iridaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 28. | Incarvillea delavayi | Bignoniaceae | Leaf | 0.0 | 0.0 | 0.0 | 0.0 |
| 29. | Arisaema galeatum | Araceae | Leaf | 0.0 | 0.0 | 0.0 | 0.0 |
| 30. | Dahlia cv. Apricot Star | Iridaceae | Leaf | 0.0 | 0.0 | 0.0 | 0.0 |
| 31. | Lilium candidum | Liliaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 32. | Hyacinthus orientalis cv. Delft Blue | Asparagaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
| 33. | Fritallaria | Liliaceae | Bulb | 0.0 | 0.0 | 0.0 | 0.0 |
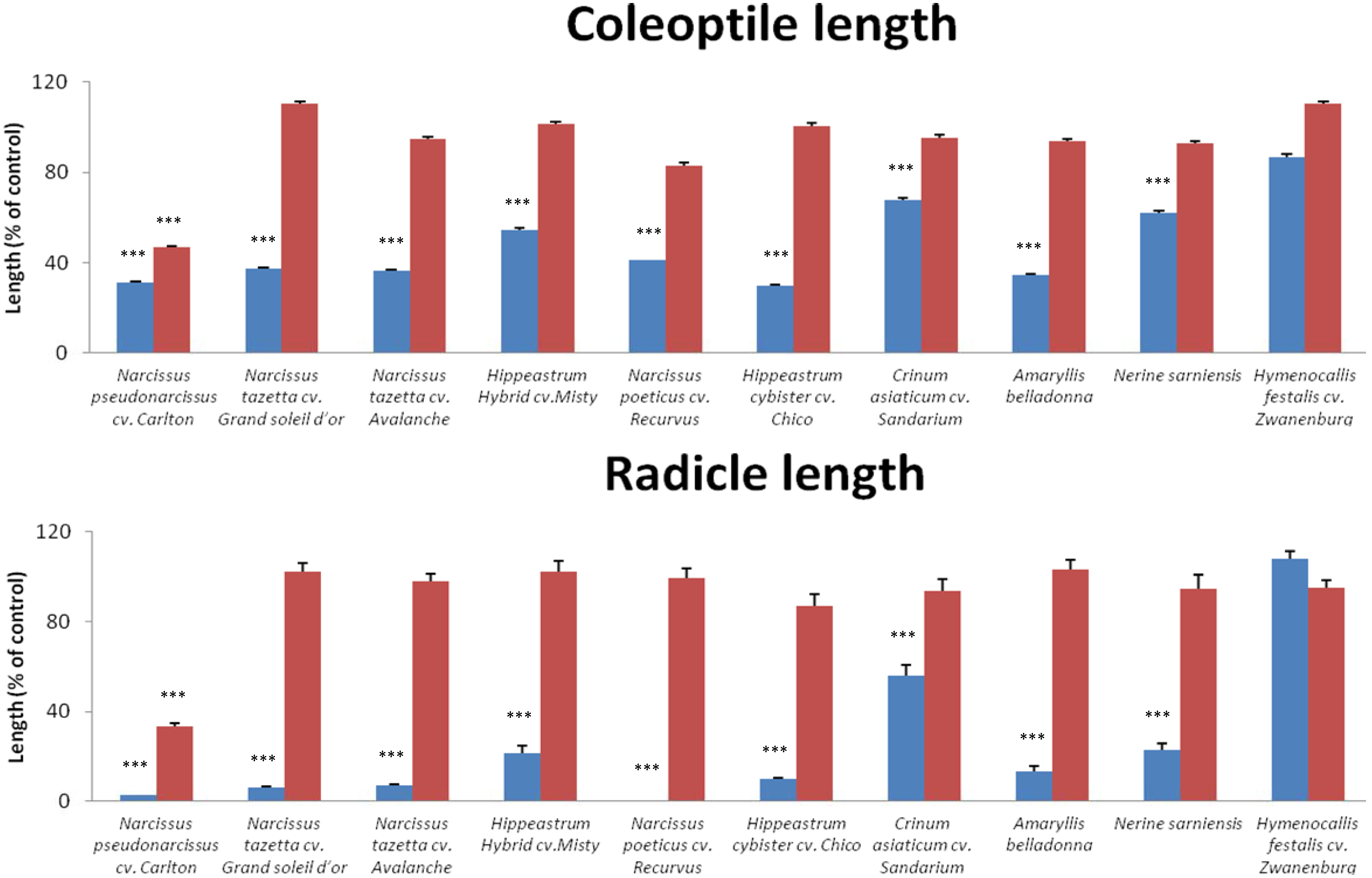
| No. | Plant name | Narciclasine | Lycorine | ||
|---|---|---|---|---|---|
| µM | mg/kgW | µM | mg/kgW | ||
| 1. | Narcissus pseudonarcissus cv. Carlton | 0.88 a | 0.27 a | nd | nd |
| 2. | Narcissus tazetta cv. Grand soleil d’or | nd | nd | 8.58 a | 2.47 a |
| 3. | Narcissus tazetta cv. Avalanche | 0.20 a | 0.06 a | 5.22 a | 1.50 a |
| 4. | Hippeastrum Hybrid cv. Misty | nd | nd | 3.26 b | 0.94 b |
| 5. | Narcissus poeticus cv. Recurvus | 0.07 b | 0.02 b | 2.14 b | 0.61 b |
| 6. | Hippeastrum cybister cv. Chico | nd | nd | nd | nd |
| 7. | Crinum asiaticum cv. Sandarium | nd | nd | 5.36 b | 1.54 b |
| 8. | Amaryllis belladonna | nd | nd | 23.21 b | 6.67 b |
| 9. | Nerine sarniensis | nd | nd | 29.43 b | 8.45 b |
| 10. | Hymenocallis festalis cv. Zwanenburg | nd | nd | nd | nd |
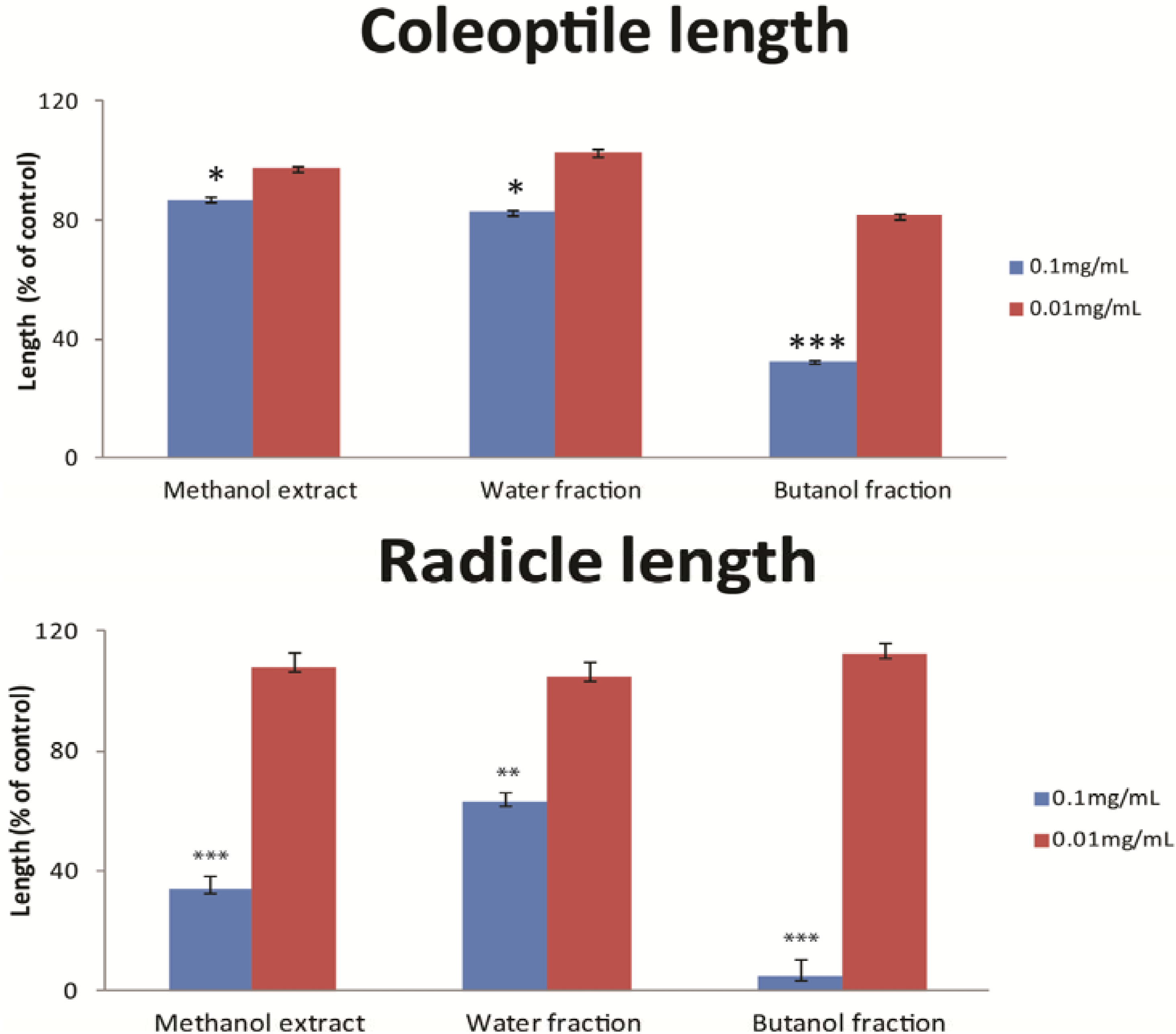
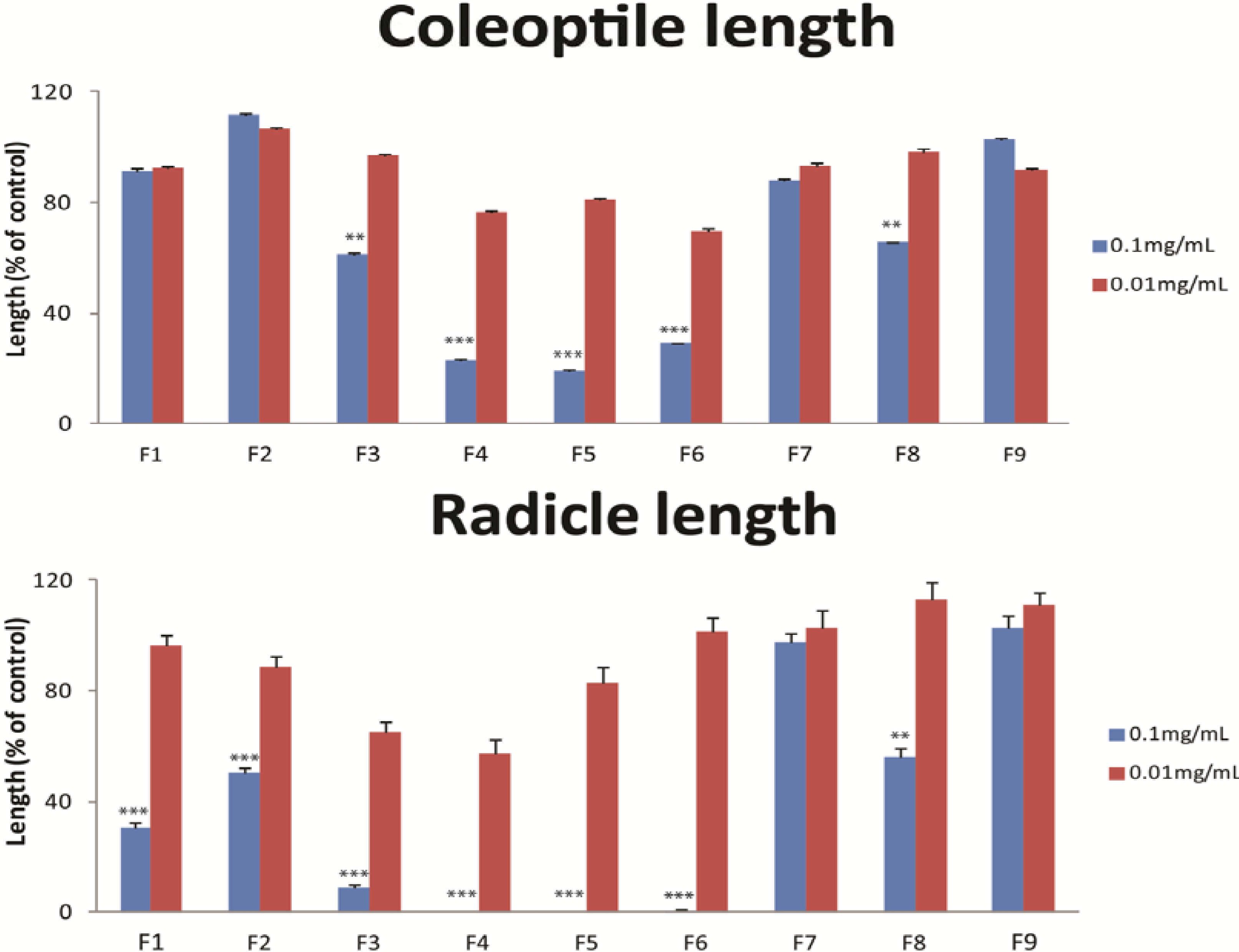
3. Experimental
3.1. Plant Materials
3.2. Chemicals
3.3. Bioassays
3.3.1. Germination Inhibition Bioassay
3.3.2. Growth Inhibition Bioassay
3.4. Identification and Purification of Active Compounds
3.4.1. Narciclasine
3.4.2. Lycorine
3.5. Growth Inhibition Activity of Narciclasine and Lycorine
4. Conclusions
Acknowledgments
References
- Benschop, M.; Kamenetsky, R.; le Nard, M.; Okubo, H.; le Hertogh, A. The Global Flower Bulb Industry: Production, Utilization, Research. In Horticultural Reviews; Wiley: Hoboken, NJ, USA, 2010; Volume 36. [Google Scholar]
- Al-Khatib, K. Tulip (Tulipa spp.), daffodil (Narcissus spp.), and iris (Iris spp.) response to preemergence herbicides. Weed Technol. 1996, 710–715. [Google Scholar]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- Lawson, H.; Wiseman, J. The effect of weeds on the growth and development of narcissus. J. Appl. Ecol. 1978, 257–272. [Google Scholar] [CrossRef]
- Lawson, H. The effects of spring-germinating weeds on narcissus. Ann. Appl. Biol. 1976, 83, 324–327. [Google Scholar] [CrossRef]
- Briggs, J. Flower bulb production. Agriculture 1970, 77, 358–363. [Google Scholar]
- Miller, T.W.; Libbey, C.R. Application of Paraquat Can Injure Field-grown Narcissus. Hort. Technol. 2010, 20, 220–223. [Google Scholar]
- Ulber, L.; Nordmeyer, H.; Zwerger, P. Resistance risk assessment within herbicide authorisation-a call for sensitivity data. Pest Manag. Sci. 2012, 69, 160–164. [Google Scholar]
- Shajib, M.T.I.; Pedersen, H.A.; Mortensen, A.G.; Kudsk, P.; Fomsgaard, I.S. Phytotoxic effect, uptake, and transformation of biochanin a in selected weed species. J. Agric. Food Chem. 2012, 60, 10715–10722. [Google Scholar] [CrossRef]
- Coelho, S. European pesticide rules promote resistance, researchers warn. Science 2009, 323, 450. [Google Scholar] [CrossRef]
- Rossing, W.A.H.; Jansma, J.E.; De Ruijter, F.J.; Schans, J. Operationalizing sustainability: Exploring options for environmentally friendly flower bulb production systems. Eur. J. Plant Pathol. 1997, 103, 217–234. [Google Scholar] [CrossRef]
- Vyvyan, J.R. Allelochemicals as leads for new herbicides and agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a natural products approach to herbicide discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef]
- Iqbal, Z.; Nasir, H.; Hiradate, S.; Fujii, Y. Plant growth inhibitory activity of Lycoris radiata Herb. and the possible involvement of lycorine as an allelochemical. Weed Biol. Manag. 2006, 6, 221–227. [Google Scholar] [CrossRef]
- Macias, F.A.; Castellano, D.; Molinillo, J.M.G. Search for a standard phytotoxic bioassay for allelochemicals. Selection of standard target species. J. Agric. Food Chem. 2000, 48, 2512–2521. [Google Scholar] [CrossRef]
- Petroski, R.J.; Stanley, D.W. Natural compounds for pest and weed control. J. Agric. Food Chem. 2009, 57, 8171–8179. [Google Scholar] [CrossRef]
- Kwant, K.; Engelen, E.; Moerkerken, A.; Keulen, B.V.; Livius, F. Stand van zaken Biobased Economy vanuit het perspectief van Agentschap NL; Dutch Ministry of Economic Affair: The Hague, The Netherlands, 2011. [Google Scholar]
- Ceriotti, G. Narciclasine: An antimitotic substance from Narcissus bulbs. Nature 1967, 213, 595–596. [Google Scholar] [CrossRef]
- Piozzi, F.; Marino, M.; Fuganti, C.; Di Martino, A. Occurrence of non-basic metabolites in Amaryllidaceae. Phytochemistry 1969, 8, 1745–1748. [Google Scholar] [CrossRef]
- Lubbe, A.; Gude, H.; Verpoorte, R.; Choi, Y. Seasonal accumulation of major alkaloids in organs of pharmaceutical crop Narcissus Carlton. Phytochemistry 2013, 88, 43–53. [Google Scholar] [CrossRef]
- Pettit, G.R.; Pettit, G.R., III; Groszek, G.; Backhaus, R.A.; Doubek, D.L.; Barr, R.J.; Meerow, A.W. An investigation of the Amaryllidaceae genus Hymenocallis. J. Nat. Prod. 1995, 58, 756–759. [Google Scholar] [CrossRef]
- Ingrassia, L.; Lefranc, F.; Mathieu, V.; Darro, F.; Kiss, R. Amaryllidaceae isocarbostyril alkaloids and their derivatives as promising antitumor agents. Transl. Oncol. 2008, 1, 1–13. [Google Scholar] [CrossRef]
- Bi, Y.R.; Yung, K.H.; Wong, Y.S. Physiological effects of narciclasine from the mucilage of Narcissus tazetta L. bulbs. Plant Sci. 1998, 135, 103–108. [Google Scholar] [CrossRef]
- Bi, Y.; Zhang, L.; Guo, J.; Yung, K.; Wong, Y. Narciclasine alters chloroplast membrane structure and inhibits 5-aminolevulinic acid and chlorophyll binding protein accumulation in wheat (Triticum aestivum) leaves. New Zealand J. Crop Hort. Sci. 2003, 31, 335–343. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, L.; Na, X.; You, J.; Hu, W.; Liang, X.; Liu, J.; Mao, L.; Wang, X.; Wang, H. Narciclasine inhibits the responses of Arabidopsis roots to auxin. Planta 2012, 236, 1–16. [Google Scholar] [CrossRef]
- Kornienko, A.; Evidente, A. Chemistry, biology and medicinal potential of narciclasine and its congeners. Chem. Rev. 2008, 108, 1982–2014. [Google Scholar] [CrossRef]
- Ingrassia, L.; Lefranc, F.; Dewelle, J.; Pottier, L.; Mathieu, V.; Spiegl-Kreinecker, S.; Sauvage, S.; El Yazidi, M.; Dehoux, M.; Berger, W. Structure-Activity Relationship Analysis of Novel Derivatives of Narciclasine (an Amaryllidaceae Isocarbostyril Derivative) as Potential Anticancer Agents. J. Med. Chem. 2009, 52, 1100–1114. [Google Scholar] [CrossRef]
- Cook, J.; Loudon, J. Chapter XI Alkaloids of the Amaryllidaceae. Alkaloids Chem. Physiol. 1952, 2, 331–352. [Google Scholar] [CrossRef]
- Silva, A.; de Andrade, J.; Machado, K.; Rocha, A.; Apel, M.; Sobral, M.; Henriques, A.; Zuanazzi, J. Screening for cytotoxic activity of extracts and isolated alkaloids from bulbs of Hippeastrum vittatum. Phytomedicine 2008, 15, 882–885. [Google Scholar] [CrossRef]
- Döpke, W.; Gruendemann, E.; Bartoszek, M.; Flatau, S. Alkaloids from Hippeastrum equestre; part 1: Phamine, a new phenanthridone alkaloid. Planta Med. 1995, 61, 564–566. [Google Scholar] [CrossRef]
- Wagner, J.; Pham, H.L.; Döpke, W. Alkaloids from Hippeastrum equestre Herb. 5. Circular dichroism studies. Tetrahedron 1996, 52, 6591–6600. [Google Scholar] [CrossRef]
- Giordani, R.B.; de Andrade, J.P.; Verli, H.; Dutilh, J.H.; Henriques, A.T.; Berkov, S.; Bastida, J.; Zuanazzi, J.A.S. Alkaloids from Hippeastrum morelianum Lem. (Amaryllidaceae). Mag. Res. Chem. 2011, 49, 668–672. [Google Scholar]
- Giordani, R.B.; Pagliosa, L.B.; Henriques, A.T.; Zuanazzi, J.A.S.; Dutilh, J.H.A. Antioxidant and anticolinesterasic effects of Hippeastrum Species (Amaryllidaceae). Quim. Nova 2008, 31, 2042–2046. [Google Scholar] [CrossRef]
- Wildman, W.; Kaufman, C.J. Alkaloids of the Amaryllidaceae. V. Alkaloids of Nerine falcata Barker and N. laticoma (Ker) Dur. and Schinz. 1. J. Am. Chem. Soc. 1955, 77, 4807–4808. [Google Scholar]
- Pettit, G.R.; Gaddamidi, V.; Goswami, A.; Cragg, G.M. Antineoplastic agents, 99. Amaryllis belladonna. J. Nat. Prod. 1984, 47, 796–801. [Google Scholar] [CrossRef]
- Arrigoni, O.; Liso, R.A.; Calabrese, G. Lycorine as an inhibitor of ascorbic acid biosynthesis. Nature 1975, 256, 513–514. [Google Scholar] [CrossRef]
- De Leo, P.; Dalessandro, G.; de Santis, A.; Arrigoni, O. Inhibitory effect of lycorine on cell division and cell elongation. Plant Cell Phys. 1973, 14, 481–486. [Google Scholar]
- Fujii, Y. Allelopathy of Lycoris radiata: Identification of lycorine and crinine as allelochemicals. J. Weed Sci. Technol. 1999, 44, 182–183. [Google Scholar]
- Ptak, A.; El Tahchy, A.; Dupire, F.; Boisbrun, M.; Henry, M.; Chapleur, Y.; Mosi, M.; Laurain-Mattar, D. LCMS and GCMS for the screening of alkaloids in natural and in vitro extracts of Leucojum aestivum. J. Nat. Prod. 2008, 72, 142–147. [Google Scholar]
- Sample Availability: Contact the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wahyuni, D.S.C.; Van der Kooy, F.; Klinkhamer, P.G.L.; Verpoorte, R.; Leiss, K. The Use of Bio-Guided Fractionation to Explore the Use of Leftover Biomass in Dutch Flower Bulb Production as Allelochemicals against Weeds. Molecules 2013, 18, 4510-4525. https://doi.org/10.3390/molecules18044510
Wahyuni DSC, Van der Kooy F, Klinkhamer PGL, Verpoorte R, Leiss K. The Use of Bio-Guided Fractionation to Explore the Use of Leftover Biomass in Dutch Flower Bulb Production as Allelochemicals against Weeds. Molecules. 2013; 18(4):4510-4525. https://doi.org/10.3390/molecules18044510
Chicago/Turabian StyleWahyuni, Dinar S. C., Frank Van der Kooy, Peter G. L. Klinkhamer, Rob Verpoorte, and Kirsten Leiss. 2013. "The Use of Bio-Guided Fractionation to Explore the Use of Leftover Biomass in Dutch Flower Bulb Production as Allelochemicals against Weeds" Molecules 18, no. 4: 4510-4525. https://doi.org/10.3390/molecules18044510
APA StyleWahyuni, D. S. C., Van der Kooy, F., Klinkhamer, P. G. L., Verpoorte, R., & Leiss, K. (2013). The Use of Bio-Guided Fractionation to Explore the Use of Leftover Biomass in Dutch Flower Bulb Production as Allelochemicals against Weeds. Molecules, 18(4), 4510-4525. https://doi.org/10.3390/molecules18044510