Dried Root of Rehmannia glutinosa Prevents Bone Loss in Ovariectomized Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
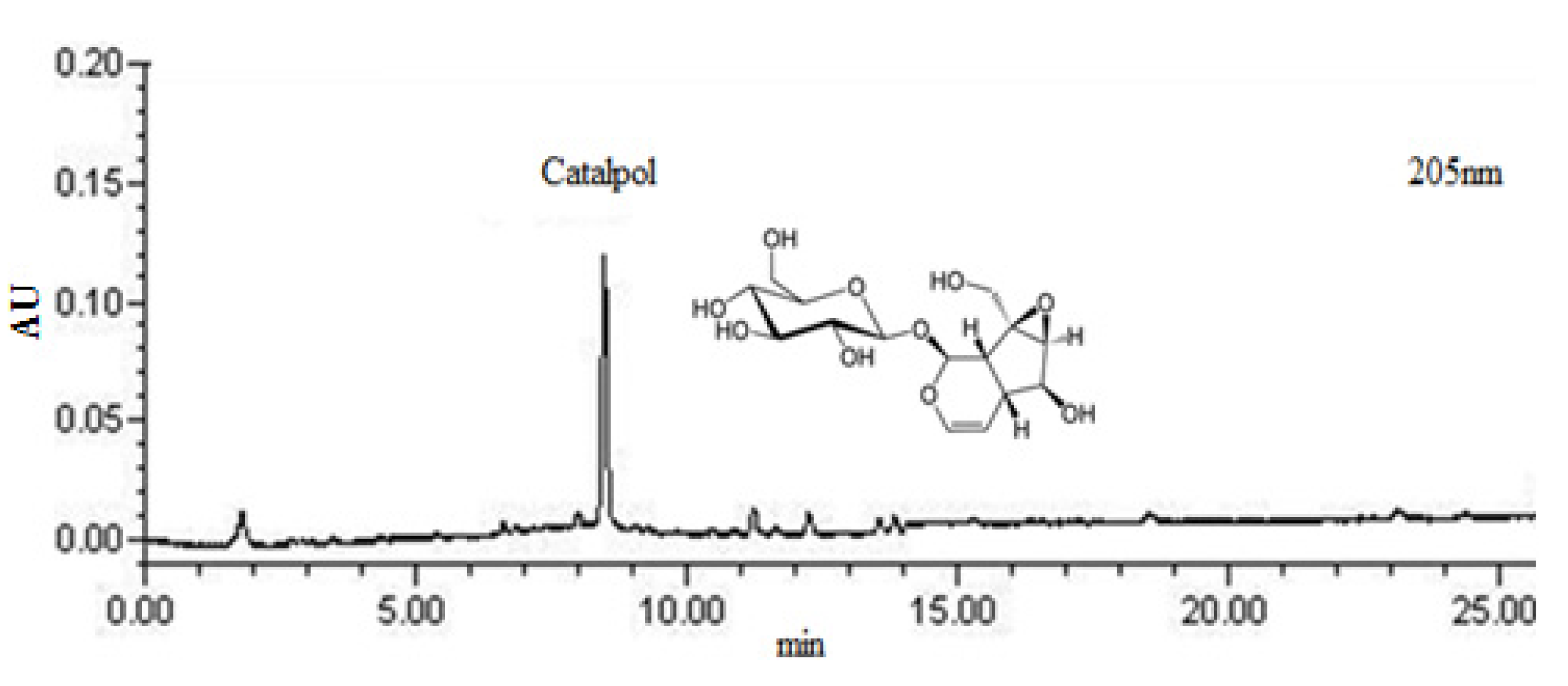
2.1. HPLC Chromatograms for Standardization of DRGE

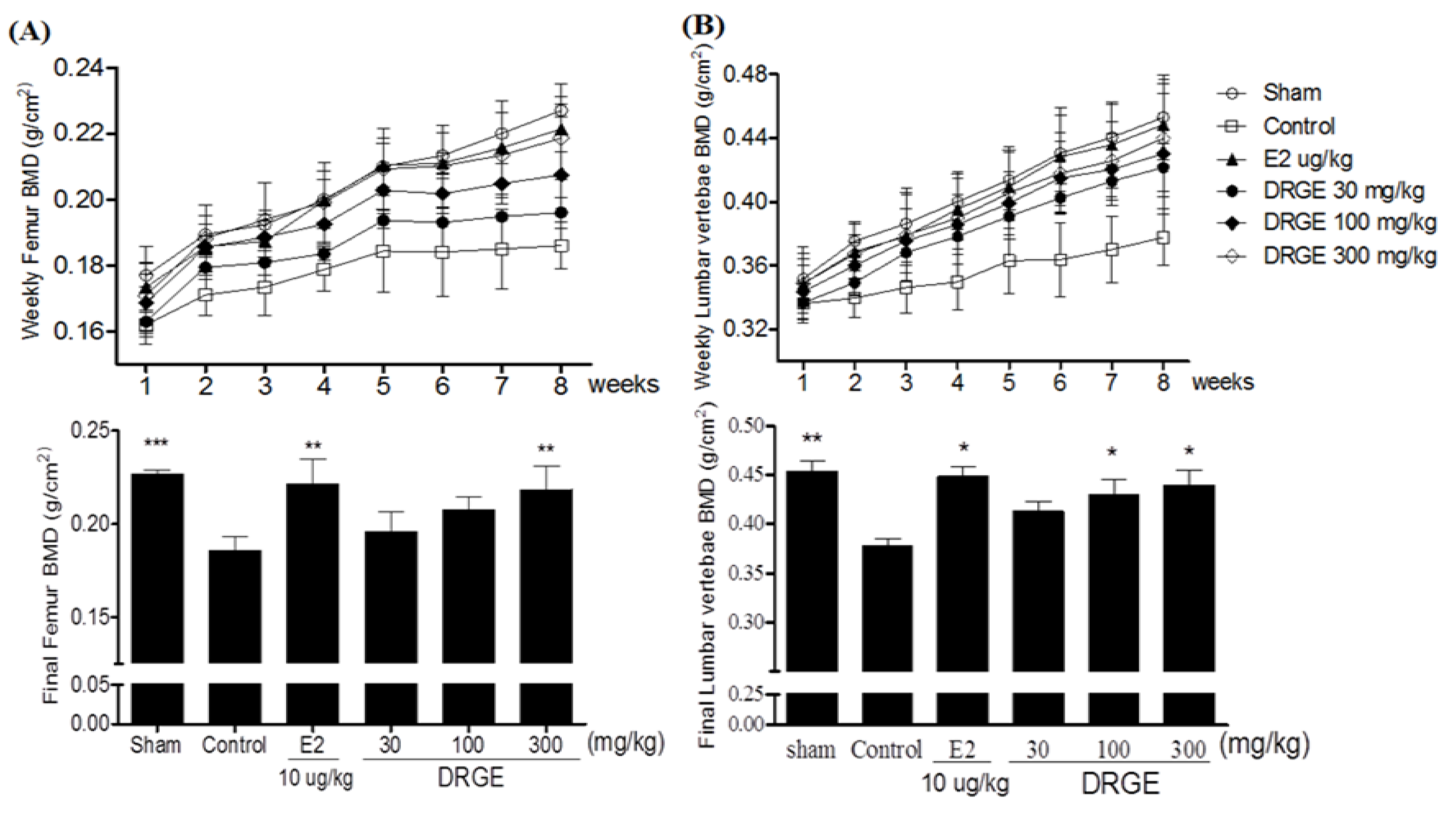
2.2. Bone Mineral Density of the Femur and Lumbar Vertebrae in Treatments of DRGE
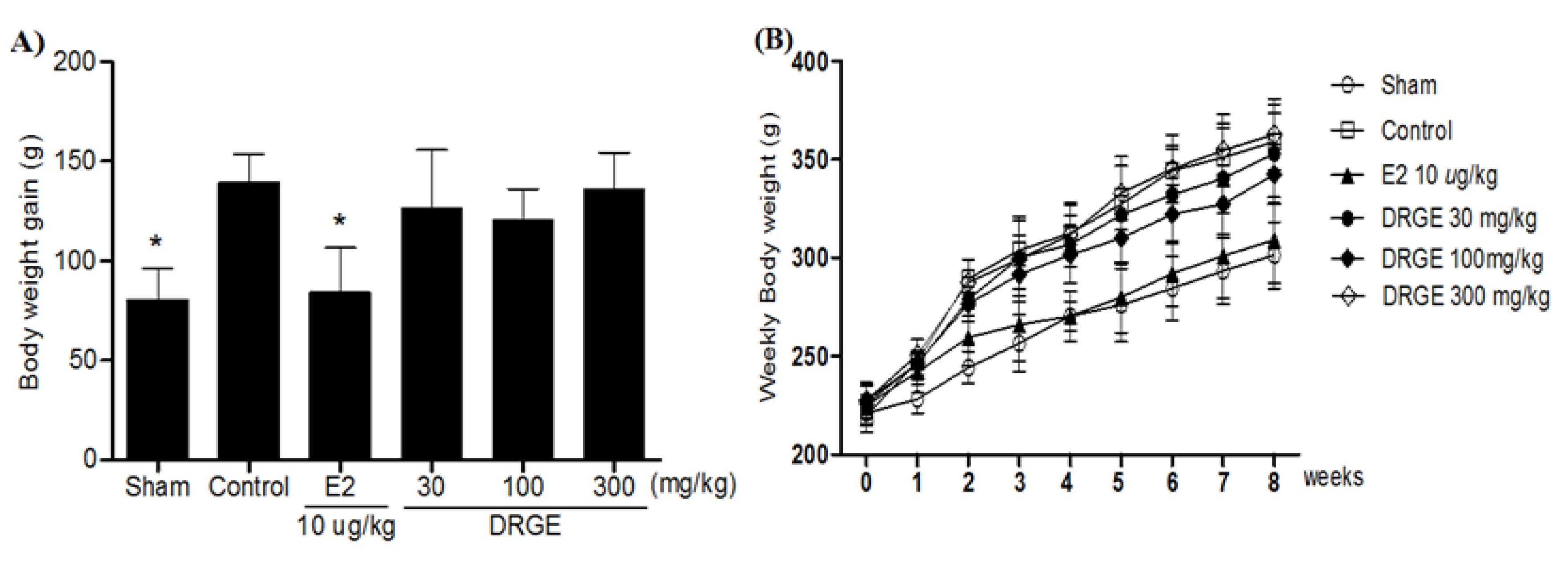
2.3. Weekly Body Weight in DRGE Treatments


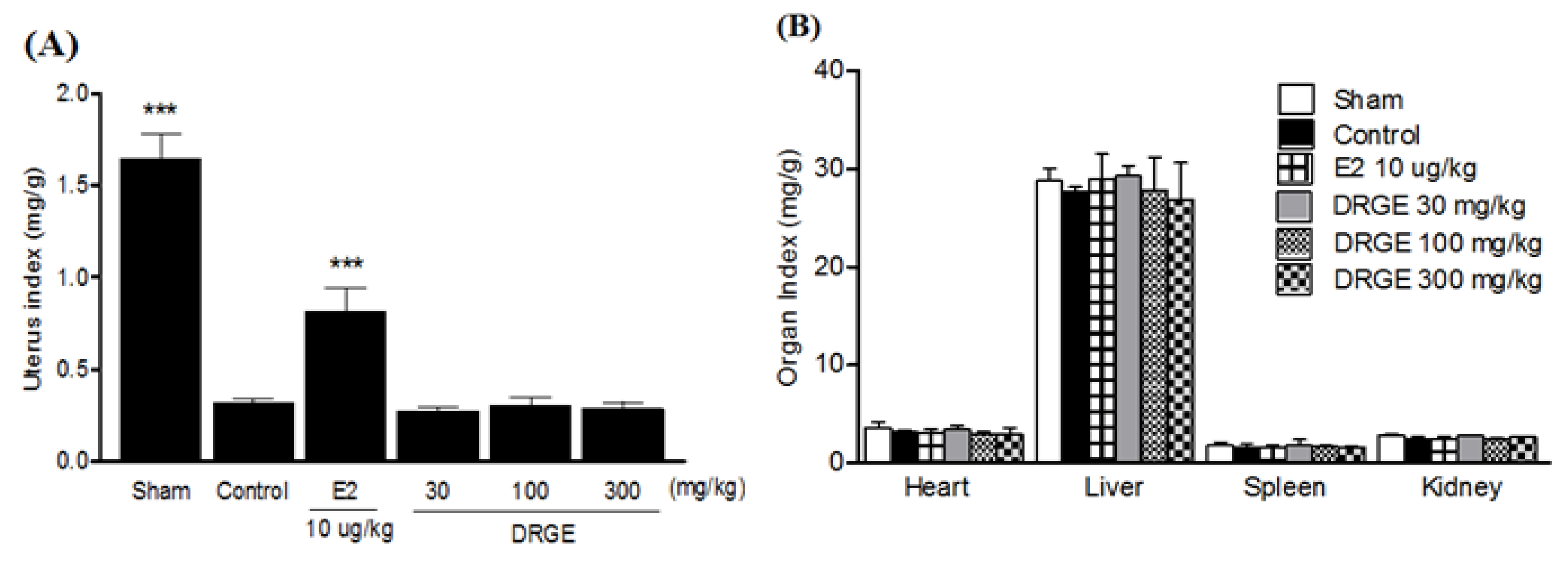
2.4. Uterus and Organ Index in Treatments of DRGE

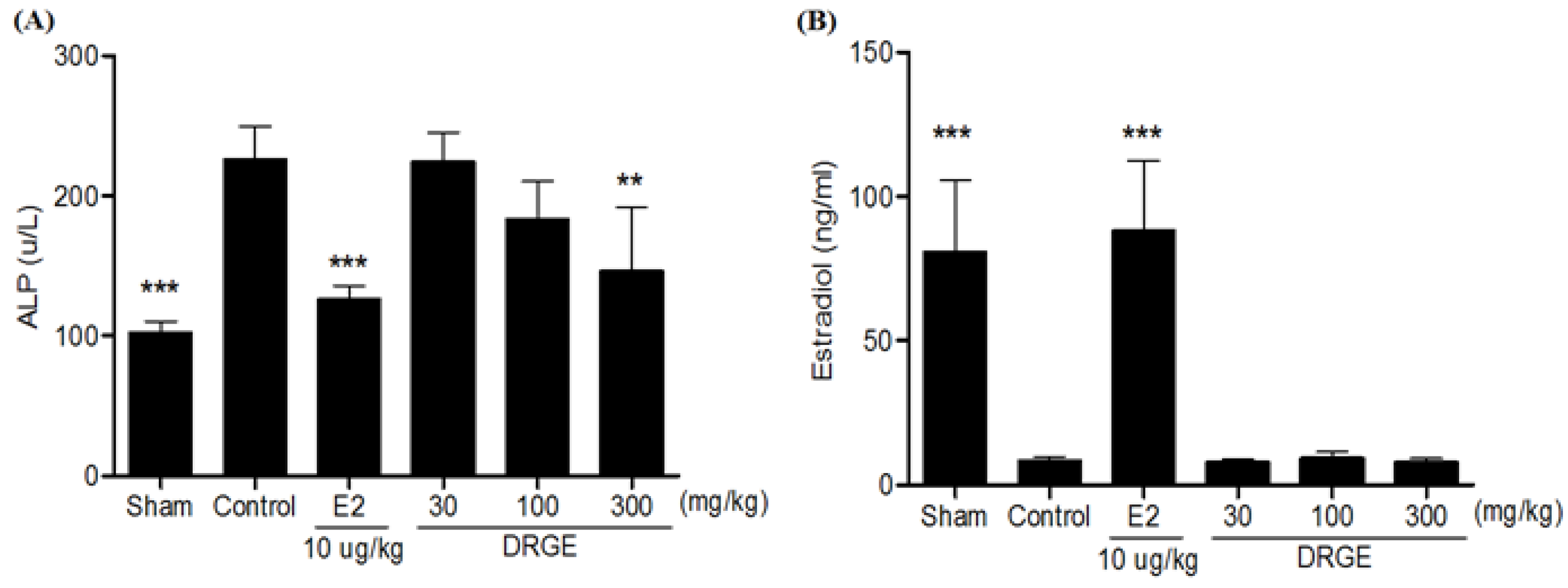
2.5. Serum ALP and Estradiol Concentration in Treatments of DRGE

2.6. Discussion
3. Experimental
3.1. Sample Preparation and HPLC Analysis
3.2. Animals and Treatments
3.3. Bone Mineral Density Measurements
3.4. Serum ALP and Estradiol Analysis
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Invest. 2005, 115, 3318–3325. [Google Scholar] [CrossRef]
- Hansen, M.A.; Overgaard, K.; Riis, B.J.; Christiansen, C. Potential risk factors for development of postmenopausal osteoporosis--examined over a 12-year period. Osteoporos. Int. 1991, 1, 95–102. [Google Scholar]
- Riggs, B.L.; Khosla, S.; Melton, L.J., 3rd. A unitary model for involutional osteoporosis: Estrogen deficiency causes both type I and type II osteoporosis in postmenopausal women and contributes to bone loss in aging men. J. Bone Miner. Res. 1998, 13, 763–773, 3rd. [Google Scholar] [CrossRef]
- Nelson, H.D.; Humphrey, L.L.; Nygren, P.; Teutsch, S.M.; Allan, J.D. Postmenopausal hormone replacement therapy: Scientific review. JAMA 2002, 288, 872–881. [Google Scholar] [CrossRef]
- Pradhan, A.D.; Manson, J.E.; Rossouw, J.E.; Siscovick, D.S.; Mouton, C.P.; Rifai, N.; Wallace, R.B.; Jackson, R.D.; Pettinger, M.B.; Ridker, P.M. Inflammatory biomarkers, hormone replacement therapy, and incident coronary heart disease: Prospective analysis from the Women’s Health Initiative observational study. JAMA 2002, 288, 980–987. [Google Scholar] [CrossRef]
- Chang, W.T.; Choi, Y.H.; van der Heijden, R.; Lee, M.S.; Lin, M.K.; Kong, H.; Kim, H.K.; Verpoorte, R.; Hankemeier, T.; van der Greef, J.; et al. Traditional processing strongly affects metabolite composition by hydrolysis in Rehmannia glutinosa roots. Chem. Pharm. Bull. 2011, 59, 546–552. [Google Scholar] [CrossRef]
- Zhang, R.X.; Li, M.X.; Jia, Z.P. Rehmannia glutinosa: Review of botany, chemistry and pharmacology. J. Ethnopharmacol. 2008, 117, 199–214. [Google Scholar] [CrossRef]
- Chao, J.C.; Chiang, S.W.; Wang, C.C.; Tsai, Y.H.; Wu, M.S. Hot water-extracted Lycium barbarum and Rehmannia glutinosa inhibit proliferation and induce apoptosis of hepatocellular carcinoma cells. World J. Gastroenterol. 2006, 12, 4478–4484. [Google Scholar]
- Zhang, D.; Wen, X.S.; Wang, X.Y.; Shi, M.; Zhao, Y. Antidepressant effect of Shudihuang on mice exposed to unpredictable chronic mild stress. J. Ethnopharmacol. 2009, 123, 55–60. [Google Scholar] [CrossRef]
- Chang, G.T.; Min, S.Y.; Kim, J.H.; Kim, S.H.; Kim, J.K.; Kim, C.H. Anti-thrombic activity of Korean herbal medicine, Dae-Jo-Whan and its herbs. Vascul. Pharmacol. 2005, 43, 283–288. [Google Scholar] [CrossRef]
- Kiho, T.; Watanabe, T.; Nagai, K.; Ukai, S. Hypoglycemic activity of polysaccharide fraction from rhizome of Rehmannia glutinosa Libosch. f. hueichingensis Hsiao and the effect on carbohydrate metabolism in normal mouse liver. Yakugaku zasshi 1992, 112, 393–400. [Google Scholar]
- Oh, K.O.; Kim, S.W.; Kim, J.Y.; Ko, S.Y.; Kim, H.M.; Baek, J.H.; Ryoo, H.M.; Kim, J.K. Effect of Rehmannia glutinosa Libosch extracts on bone metabolism. Clinica Chimica Acta 2003, 334, 185–195. [Google Scholar] [CrossRef]
- Jin, U.H.; Kim, D.I.; Lee, T.K.; Lee, D.N.; Kim, J.K.; Lee, I.S.; Kim, C.H. Herbal formulation, Yukmi-jihang-tang-Jahage, regulates bone resorption by inhibition of phosphorylation mediated by tyrosine kinase Src and cyclooxygenase expression. J. Ethnopharmacol. 2006, 106, 333–343. [Google Scholar] [CrossRef]
- Liu, H.D.; Lin, F.S.; Li, E.; Wu, M.S.; Tong, X.X. The influence of the different components of nourishing kidney herbs on osteoporosis rats. Zhongguo Zhongyao Zazhi 2003, 28, 262–265. [Google Scholar]
- Chang, W.T.; Thissen, U.; Ehlert, K.A.; Koek, M.M.; Jellema, R.H.; Hankemeier, T.; van der Greef, J.; Wang, M. Effects of growth conditions and processing on Rehmannia glutinosa using fingerprint strategy. Planta medica 2006, 72, 458–467. [Google Scholar] [CrossRef]
- Hoegh-Andersen, P.; Tanko, L.B.; Andersen, T.L.; Lundberg, C.V.; Mo, J.A.; Heegaard, A.M.; Delaisse, J.M.; Christgau, S. Ovariectomized rats as a model of postmenopausal osteoarthritis: Validation and application. Arthritis Res. Ther. 2004, 6, R169–R180. [Google Scholar] [CrossRef] [Green Version]
- Jee, W.S.; Yao, W. Overview: Animal models of osteopenia and osteoporosis. J. Musculoskelet. Neuronal Interact. 2001, 1, 193–207. [Google Scholar]
- Lelovas, P.P.; Xanthos, T.T.; Thoma, S.E.; Lyritis, G.P.; Dontas, I.A. The laboratory rat as an animal model for osteoporosis research. Comp. Med. 2008, 58, 424–430. [Google Scholar]
- Nishizawa, Y.; Nakamura, T.; Ohta, H.; Kushida, K.; Gorai, I.; Shiraki, M.; Fukunaga, M.; Hosoi, T.; Miki, T.; Chaki, O.; et al. Guidelines for the use of biochemical markers of bone turnover in osteoporosis (2004). J. Bone Miner. Metab. 2005, 23, 97–104. [Google Scholar]
- Yogesh, H.S.; Chandrashekhar, V.M.; Katti, H.R.; Ganapaty, S.; Raghavendra, H.L.; Gowda, G.K.; Goplakhrishna, B. Anti-osteoporotic activity of aqueous-methanol extract of Berberis aristata in ovariectomized rats. J. Ethnopharmacol. 2011, 134, 334–338. [Google Scholar] [CrossRef]
- Devareddy, L.; Khalil, D.A.; Smith, B.J.; Lucas, E.A.; Soung, D.Y.; Marlow, D.D.; Arjmandi, B.H. Soy moderately improves microstructural properties without affecting bone mass in an ovariectomized rat model of osteoporosis. Bone 2006, 38, 686–693. [Google Scholar] [CrossRef]
- Dang, Z.C.; Van Bezooijen, R.L.; Karperien, M.; Papapoulos, S.E.; Lowik, C.W.G.M. Exposure of KS483 cells to estrogen enhances osteogenesis and inhibits adipogenesis. J. Bone Miner. Res. 2002, 17, 394–405. [Google Scholar] [CrossRef]
- Heine, P.A.; Taylor, J.A.; Iwamoto, G.A.; Lubahn, D.B.; Cooke, P.S. Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc. Natl. Acad. Sci. USA 2000, 97, 12729–12734. [Google Scholar]
- Joyner, J.M.; Hutley, L.J.; Cameron, D.P. Estrogen receptors in human preadipocytes. Endocrine 2001, 15, 225–230. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Korach, K.S. Oestrogen receptor knockout mice: Roles for oestrogen receptors alpha and beta in reproductive tissues. Reproduction 2003, 125, 143–149. [Google Scholar] [CrossRef]
- Raju, B.L.; Lin, S.J.; Hou, W.C.; Lai, Z.Y.; Liu, P.C.; Hsu, F.L. Antioxidant iridoid glucosides from Wendlandia formosana. Nat. Prod. Res. 2004, 18, 357–364. [Google Scholar] [CrossRef]
- Mody, N.; Parhami, F.; Sarafian, T.A.; Demer, L.L. Oxidative stress modulates osteoblastic differentiation of vascular and bone cells. Free Radical. Bio. Med. 2001, 31, 509–519. [Google Scholar] [CrossRef]
- Kim, H.M.; An, C.S.; Jung, K.Y.; Choo, Y.K.; Park, J.K.; Nam, S.Y. Rehmannia glutinosa inhibits tumour necrosis factor-alpha and interleukin-1 secretion from mouse astrocytes. Pharmacol. Res. 1999, 40, 171–176. [Google Scholar] [CrossRef]
- Evans, D.B.; Bunning, R.A.; Russell, R.G. The effects of recombinant human interleukin-1 beta on cellular proliferation and the production of prostaglandin E2, plasminogen activator, osteocalcin and alkaline phosphatase by osteoblast-like cells derived from human bone. Biochem. biophysical Res. Commun. 1990, 166, 208–216. [Google Scholar] [CrossRef]
- Stashenko, P.; Dewhirst, F.E.; Peros, W.J.; Kent, R.L.; Ago, J.M. Synergistic interactions between interleukin 1, tumor necrosis factor, and lymphotoxin in bone resorption. J. Immunol. 1987, 138, 1464–1468. [Google Scholar]
- Sample Availability: Samples of the DRGE is available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lim, D.W.; Kim, Y.T. Dried Root of Rehmannia glutinosa Prevents Bone Loss in Ovariectomized Rats. Molecules 2013, 18, 5804-5813. https://doi.org/10.3390/molecules18055804
Lim DW, Kim YT. Dried Root of Rehmannia glutinosa Prevents Bone Loss in Ovariectomized Rats. Molecules. 2013; 18(5):5804-5813. https://doi.org/10.3390/molecules18055804
Chicago/Turabian StyleLim, Dong Wook, and Yun Tai Kim. 2013. "Dried Root of Rehmannia glutinosa Prevents Bone Loss in Ovariectomized Rats" Molecules 18, no. 5: 5804-5813. https://doi.org/10.3390/molecules18055804
APA StyleLim, D. W., & Kim, Y. T. (2013). Dried Root of Rehmannia glutinosa Prevents Bone Loss in Ovariectomized Rats. Molecules, 18(5), 5804-5813. https://doi.org/10.3390/molecules18055804