Acetylcholinesterase Inhibition and in Vitro and in Vivo Antioxidant Activities of Ganoderma lucidum Grown on Germinated Brown Rice


Abstract
:1. Introduction
2. Results and Discussion
2.1. Total Phenolic, Flavonoid, Ascorbic Acid, β-Carotene, Lycopene Content & Yield of GLBR Extract

{kind=link}
{kind=link}
| Phenolic compound in GLBR | Composition (mg/mL) |
|---|---|
| Quercetin | 13.53 |
| Ursolic acid | 12.87 |
| Keampferol | 8.62 |
| Terpeniol | 7.35 |
| Coumarin | 5.65 |
| Catechin | 4.09 |
| Myricetin | 3.85 |
| Ferulic acid | 2.13 |
| Thymol | 0.67 |
| Rutin | 0.65 |
| Lupane | 0.29 |
| Cyanidin | 0.04 |
| Elagic acid | 0.001 |
| Caffeic acid | nd |
| Epicatechin | nd |
2.2. HPLC Analysis of GLBR Extract
2.3. Antioxidant Activity in Vitro
2.3.1. DPPH Radical Scavenging Activity of GLBR Extract
| Extract | DPPH radical scavenging activity | ABTS radical scavenging activity | Hydroxyl radical scavenging activity | Superoxide radical scavenging activity | Metal chelating activity | Lipid peroxidation | Cellular antioxidant activity |
| ICa50 value mg/mL | ICa50 value mg/mL | ICa50 value mg/mL | ICa50 value mg/mL | ICa50 value mg/mL | ICa50 value mg/mL | ICa50 value mg/mL | |
| GLBR | 0.64 ± 0.02 a | 0.44 ± 0.02 a | 0.164 ± 0.017 a | 0.62 ± 0.05 a | 0.84 ± 0.01 a | 1.48 ± 0.13 a | 1.46 ± 0.17 a |
| Positive control | 0.19 ± 0.002 b (Ascorbic Acid) | 0.02 ± 0.0002 b (Ascorbic Acid) | 0.005 ± 0.0003 b (Ascorbic Acid) | 0.053 ± 0.002 b (Ascorbic Acid) | 0.009 ± 0.0002 b (EDTA) | 0.59 ± 0.06 b (Ascorbic Acid) | 0.11 ± 0.02 b (Ascorbic Acid) |
2.3.2. ABTS Radical Scavenging Activity of GLBR Extract
2.3.3. Hydroxyl Radical Scavenging Activity
2.3.4. Superoxide Radical Scavenging Activity
2.3.5. Effect of GLBR Extract on Metal Chelating Activity
2.3.6. Effect of GLBR Extract on Lipid Peroxidation
2.3.7. Effect of GLBR Extract on Cellular Antioxidant Activity (CAA)
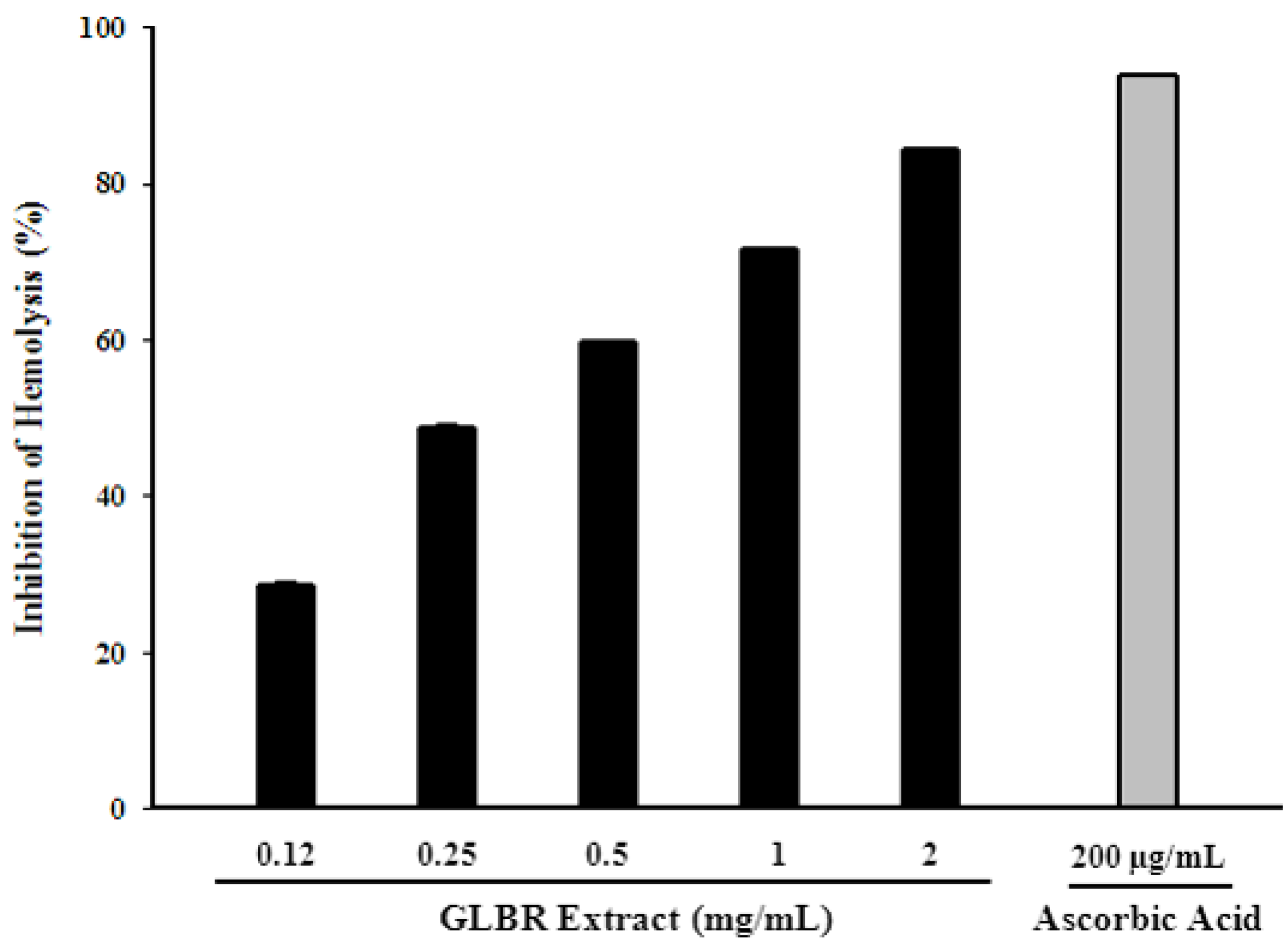
2.3.8. Effect of GLBR Extract on Inhibition of Erythrocyte Haemolysis

2.4. Antioxidant Activities in Vivo
| Group | Serum | Liver | Brain | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SOD (U/mL) | GPx (U/mL) | CAT (U/mL) | SOD (U/mL) | GPx (U/mL) | CAT (U/mL) | SOD (U/mL) | GPx (U/mL) | CAT (U/mL) | |
| Control | 43.27 ± 5.03 a | 48.98 ± 4.67 a | 13.2 ± 0.70 a | 35.3 ± 2.04 a | 52.5 ± 4.30 a | 14.4 ± 0.65 a | 98.41 ± 5.55 a | 262.67 ± 8.12 a | 5.67 ± 0.99 a |
| Positive control | 61.53 ± 2.93 b | 51.69 ± 2.52 a | 14.7 ± 0.77 a | 41.7 ± 2.49 a | 61.7 ± 3.90 b | 16.3 ± 0.79 b | 110.45 ± 4.65 b | 276.07 ± 11.05 a | 7.27 ± 1.03 a |
| Extract (100 mg/Kg) | 44.25 ± 2.23 a | 49.79 ± 3.03 a | 14.1 ± 0.95 a | 31.7 ± 3.11 a | 65.1 ± 4.30 c | 16.9 ± 0.80 c | 123.41 ± 4.39 c | 275.94 ± 6.91 a | 7.82 ± 1.06 b |
| Extract (200 mg/Kg) | 57.56 ± 5.15 c | 53.08 ± 2.44 a | 15.7 ± 0.61 b | 52.8 ± 4.37 b | 70.5 ± 2.57 d | 18.1 ± 0.94 d | 127.64 ± 7.16 d | 294.35 ± 8.05 b | 9.93 ± 1.04 c |
| Extract (400 mg/Kg) | 71.72 ± 8.18 d | 60.05 ± 2.36 b | 17.5 ± 0.74 c | 67.3 ± 4.20 c | 79.4 ± 3.84 e | 19.9 ± 0.46 e | 138.38 ± 7.11 e | 305.55 ± 6.66 c | 11.29 ± 0.98 d |
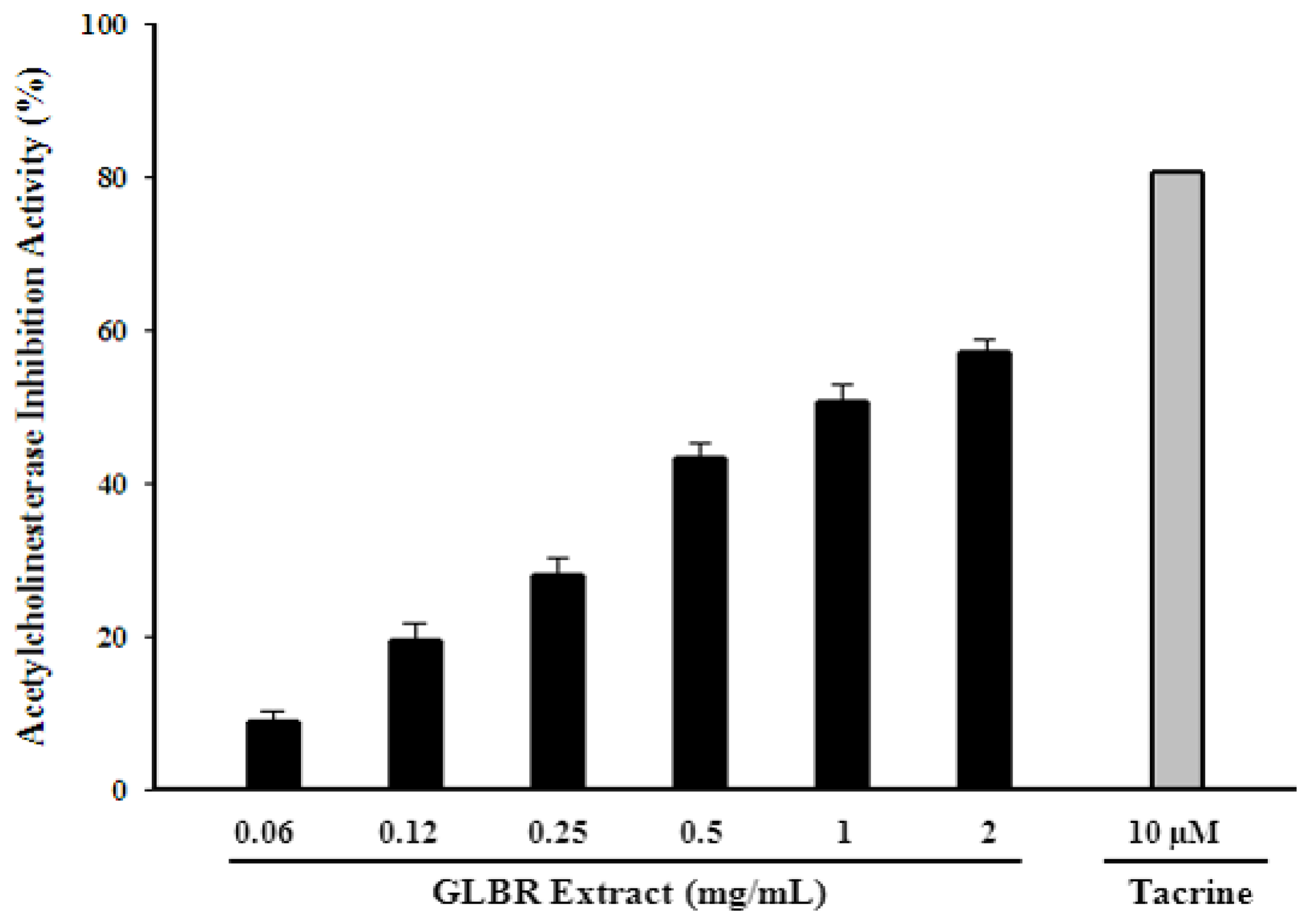
2.5. Acetylcholinesterase (AChE) Inhibitory Activity

3. Experimental
3.1. Chemicals and Materials
3.2. Preparation of Ganoderma lucidum Grown on Germinated Brown Rice (GLBR) Extract
3.3. Measurement of Total Phenolic, Flavonoid, Ascorbic Acid, β-carotene, Lycopene Content
3.4. High Performance Liquid Chromatography (HPLC) Profiling of the Extract
3.5. Assay of Antioxidant Activity in Vitro
3.5.1. DPPH Radical-Scavenging Activity
3.5.2. ABTS Radical-Scavenging Activity
3.5.3. Hydroxyl Radical Scavenging Activity
3.5.4. Superoxide Radical Scavenging Activity
3.5.5. Metal Chelating Activity
3.5.6. Inhibition of Lipid Peroxidation Using Thiobarbituric Acid Reactive Substances (TBARS)
3.5.7. Cell Culture
3.5.8. Cellular Antioxidant Activity (CAA)
3.5.9. Anti-Hemolysis Activity
3.6. Assay of Antioxidant Activity in Vivo
3.6.1. Animals and Experimental Design
3.6.2. Biochemical Assay
3.7. In-Vitro Acetylcholinesterase Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Styskal, J.L.; Remmen, H.V.; Richardson, A.; Salmon, A.B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free Radic. Biol. Med. 2012, 52, 46–58. [Google Scholar] [CrossRef]
- Sun, J.; He, H.; Xie, B.J. Novel antioxidant peptides from fermented mushroom Ganoderma lucidum. J. Agric. Food Chem. 2004, 52, 6646–6652. [Google Scholar] [CrossRef]
- Zhou, D.Y.; Zhu, B.W.; Qiao, L.; Wu, H.T.; Li, D.M.; Yang, J.F.; Murata, Y. In vitro antioxidant activity of enzymatic hydrolysates prepared from abalone (Haliotis discus hannai Ino) viscera. Food Bioprod. Process. 2012, 90, 148–154. [Google Scholar]
- Heleno, S.A.; Barros, L.; Martins, A.; Queiroz, M.J.R.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Fruiting body, spores and in vitro produced mycelium of Ganoderma lucidum from Northeast Portugal: A comparative study of the antioxidant potential of phenolic and polysaccharidic extracts. Food Res. Int. 2012, 46, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Lahiri, D.K.; Farlow, M.R.; Greig, N.H.; Sambamurti, K. Current drug targets for Alzheime’s disease treatment. Drug Dev. Res. 2002, 56, 267–281. [Google Scholar] [CrossRef]
- Bastianetto, S.; Quirion, R. Natural extracts as possible protective agents of brain aging. Neurobiol. Ag. 2002, 23, 891–897. [Google Scholar] [CrossRef]
- Duthie, G.G.; Duthie, S.J.; Kyle, J.A.M. Plant polyphenols in cancer and heart disease: Implications as nutritional antioxidants. Nutr. Res. Rev. 2000, 13, 79–106. [Google Scholar] [CrossRef]
- Ferreira, I.C.F.R.; Vaz, J.A.; Vasconcelos, M.H.; Martins, A. Compounds from wild mushrooms with antitumor potential. Anti-cancer Agents Med. Chem. 2010, 10, 424–436. [Google Scholar] [CrossRef]
- Yan, K.; Zhang, W.; Yu, H.; Wang, H.; Zhang, X. New polysaccharide compounds derived from submerged culture of Ganoderma lucidum and Lycium barbarum. Food Technol. Biotechnol. 2010, 48, 94–101. [Google Scholar]
- Oh, S.H.; Choi, W.G. Production of the quality germinated brown rices containing high c-aminobutyric acid by chitosan application. Kor. J. Biotechnol. Bioeng. 2000, 15, 615–620. [Google Scholar]
- Bau, H.M.; Villaume, C.H.; Mejean, L. Effects of soybean (Glycine max) germination on biologically active components, nutritional values of seeds, and biological characteristics in rat. Nahrung 2000, 44, 2–6. [Google Scholar] [CrossRef]
- Paucar-Menacho, L.M.; Berhow, M.A.; Gontijo Mandarino, J.M.; Chang, Y.K.; Gonzalez De Mejia, E. Effect of time and temperature on bioactive compounds in germinated Brazilian soybean cultivar BRS 258. Food Res. Int. 2010, 43, 1856–1865. [Google Scholar] [CrossRef]
- Zbarsky, V.; Datla, K.P.; Parkar, S.; Rai, D.K.; Aruoma, O.I.; Dexter, D.T. Neuroprotective properties of the natural phenolic antioxidants curcumin and naringenin but not quercetin and fisetin in a 6-OHDA model of Parkinson’s disease. Free Radc. Res. 2005, 39, 1119–25. [Google Scholar] [CrossRef]
- Li, L.; Zhang, X.; Cui, L.; Wang, L.; Liu, H.; Ji, H; Du, Y. Ursolic acid promotes the neuroprotection by activating Nrf2 pathway after cerebral ischemia in mice. Brain Res. 2013, 1497, 32–39. [Google Scholar] [CrossRef]
- Mahendran, S.; Anandapandian, K.T.K.; Shankar, T.; Chellaram, C.; Vijayabaskar, P. Antioxidant properties of Ganoderma Lucidum crude Exopolysaccharide. Indian J. Innov. Dev. 2012, 1, 1–6. [Google Scholar]
- Wickens, A.P. Aging and the free radical theory. Respir. Physiol. Neurobiol. 2001, 128, 379–391. [Google Scholar] [CrossRef]
- Jayakumar, T.; Thomas, P.A; Geraldine, P. In-vitro antioxidant activities of an ethanolic extract of the oyster mushroom, Pleurotus ostreatu. Innov. Food Sci. Emerg. Technol. 2009, 10, 228–234. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Novel antioxidants in food quality preservation and health promotion. Eur. J. Lipid Sci. Technol. 2010, 112, 930–940. [Google Scholar] [CrossRef]
- Lee, J.M.; Kwon, H.; Jeong, H.; Lee, J.W.; Lee, S.Y.; Baek, S.J.; Surh, Y.J. Inhibition of lipid peroxidation and oxidative DNA damage by Ganoderma lucidum. Phytother. Res. 2001, 15, 245–249. [Google Scholar] [CrossRef]
- Molina-Holgado, F.; Hider, R.C.; Gaeta, A.; Williams, R.; Francis, P. Metals ions and neurodegeneration. Biometals 2007, 20, 639–654. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Liu, J.; Jia, L.; Kan, J.; Jin, C.H. In vitro and in vivo antioxidant activity of ethanolic extract of white button mushroom (Agaricus bisporus). Food Chem. Toxicol. 2013, 51, 310–316. [Google Scholar] [CrossRef]
- Jeon, T.I.; Hwang, S.G.; Lim, B.O.; Park, D.K. Extracts of Phellinus linteus grown on germinated brown rice suppress liver damage induced by carbon tetrachloride in rats. Biotechnol. Lett. 2003, 25, 2093–2096. [Google Scholar] [CrossRef]
- Sakanaka, S.; Tachibana, Y.; Okada, Y. Preparation and antioxidant properties of extracts of Japanese persimmon tea (kakinoha-cha). Food Chem. 2005, 89, 569–575. [Google Scholar] [CrossRef]
- Barros, L.; Ferreira, M.J.; Queiros, B.; Ferreira, I.C.F.R.; Baptista, P. Total phenols, ascorbic acid, b-carotene and lycopene in Portuguese wild edible mushrooms and their antioxidant activities. Food Chem. 2007, 100, 413–419. [Google Scholar]
- Jeong, C.H.; Choi, G.N.; Kim, J.H.; Kwak, J.H.; Heo, H.J.; Shim, K.H.; Cho, B.R.; Bae, Y.I.; Choi, J.S. In vitro antioxidant activities and phenolic composition of hot water extract from different parts of Cudrania tricuspidata. J. Food Sci. Nutr. 2009, 14, 283–289. [Google Scholar] [CrossRef]
- Rosen, G.M.; Rauckman, E.J. Spin trapping of superoxide and hydroxyl radicals. Methods Enzymol. 1984, 105, 198–209. [Google Scholar] [CrossRef]
- Guo, Q.; Zhao, B.; Packer, L. Electron spin resonance study of free radicals formed from a procyanidin-rich pine (Pinus maritima) bark extract, pycnogenol. Free Radic. Biol. Med. 1999, 27, 1308–1312. [Google Scholar] [CrossRef]
- Dinis, T.C.P.; Madeira, V.M.C.; Almeida, L.M. Action of phenolic derivatives (acetoaminophen, salicylate and 5-aminosalicylate) as inhibitors of membrane lipid peroxidation and as peroxyl radical scavengers. Arch. Biochem. Biophys. 1994, 315, 161–169. [Google Scholar] [CrossRef]
- Pinela, J.; Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Influence of the drying method in the antioxidant potential of four shrubby flowering plants from the tribe Genisteae (Fabaceae). Food Chem. Toxicol. 2011, 49, 2983–2989. [Google Scholar] [CrossRef]
- Zhang, A.; Zhu, Q.Y.; Luk, Y.S.; Ho, K.Y.; Fung, K.P.; Chen, Z.Y. Inhibitory effects of jasmine green tea epicatechin isomers on free radical-induced lysis of red blood cells. Life Sci. 1997, 61, 383–394. [Google Scholar] [CrossRef]
- Rahman, A.U.; Choudhary, M.I. Bioactive natural products as a potential source of new pharmacophores a theory of memory. Pure Appl. Chem. 2001, 73, 555–560. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds quercetin, ursolic acid, keampferol, terpeniol, coumarin, catechin, myricetin, ferulic acid, thymol, rutin, lupine, cyaniding and elagic acid are available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hasnat, M.A.; Pervin, M.; Lim, B.O. Acetylcholinesterase Inhibition and in Vitro and in Vivo Antioxidant Activities of Ganoderma lucidum Grown on Germinated Brown Rice. Molecules 2013, 18, 6663-6678. https://doi.org/10.3390/molecules18066663
Hasnat MA, Pervin M, Lim BO. Acetylcholinesterase Inhibition and in Vitro and in Vivo Antioxidant Activities of Ganoderma lucidum Grown on Germinated Brown Rice. Molecules. 2013; 18(6):6663-6678. https://doi.org/10.3390/molecules18066663
Chicago/Turabian StyleHasnat, Md. Abul, Mehnaz Pervin, and Beong Ou Lim. 2013. "Acetylcholinesterase Inhibition and in Vitro and in Vivo Antioxidant Activities of Ganoderma lucidum Grown on Germinated Brown Rice" Molecules 18, no. 6: 6663-6678. https://doi.org/10.3390/molecules18066663
APA StyleHasnat, M. A., Pervin, M., & Lim, B. O. (2013). Acetylcholinesterase Inhibition and in Vitro and in Vivo Antioxidant Activities of Ganoderma lucidum Grown on Germinated Brown Rice. Molecules, 18(6), 6663-6678. https://doi.org/10.3390/molecules18066663