Isolation and Characterization of a Novel Lectin from the Edible Mushroom Stropharia rugosoannulata


Abstract
:1. Introduction
2. Results and Discussion
2.1. Results



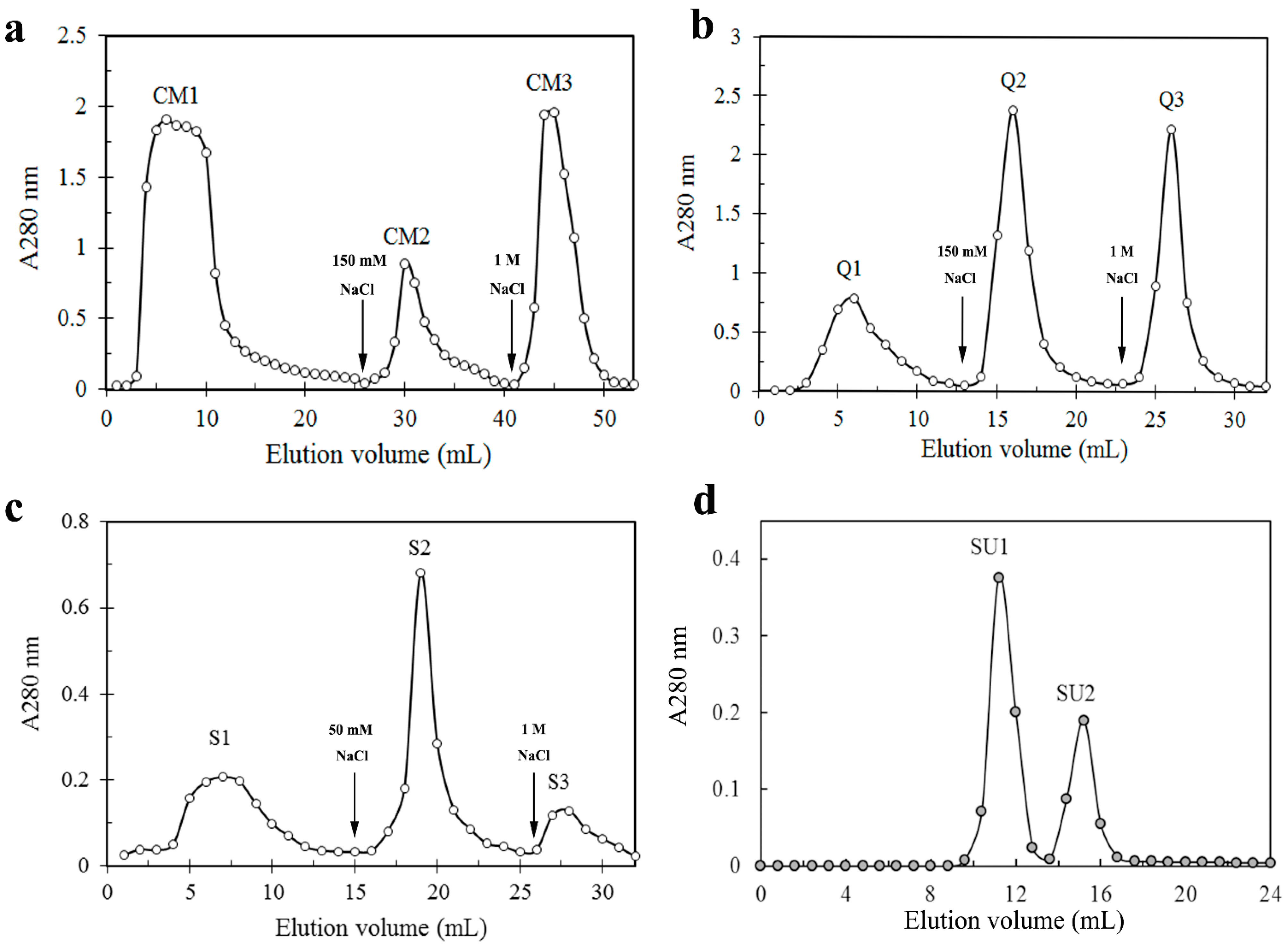{kind=link}
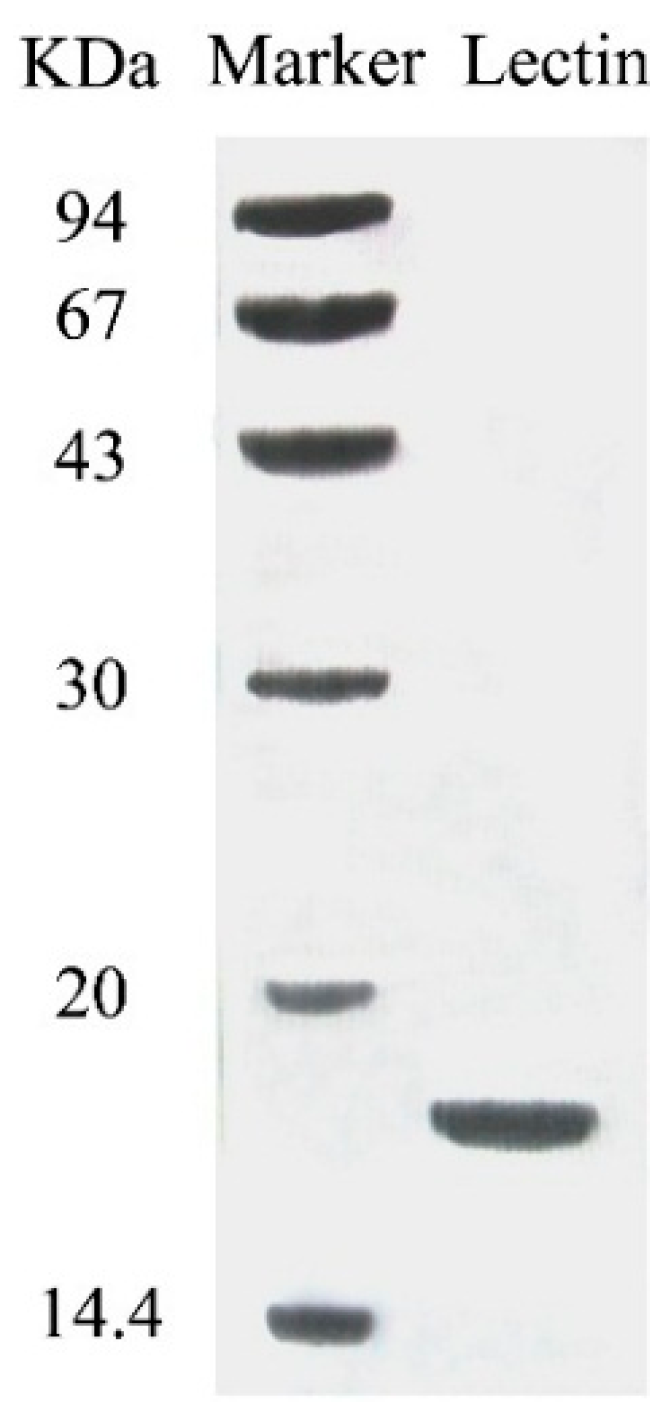{kind=link}
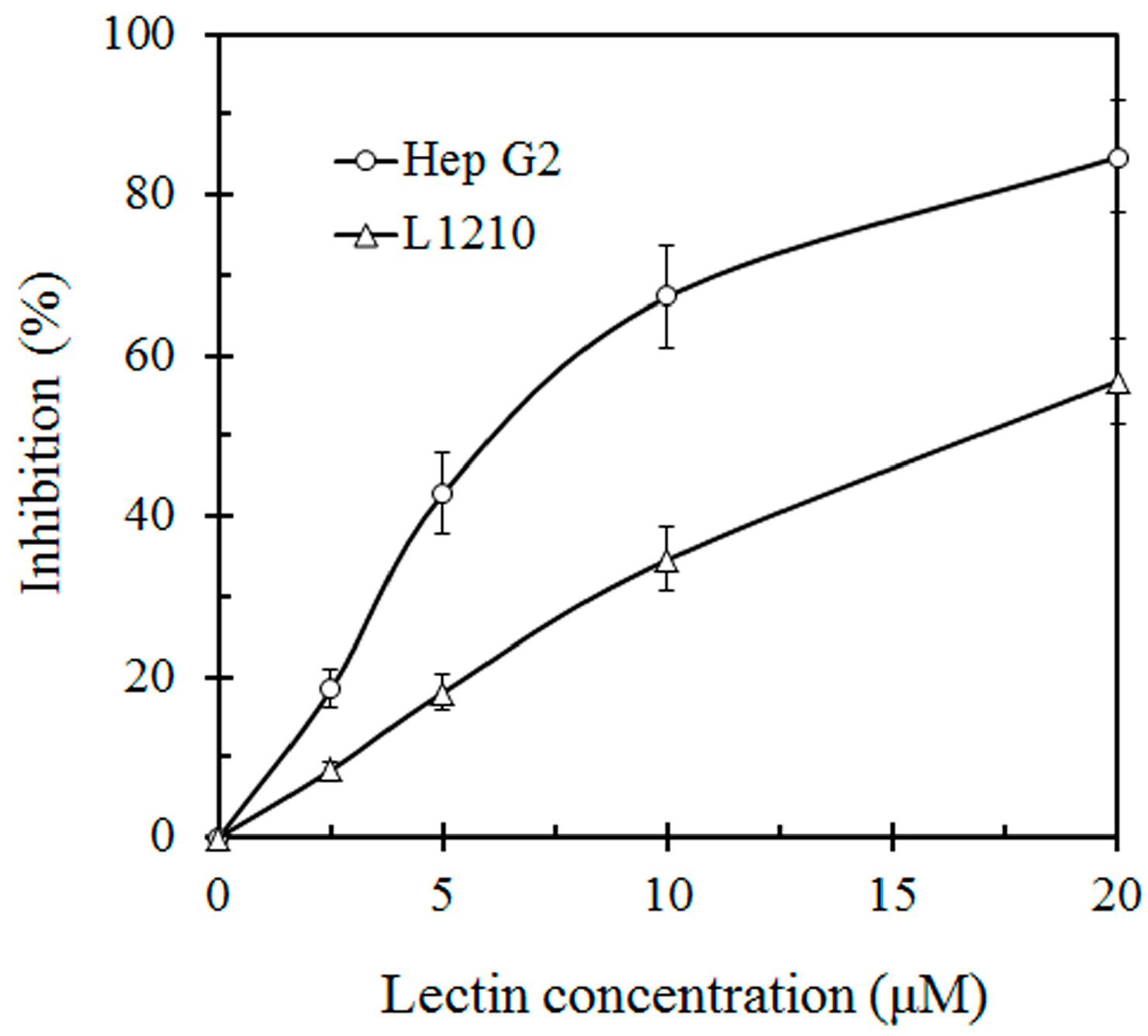{kind=link}
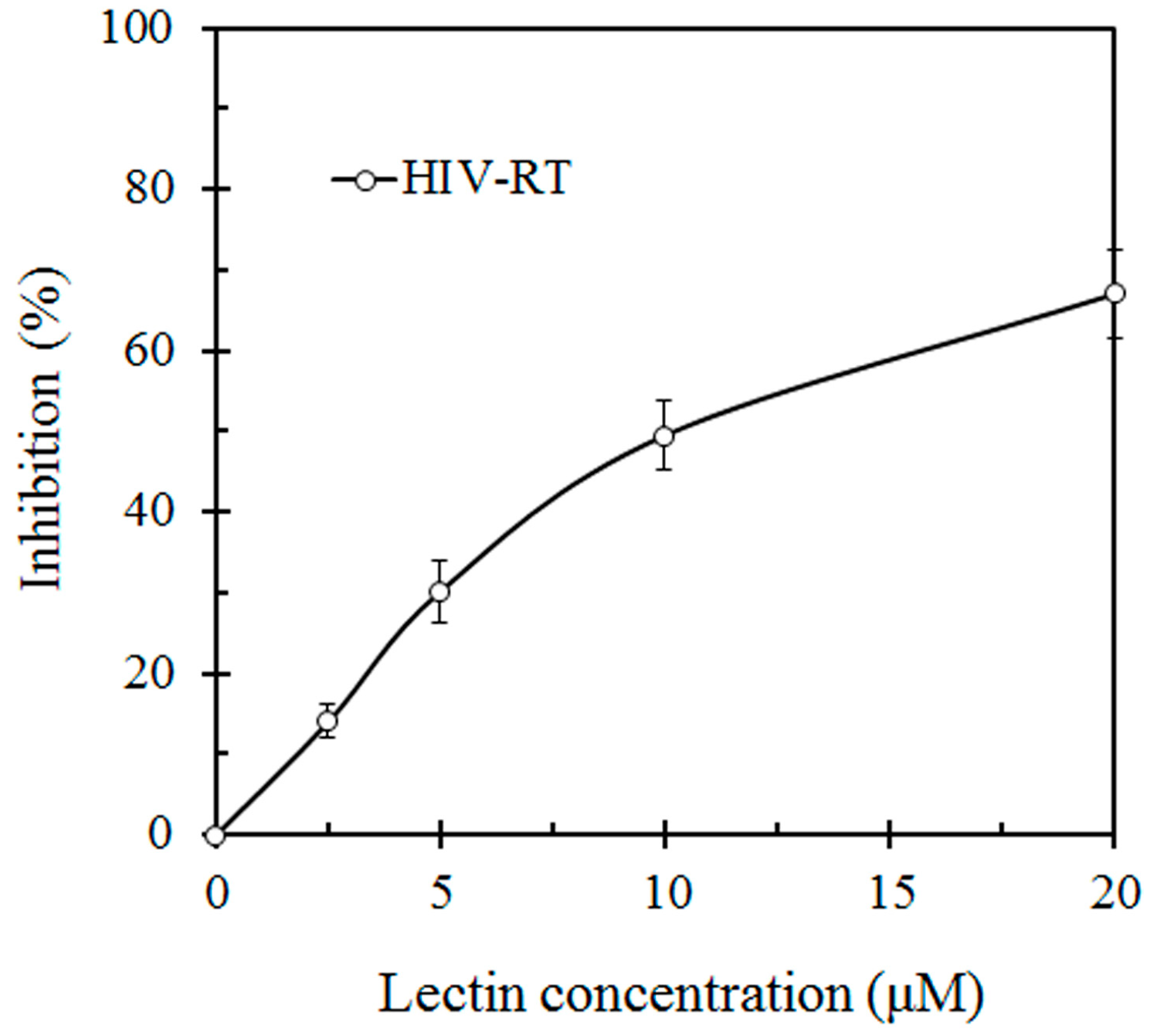{kind=link}
| Fraction | Yield (mg) | Specific Activity (U/mg) | Total Activity (U) | Recovery of Activity (%) | Folds of Purification |
|---|---|---|---|---|---|
| CM | 11,203.8 | 0.7 | 8105.0 | 100.0 | 1.0 |
| CM1 | 2125.7 | - | - | - | - |
| CM2 | 280.5 | 11.3 | 3170.6 | 39.1 | 16.1 |
| CM3 | 1286.3 | - | - | - | - |
| Q1 | 51. | - | - | - | - |
| Q2 | 44.6 | 44.4 | 1978.2 | 24.4 | 63.4 |
| Q3 | 46.9 | - | - | - | - |
| S1 | 11.9 | - | - | - | - |
| S2 | 12.3 | 117.8 | 1453.6 | 17.9 | 168.3 |
| S3 | 5.6 | - | - | - | - |
| SU1 | 2.0 | 320.0 | 640.0 | 7.9 | 457.1 |
| SU2 | 1.2 | - | - | - | - |
| Temperature (°C) | 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | 90 | 100 |
|---|---|---|---|---|---|---|---|---|---|---|
| Hemagglutinating activity (U) | 64 | 64 | 64 | 64 | 64 | 64 | 32 | 16 | 16 | 0 |
| The Hemagglutinating Activity of SRL in Different Concentrations of HCl | ||||||
|---|---|---|---|---|---|---|
| HCl concentration (M) | 6 (pH 2.2) | 12.5 (pH 1.9) | 25 (pH 1.6) | 50 (pH 1.3) | 100 (pH 1.0) | 200 (pH 0.7) |
| Hemagglutinating Activity (U) | 64 | 64 | 32 | 8 | 0 | 0 |
| The Hemagglutinating Activity of SRL in Different Concentrations of NaOH | ||||||
| NaOH concentration (M) | 6 (pH 11.7) | 12.5 (pH 12.1) | 25 (pH 12.4) | 50 (pH 12.7) | 100 (pH 13.0) | 200 (pH 13.3) |
| Hemagglutinating Activity (U) | 64 | 32 | 16 | 8 | 0 | 0 |
| Cations | 10 mM | 5 mM | 2.5 mM | 1.25 mM |
|---|---|---|---|---|
| Cd2+ | 0 | 0 | 2 | 4 |
| Fe3+ | 64 | 64 | 64 | 64 |
| Cu2+ | 64 | 64 | 64 | 64 |
| Hg2+ | 0 | 0 | 2 | 4 |
| Al3+ | 64 | 64 | 64 | 64 |
| Pb2+ | 4 | 16 | 32 | 64 |
| Zn2+ | 32 | 32 | 64 | 64 |
| Ca2+ | 32 | 64 | 64 | 64 |
| Mn2+ | 32 | 64 | 64 | 64 |
| Mg2+ | 64 | 64 | 64 | 64 |
| K+ | 64 | 64 | 64 | 64 |
| Fe2+ | 0 | 4 | 16 | 64 |
| Sugar | Minimum Inhibitory Concentration of Sugar (mM) |
|---|---|
| l-Sorbose | No |
| Raffinose | No |
| l-Rhamnose | No |
| d-Fructose | No |
| d-Mannose | No |
| Cellobiose | No |
| l-Arabinose | No |
| d-Xylose | No |
| d-Melibiose | No |
| Lactose | No |
| Inulin | 0.1 M |
| Maltose | No |
| d-Galactose | No |
| d-Glucose | No |


2.2. Discussion
| Species | N-Terminal Sequence |
|---|---|
| Stropharia rugosoannulata | IKSGVYRIVSWQGALGPEAR |
| Agaricus bisporus [21] | MGGSGTSGSL |
| Agrocybe aegerita [22] | NISAGTSVDL |
| Coprinopsis cinerea [23] | IPLEGTFGDR |
| Flammulina velutipes [24] | TSLTFQLAYL |
| Laccaria bicolor [25] | SHLYGDGVAL |
| Pleurotus cornucopiae [26] | SDSTWTFAML |
3. Experimental Section
3.1. Isolation of Lectins
3.2. Determination of Molecular Mass and N-Terminal Sequence
3.3. Assay for Hemagglutinating Activity
3.4. Assay of Antiproliferative Activity on Tumor Cell Lines
3.5. Assay for HIV-1 Reverse Transcriptase Inhibitory Activity
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lam, S.K.; Ng, T.B. Lectins: Production and practical applications. Appl. Microbiol. Biotechnol. 2011, 89, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.H.; Chan, H.Y.; Ng, T.B. A mannose/glucose-specific lectin from chinese evergreen chinkapin (castanopsis chinensis). Biochim. Biophys. Acta 2008, 1780, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Shao, B.; Wang, S.; Zhou, J.; Ke, L.; Rao, P. A novel lectin from fresh rhizome of alisma orientale (sam.) juzep. Process. Biochem. 2011, 46, 1554–1559. [Google Scholar] [CrossRef]
- Terada, T.; Watanabe, Y.; Tateno, H.; Naganuma, T.; Ogawa, T.; Muramoto, K.; Kamiya, H. Structural characterization of a rhamnose-binding glycoprotein (lectin) from spanish mackerel (Scomberomorous niphonius) eggs. Biochim. Biophys. Acta-Gen. Subj. 2007, 1770, 617–629. [Google Scholar] [CrossRef]
- Ng, T.B.; Lam, Y.W.; Woo, N.Y.S. The immunostimulatory activity and stability of grass carp (Ctenopharyngodon idellus) roe lectin. Vet. Immunol. Immunopathol. 2003, 94, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Sudakevitz, D.; Imberty, A.; Gilboa-Garber, N. Production, properties and specificity of a new bacterial l-fucose- and d-arabinose-binding lectin of the plant aggressive pathogen Ralstonia solanacearum, and its comparison to related plant and microbial lectins. J. Biochem. 2002, 132, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.K.; Wang, H.X.; Ng, T.B. Purification and characterization of a novel lectin from the toxic wild mushroom Inocybe umbrinella. Toxicon 2009, 53, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.R.; Liu, Q.H.; Wang, H.X.; Ng, T.B. A novel lectin with potent antitumor, mitogenic and hiv-1 reverse transcriptase inhibitory activities from the edible mushroom Pleurotus citrinopileatus. Biochim. Biophys. Acta 2008, 1780, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, J.; Wang, H.; Ng, T.B. First isolation and characterization of a novel lectin with potent antitumor activity from a Russula mushroom. Phytomedicine 2010, 17, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Diaz, E.M.; Vicente-Manzanares, M.; Sacristan, M.; Vicente, C.; Legaz, M.E. Fungal lectin of Peltigera canina induces chemotropism of compatible nostoc cells by constriction-relaxation pulses of cyanobiont cytoskeleton. Plant Signal. Behav. 2011, 6, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Varrot, A.; Basheer, S.M.; Imberty, A. Fungal lectins: Structure, function and potential applications. Curr. Opin. Struct. Biol. 2013, 23, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B. Peptides and proteins from fungi. Peptides 2004, 25, 1055–1073. [Google Scholar] [CrossRef]
- Guillot, J.; Konska, G. Lectins in higher fungi. Biochem. Syst. Ecol. 1997, 25, 203–230. [Google Scholar] [CrossRef]
- He, P.X.; Geng, L.J.; Wang, J.Z.; Xu, C.P. Production, purfication, molecular characterization and bioactivities of exopolysaccharides produced by the wine cap culinary-medicinal mushroom, Stropharia rugosoannulata 2# (higher basidiomycetes). Int. J. Med. Mushrooms 2012, 14, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Tokuyama, S.; Nagai, K.; Yasuda, N.; Noguchi, K.; Matsumoto, T.; Hirai, H.; Kawagishi, H. Strophasterols A to D with an unprecedented steroid skeleton: From the mushroom Stropharia rugosoannulata. Angew. Chem. Int. Ed. Engl. 2012, 51, 10820–10822. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fushimi, K.; Tokuyama, S.; Ohno, M.; Miwa, T.; Koyama, T.; Yazawa, K.; Nagai, K.; Matsumoto, T.; Hirai, H.; et al. Functional-food constituents in the fruiting bodies of Stropharia rugosoannulata. Biosci. Biotechnol. Biochem. 2011, 75, 1631–1634. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kobori, H.; Kawaide, M.; Suzuki, T.; Choi, J.H.; Yasuda, N.; Noguchi, K.; Matsumoto, T.; Hirai, H.; Kawagishi, H. Isolation of bioactive steroids from the Stropharia rugosoannulata mushroom and absolute configuration of strophasterol b. Biosci. Biotechnol. Biochem. 2013, 77, 1779–1781. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Liu, Q.H.; Ng, T.B.; Liu, H.Z.; Li, J.Q.; Chen, G.; Sheng, H.Y.; Xie, Z.L.; Wang, H.X. Isolation and characterization of a novel lectin from the mushroom Armillaria luteo-virens. Biochem. Biophys. Res. Commun. 2006, 345, 1573–1578. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B.; Liu, Q.H. Isolation of a new heterodimeric lectin with mitogenic activity from fruiting bodies of the mushroom Agrocybe cylindracea. Life Sci. 2002, 70, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B.; Ooi, V.E.C. Lectins from mushrooms. Mycol. Res. 1998, 102, 897–906. [Google Scholar] [CrossRef]
- Crenshaw, R.W.; Harper, S.N.; Moyer, M.; Privalle, L.S. Isolation and characterization of a cdna clone encoding a lectin gene from agaricus-bisporus. Plant Physiol. 1995, 107, 1465–1466. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.G.; Sun, H.; Tong, X.; Qi, Y.P. An antitumour lectin from the edible mushroom Agrocybe aegerita. Biochem. J. 2003, 374, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Walti, M.A.; Walser, P.J.; Thore, S.; Grunler, A.; Bednar, M.; Kunzler, M.; Aebi, M. Structural basis for chitotetraose coordination by cgl3, a novel galectin-related protein from Coprinopsis cinerea. J. Mol. Biol. 2008, 379, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.L.; Hsu, C.I.; Lin, R.H.; Kao, C.L.; Lin, J.Y. A new fungal immunomodulatory protein, fip-fve isolated from the edible mushroom, Flammulina velutipes and its complete amino-acid-sequence. Eur. J. Biochem. 1995, 228, 244–249. [Google Scholar] [CrossRef]
- Martin, F.; Aerts, A.; Ahren, D.; Brun, A.; Danchin, E.G.J.; Duchaussoy, F.; Gibon, J.; Kohler, A.; Lindquist, E.; Pereda, V.; et al. The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis. Nature 2008, 452, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Oguri, S.; Ando, A.; Nagata, Y. A novel developmental stage-specific lectin of the basidiomycete Pleurotus cornucopiae. J. Bacteriol. 1996, 178, 5692–5698. [Google Scholar] [PubMed]
- Zheng, S.Y.; Li, C.X.; Ng, T.B.; Wang, H.X. A lectin with mitogenic activity from the edible wild mushroom Boletus edulis. Process Biochem. 2007, 42, 1620–1624. [Google Scholar] [CrossRef]
- Han, C.H.; Liu, Q.H.; Ng, T.B.; Wang, H.X. A novel homodimeric lactose-binding lectin from the edible split gill medicinal mushroom Schizophyllum commune. Biochem. Biophys. Res. Commun. 2005, 336, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B.; Liu, W.K.; Ooi, V.E.C.; Chang, S.T. Isolation and characterization of 2 distinct lectins with antiproliferative activity from the cultured mycelium of the edible mushroom Tricholoma mongolicum. Int. J. Pept. Protein Res. 1995, 46, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.Q.; Sun, J.; Wang, H.X.; Ng, T.B. A novel lectin with antiproliferative activity from the medicinal mushroom Pholiota adiposa. Acta Biochim. Pol. 2009, 56, 415–421. [Google Scholar] [PubMed]
- Liu, Q.H.; Wang, H.X.; Ng, T.B. Isolation and characterization of a novel lectin from the wild mushroom Xerocomus spadiceus. Peptides 2004, 25, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage-T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Tian, G.; Geng, X.; Zhao, Y.; Ng, T.B.; Zhao, L.; Wang, H. Isolation and Characterization of a Novel Lectin from the Edible Mushroom Stropharia rugosoannulata. Molecules 2014, 19, 19880-19891. https://doi.org/10.3390/molecules191219880
Zhang W, Tian G, Geng X, Zhao Y, Ng TB, Zhao L, Wang H. Isolation and Characterization of a Novel Lectin from the Edible Mushroom Stropharia rugosoannulata. Molecules. 2014; 19(12):19880-19891. https://doi.org/10.3390/molecules191219880
Chicago/Turabian StyleZhang, Weiwei, Guoting Tian, Xueran Geng, Yongchang Zhao, Tzi Bun Ng, Liyan Zhao, and Hexiang Wang. 2014. "Isolation and Characterization of a Novel Lectin from the Edible Mushroom Stropharia rugosoannulata" Molecules 19, no. 12: 19880-19891. https://doi.org/10.3390/molecules191219880
APA StyleZhang, W., Tian, G., Geng, X., Zhao, Y., Ng, T. B., Zhao, L., & Wang, H. (2014). Isolation and Characterization of a Novel Lectin from the Edible Mushroom Stropharia rugosoannulata. Molecules, 19(12), 19880-19891. https://doi.org/10.3390/molecules191219880