Different Accumulation Profiles of Multiple Components Between Pericarp and Seed of Alpinia oxyphylla Capsular Fruit as Determined by UFLC-MS/MS

Abstract
:1. Introduction

2. Results and Discussion
2.1. Selection of the Extraction Method
2.2. Analytical Method Validation
2.3. Quantitative Analysis of Seed and Pericarp Samples


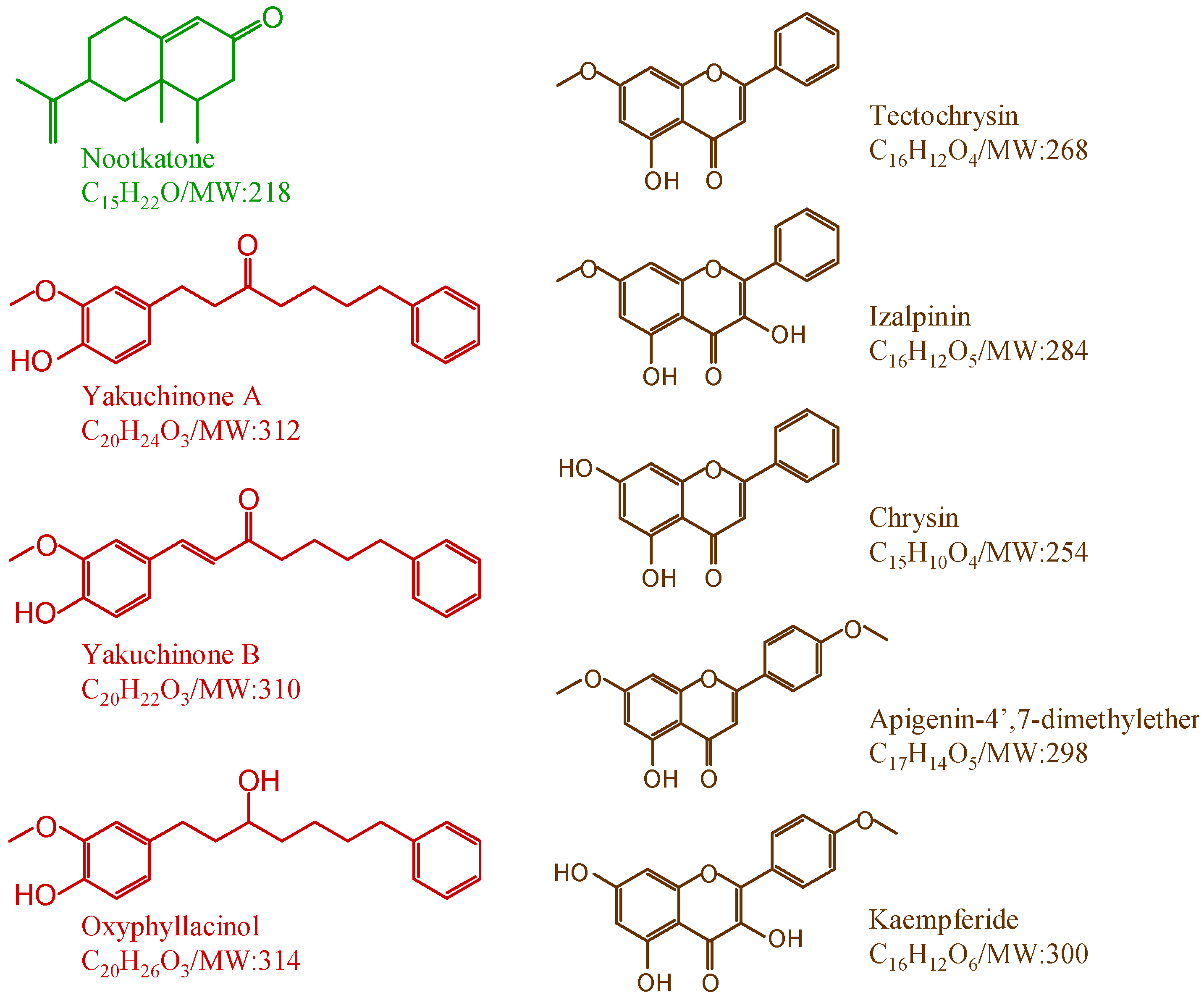{kind=link}
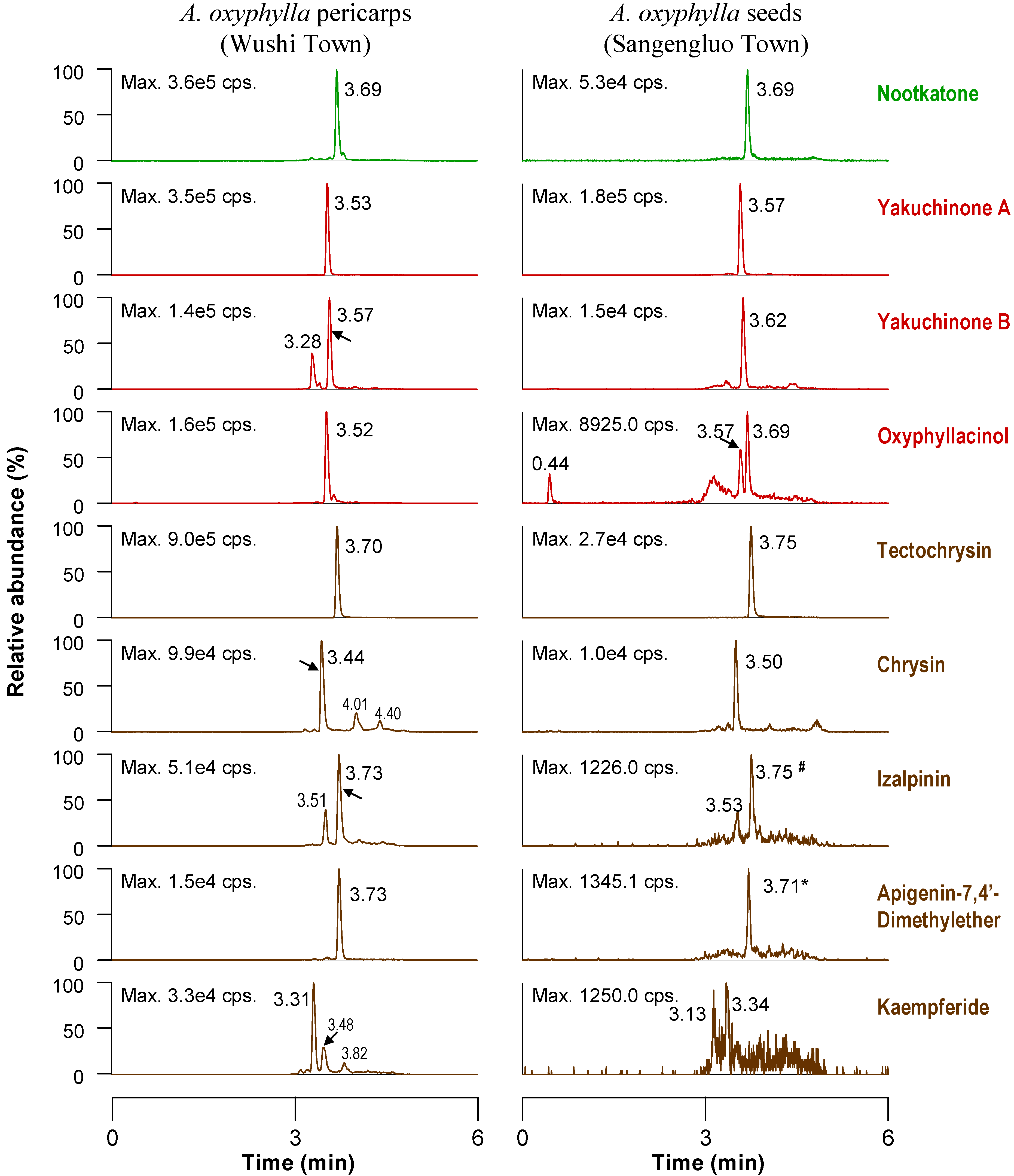{kind=link}
{kind=link}
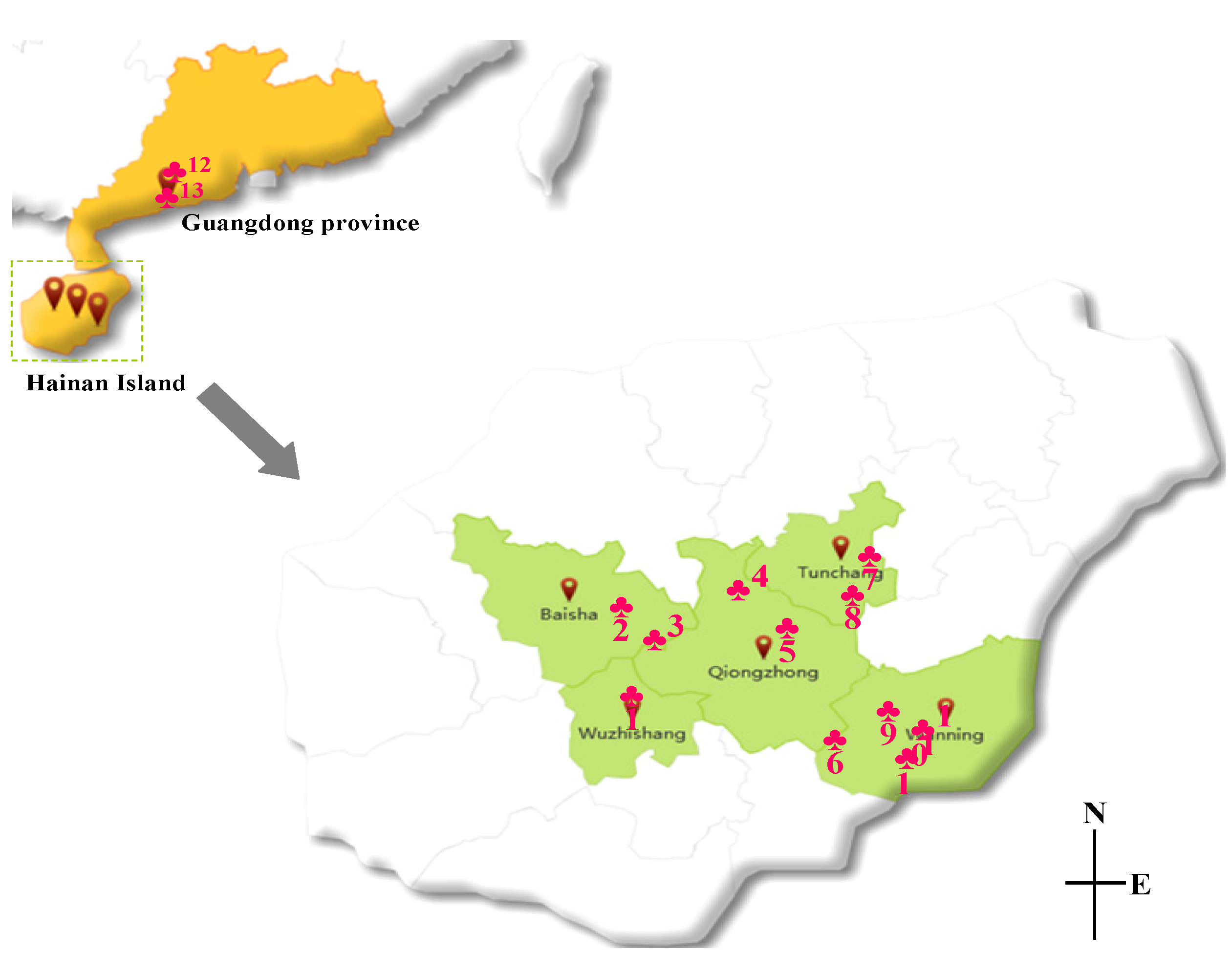{kind=link}
| Compound | Nootkatone | Yakuchinone A | Yakuchinone B | Oxyphyllacinol | Tectochrysin | Izalpinin | Chrysin | Kaempferide | Apigenin-7,4'-dimethylether |
|---|---|---|---|---|---|---|---|---|---|
| Pericarps of A. oxyphylla | |||||||||
| Precision (RSD, %) | |||||||||
| Intraday (n = 6) | 1.63 | 6.59 | 1.23 | 6.00 | 2.31 | 4.64 | 4.31 | 7.02 | 2.75 |
| Interday (n = 15) | 2.74 | 10.7 | 1.81 | 4.18 | 2.97 | 5.28 | 8.11 | 7.99 | 3.39 |
| Recovery | |||||||||
| Mean ± SD, %, n = 6 | 105.8 ± 3.1 | 100.0 ± 9.9 | 105.4 ± 5.3 | 103.2 ± 4.2 | 107.6 ± 3.8 | 93.34 ± 5.88 | 102.0 ± 8.8 | 96.91 ± 3.26 | 97.13 ± 6.12 |
| RSD (%) | 2.93 | 9.90 | 5.03 | 4.07 | 3.53 | 6.30 | 8.63 | 3.36 | 6.30 |
| Seeds A. oxyphylla | |||||||||
| Precision (RSD, %) | |||||||||
| Intraday (n = 6) | 4.08 | 2.62 | —— a | 3.48 | 1.20 | —— | 4.67 | —— | —— |
| Interday (n = 15) | 6.34 | 4.40 | —— | 6.74 | 5.47 | —— | 4.28 | —— | —— |
| Recovery | |||||||||
| Mean ± SD, %, n = 6 | 95.67 ± 5.57 | 101.8 ± 4.23 | —— | 98.40 ± 3.59 | 106.7 ± 3.05 | —— | 96.48 ± 5.56 | —— | —— |
| RSD (%) | 5.82 | 4.16 | —— | 3.65 | 2.86 | —— | 5.76 | —— | —— |
| Production regions | Nootkatone | Yakuchinone A | Yakuchinone B | Oxyphyllacinol | Tectochrysin | Izalpinin | Chrysin | Kaempferide | Apigenin-7,4'-dimethylether |
|---|---|---|---|---|---|---|---|---|---|
| Pericarps (1) | 46.8 ± 4.2 | 2961 ± 164 | 21.1 ± 0.8 | 2579 ± 106 | 201 ± 9 | 47.9 ± 1.6 | 60.9 ± 1.9 | 20.1 ± 0.9 | 268 ± 15 |
| Pericarps (2) | 35.1 ± 3.4 | 6440 ± 282 | 63.6 ± 7.6 | 5519 ± 177 | 139 ± 17 | 32.5 ± 2.3 | 165 ± 18 | 27.6 ± 2.5 | 242 ± 35 |
| Pericarps (3) | 58.5 ± 3.9 | 2913 ± 149 | 23.3 ± 0.7 | 2552 ± 228 | 278 ± 11 | 64.5 ± 3.2 | 69.0 ± 3.8 | 25.6 ± 0.6 | 186 ± 7 |
| Pericarps (4) | 48.9 ± 6.2 | 4146 ± 270 | 35.0 ± 3.6 | 3666 ± 342 | 170 ± 24 | 42.0 ± 4.2 | 145 ± 8 | 38.9 ± 2.9 | 304 ± 29 |
| Pericarps (5) | 158 ± 17 | 5390 ± 752 | 45.3 ± 6.3 | 4647 ± 514 | 266 ± 5 | 59.7 ± 3.4 | 245 ± 21 | 36.2 ± 2.7 | 308 ± 6 |
| Pericarps (6) | 15.8 ± 1.0 | 1772 ± 62 | 90.5 ± 5.8 | 1699 ± 113 | 187 ± 9 | 30.7 ± 2.0 | 64.2 ± 2.7 | 23.9 ± 1.5 | 519 ± 28 |
| Pericarps (7) | 13.0 ± 0.4 | 3473 ± 449 | 91.5 ± 91.5 | 3177 ± 308 | 208 ± 15 | 33.5 ± 1.9 | 109 ± 7 | 12.4 ± 0.8 | 232 ± 23 |
| Pericarps (8) | 6.56 ± 0.04 | 2799 ± 339 | 110 ± 3 | 2724 ± 287 | 305 ± 14 | 38.3 ± 0.9 | 76.1 ± 5.6 | 14.9 ± 1.0 | 796 ± 14 |
| Pericarps (9) | 48.9 ± 6.2 | 3853 ± 345 | 44.9 ± 3.0 | 3641 ± 118 | 180 ± 12 | 38.6 ± 2.2 | 137 ± 8 | 30.1 ± 1.5 | 325 ± 23 |
| Pericarps (10) | 6.28 ± 0.22 | 5916 ± 600 | 66.4 ± 4.0 | 5230 ± 232 | 138 ± 10 | 31.1 ± 1.7 | 101 ± 11 | 23.2 ± 2.5 | 298 ± 20 |
| Pericarps (11) | 25.5 ± 1.9 | 1114 ± 240 | 75.0 ± 5.3 | 1114 ± 80 | 192 ± 8 | 21.4 ± 1.5 | 98.8 ± 3.3 | 19.8 ± 0.4 | 378 ± 17 |
| Pericarps (12) | 15.2 ± 1.2 | 6657 ± 504 | 16.4 ± 0.5 | 5656 ± 485 | 213 ± 4 | 66.7 ± 2.5 | 212 ± 8 | 18.4 ± 0.1 | 121 ± 5 |
| Pericarps (13) | 59.8 ± 2.9 | 3073 ± 189 | 98.5 ± 3.4 | 2973 ± 112 | 186 ± 2 | 30.2 ± 1.6 | 98.6 ± 5 | 20.0 ± 0.4 | 441 ± 20 |
| Production regions | Nootkatone | Yakuchinone A | Yakuchinone B | Oxyphyllacinol | Tectochrysin | Izalpinin | Chrysin | Kaempferide | Apigenin-7,4'-dimethylether |
|---|---|---|---|---|---|---|---|---|---|
| Seeds (1) | 2416 ± 134 | 2.74 ± 0.14 | —— | 7.77 ± 0.13 | 0.77 ± 0.05 | —— | 0.55 ± 0.04 | —— | —— |
| Seeds (2) | 3028 ± 77 | 0.90 ± 0.03 | —— | 6.64 ± 0.09 | 0.41 ± 0.03 | —— | 0.59 ± 0.02 | —— | —— |
| Seeds (3) | 2888 ± 217 | 1.13 ± 0.03 | —— | 5.05 ± 0.38 | 0.44 ± 0.02 | —— | 0.53 ± 0.04 | —— | —— |
| Seeds (4) | 3098 ± 127 | 1.11 ± 0.00 | —— | 5.36 ± 0.26 | —— | —— | 0.64 ± 0.01 | —— | —— |
| Seeds (5) | 2945 ± 269 | 1.24 ± 0.07 | —— | 6.56 ± 0.26 | 0.65 ± 0.01 | —— | 1.80 ± 0.09 | —— | —— |
| Seeds (6) | 3692 ± 197 | —— a | —— | 5.63 ± 0.12 | —— | —— | —— | —— | —— |
| Seeds (7) | 2878 ± 171 | 2.94 ± 0.02 | —— | 5.63 ± 0.09 | 0.39 ± 0.02 | —— | 0.76 ± 0.02 | —— | —— |
| Seeds (8) | 2522 ± 48 | 0.40 ± 0.03 | —— | 2.61 ± 0.11 | 0.40 ± 0.02 | —— | —— | —— | 1.85 ± 0.09 |
| Seeds (9) | 2700 ± 112 | 3.21 ± 0.09 | 0.46 ± 0.01 | 9.85 ± 0.30 | 0.82 ± 0.01 | —— | 1.84 ± 0.05 | —— | 1.47 ± 0.06 |
| Seeds (10) | 3095 ± 31 | 0.43 ± 0.05 | —— | 5.91 ± 0.22 | —— | —— | 0.43 ± 0.01 | —— | —— |
| Seeds (11) | 2750 ± 36 | 1.11 ± 0.00 | —— | 4.22 ± 0.11 | 0.61 ± 0.05 | —— | 0.43 ± 0.02 | —— | 1.00 ± 0.04 |
| Seeds (12) | 1898 ± 117 | 3.90 ± 0.05 | —— | 8.54 ± 0.27 | —— | —— | 0.60 ± 0.05 | —— | —— |
| Seeds (13) | 2501 ± 105 | 5.17 ± 0.10 | —— | 10.7 ± 0.22 | 1.08 ± 0.03 | —— | 1.39 ± 0.03 | —— | 2.03 ± 0.10 |

3. Experimental
3.1. Chemicals and Materials
3.2. Preparation of Sample Solutions

3.3. Analysis of the Nine Secondary Metabolites Occurring in Seeds and Pericarps of A. oxyphylla
3.4. Method Validation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef]
- Yazaki, K. ABC transporters involved in the transport of plant secondary metabolites. FEBS Lett. 2006, 580, 1183–1191. [Google Scholar] [CrossRef]
- Harborne, J.B. Plant Secondary Metabolism, in Plant Ecology, 2nd ed.; Crawley, M.J., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2009. [Google Scholar] [CrossRef]
- Bourgaud, F.; Gravot, A.; Milesi, S.; Gontier, E. Production of plant secondary metabolites: A historical perspective. Plant Sci. 2001, 161, 839–851. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2010; Volume 1, pp. 1–274. [Google Scholar]
- But, P.P.H.; Kimura, T.; Guo, J.X.; Sung, C.K.; Han, B.H. International Collation of Traditional and Folk Medicine: Northeast Asia Part II; World Scientific: Singapore, 1997; pp. 202–203. [Google Scholar]
- Zhang, J.Q.; Wang, S.; Li, Y.H.; Xu, P.; Chen, F.; Tan, Y.; Duan, J.A. Anti-diarrheal constituents of Alpinia oxyphylla. Fitoterapia 2013, 89, 149–156. [Google Scholar] [CrossRef]
- Chen, F.; Li, H.L.; Li, Y.H.; Tan, Y.F.; Zhang, J.Q. Quantitative analysis of the major constituents in Chinese medicinal preparation SuoQuan formulae by ultra fast high performance liquid chromatography/quadrupole tandem mass spectrometry. Chem. Cent. J. 2013, 7. [Google Scholar] [CrossRef]
- Chun, K.S.; Kang, J.Y.; Kim, O.H.; Kang, H.; Surh, Y.J. Effects of yakuchinone A and yakuchinone B on the phorbol ester-induced expression of COX-2 and iNOS and activation of NF-kappaB in mouse skin. J. Environ. Pathol. Toxicol. Oncol. 2002, 21, 131–139. [Google Scholar]
- Chun, K.S.; Park, K.K.; Lee, J.; Kang, M.; Surh, Y.J. Inhibition of mouse skin tumor promotion by anti-inflammatory diarylheptanoids derived from Alpinia oxyphylla Miquel (Zingiberaceae). Oncol. Res. 2002, 13, 37–45. [Google Scholar]
- Shin, T.Y.; Won, J.H.; Kim, H.M.; Kim, S.H. Effect of Alpinia oxyphylla fruit extract on compound 48/80-induced anaphylactic reactions. Am. J. Chin. Med. 2001, 29, 293–302. [Google Scholar] [CrossRef]
- Yamahara, J.; Li, Y.H.; Tamai, Y. Anti-ulcer effect in rats of bitter cardamom constituents. Chem. Pharm. Bull. 1990, 38, 3053–3054. [Google Scholar] [CrossRef]
- Yu, X.Y.; An, L.J.; Wang, Y.Q.; Zhao, H.; Gao, C.Z. Neuroprotective effect of Alpinia oxyphylla Miq. fruits against glutamate-induced apoptosis in cortical neurons. Toxicol. Lett. 2003, 144, 205–212. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Cheang, L.C.; Wang, M.W.; Li, G.H.; Chu, I.K.; Lin, Z.X.; Lee, S.M. Ethanolic extract of fructus Alpinia oxyphylla protects against 6-hydroxydopamine-induced damage of PC12 cells in vitro and dopaminergic neurons in zebrafish. Cell. Mol. Neurobiol. 2012, 32, 27–40. [Google Scholar] [CrossRef]
- Song, W.J.; Li, Y.H.; Wang, J.G.; Li, Z.Y.; Zhang, J,Q. Characterization of nucleobases and nucleosides in the fruit of Alpinia oxyphylla collected from different cultivation regions. Drug Test. Anal. 2014, 6, 239–245. [Google Scholar] [CrossRef]
- Qing, Z.J.; Yong, W.; Hui, L.Y.; Yong, L.W.; Long, L.H.; Ao, D.J.; Xia, P.L. Two new natural products from the fruits of Alpinia oxyphylla with inhibitory effects on nitric oxide production in lipopolysaccharideactivated RAW264.7 macrophage cells. Arch. Pharm. Res. 2012, 35, 2143–2146. [Google Scholar] [CrossRef]
- Li, Y.H.; Chen, F.; Wang, J.F.; Wang, Y.; Zhang, J.Q.; Guo, T. Analysis of nine compounds from Alpinia oxyphylla fruit at different harvest time using UFLC-MS/MS and an extraction method optimized by orthogonal design. Chem. Cent. J. 2013, 7. [Google Scholar] [CrossRef]
- Bauer, R.; Franz, G. Modern European monographs for quality control of Chinese herbs. Planta Med. 2010, 76, 2004–2011. [Google Scholar] [CrossRef]
- Zhao, S.K.; Huang, Y.Y. Traditional processing for Alpinia oxyphylla capsularfruit. Chin. Tradit. Pat. Med. 1994, 16, 24–25. [Google Scholar]
- Wu, D.L.; Jin, C.S.; Kou, W.Q.; Xu, F.Q.; Zhou, Y. Comparative studies for medicinal parts of Alpinia oxyphylla capsularfruit. Chin. J. Exp. Tradit. Med. Form. 2007, 13, 1–3. [Google Scholar]
- Miyazawa, M.; Nakamura, Y.; Ishikawa, Y. Insecticidal sesquiterpene from Alpinia oxyphylla against Drosophila melanogaster. J. Agric. Food. Chem. 2000, 48, 3639–3641. [Google Scholar] [CrossRef]
- Dietrich, G.; Dolan, M.C.; Peralta-Cruz, J.; Schmidt, J.; Piesman, J.; Eisen, R.J.; Karchesy, J.J. Repellent activity of fractioned compounds from Chamaecyparis nootkatensis essential oil against nymphal Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2006, 43, 957–961. [Google Scholar] [CrossRef]
- Panella, N.A.; Dolan, M.C.; Karchesy, J.J.; Xiong, Y.; Peralta-Cruz, J.; Khasawneh, M.; Montenieri, J.A.; Maupin, G.O. Use of novel compounds for pest control: Insecticidal and acaricidal activity of essential oil components from heartwood of Alaska yellow cedar. J. Med. Entomol. 2005, 42, 352–358. [Google Scholar] [CrossRef]
- Zhu, B.C.; Henderson, G.; Chen, F.; Maistrello, L.; Laine, R.A. Nootkatone is a repellent for Formosan subterranean termite (Coptotermes formosanus). J. Chem. Ecol. 2001, 27, 523–531. [Google Scholar] [CrossRef]
- Maistrello, L.; Henderson, G.; Laine, R.A. Efficacy of vetiver oil and nootkatone as soil barriers against Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Chem. Ecol. 2001, 94, 1532–1537. [Google Scholar]
- Maistrello, L.; Henderson, G.; Laine, R.A. Comparative effects of vetiver oil, nootkatone and disodium octaborate tetrahydrate on Coptotermes formosanus and its symbiotic fauna. Pest. Manag. Sci. 2003, 59, 58–68. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef]
- Lin, R.J.; Yen, C.M.; Chou, T.H.; Chiang, F.Y.; Wang, G.H.; Tseng, Y.P.; Wang, L.; Huang, T.W.; Wang, H,C.; Chan, L.P.; et al. Antioxidant, anti-adipocyte differentiation, antitumor activity and anthelmintic activities against Anisakis simplex and Hymenolepis nana of yakuchinone A from Alpinia oxyphylla. BMC Complement. Altern. Med. 2013, 13. [Google Scholar] [CrossRef]
- Chen, F.; Li, H.L.; Tan, Y.F.; Li, Y.H.; Zhang, J.Q. Validated method to measure yakuchinone A in plasma by LC-MS/MS and its application to a pharmacokinetic study in rats. Chem. Cent. J. 2014, 8. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the nine compounds and botanical extract solutions are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, F.; Li, H.-L.; Tan, Y.-F.; Guan, W.-W.; Zhang, J.-Q.; Li, Y.-H.; Zhao, Y.-S.; Qin, Z.-M. Different Accumulation Profiles of Multiple Components Between Pericarp and Seed of Alpinia oxyphylla Capsular Fruit as Determined by UFLC-MS/MS. Molecules 2014, 19, 4510-4523. https://doi.org/10.3390/molecules19044510
Chen F, Li H-L, Tan Y-F, Guan W-W, Zhang J-Q, Li Y-H, Zhao Y-S, Qin Z-M. Different Accumulation Profiles of Multiple Components Between Pericarp and Seed of Alpinia oxyphylla Capsular Fruit as Determined by UFLC-MS/MS. Molecules. 2014; 19(4):4510-4523. https://doi.org/10.3390/molecules19044510
Chicago/Turabian StyleChen, Feng, Hai-Long Li, Yin-Feng Tan, Wei-Wei Guan, Jun-Qing Zhang, Yong-Hui Li, Yuan-Sheng Zhao, and Zhen-Miao Qin. 2014. "Different Accumulation Profiles of Multiple Components Between Pericarp and Seed of Alpinia oxyphylla Capsular Fruit as Determined by UFLC-MS/MS" Molecules 19, no. 4: 4510-4523. https://doi.org/10.3390/molecules19044510
APA StyleChen, F., Li, H.-L., Tan, Y.-F., Guan, W.-W., Zhang, J.-Q., Li, Y.-H., Zhao, Y.-S., & Qin, Z.-M. (2014). Different Accumulation Profiles of Multiple Components Between Pericarp and Seed of Alpinia oxyphylla Capsular Fruit as Determined by UFLC-MS/MS. Molecules, 19(4), 4510-4523. https://doi.org/10.3390/molecules19044510