High Efficiency Acetylcholinesterase Immobilization on DNA Aptamer Modified Surfaces

Abstract
:1. Introduction
2. Results and Discussion
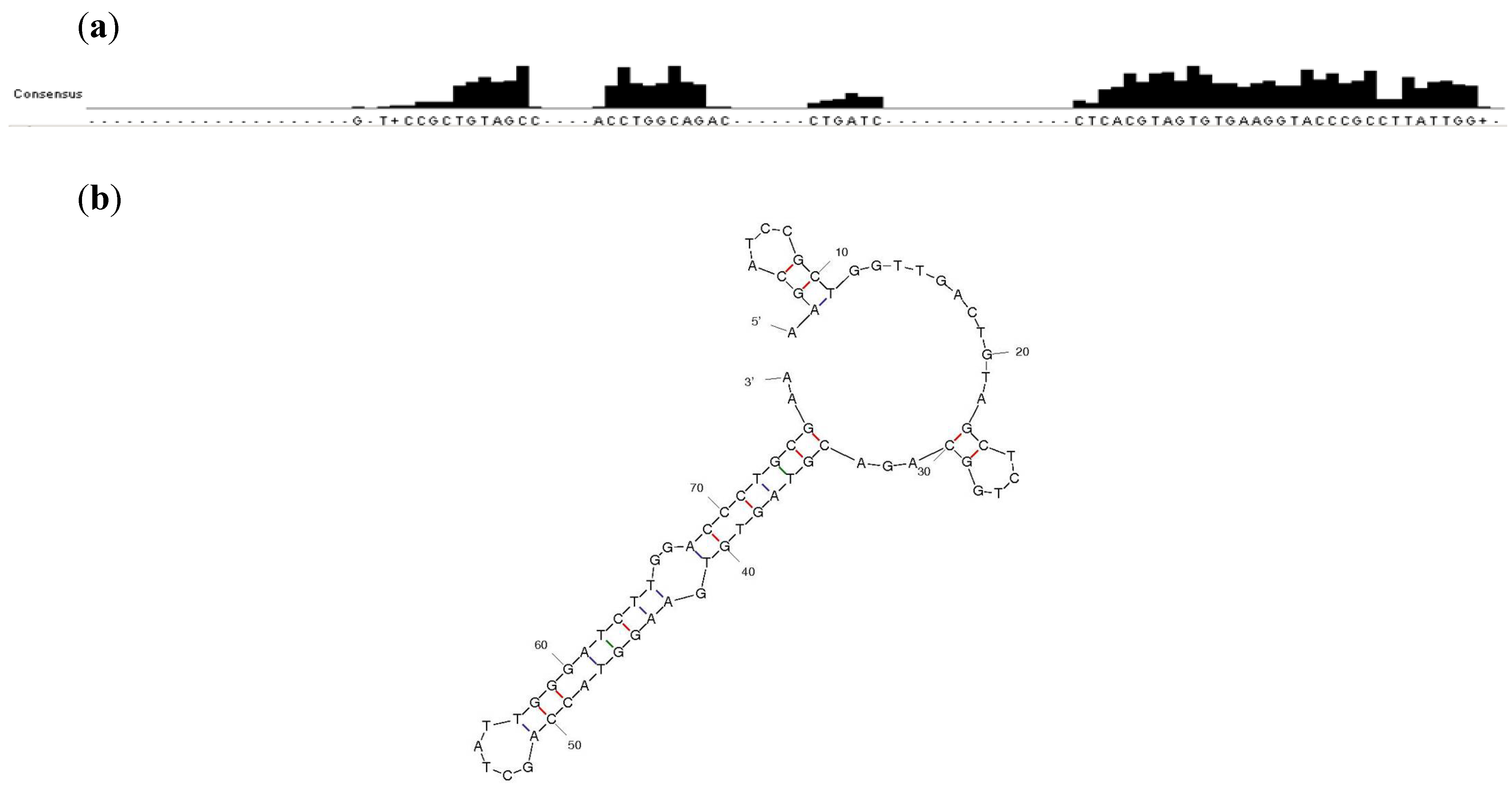
2.1. Sequence Analysis and Binding Characterization


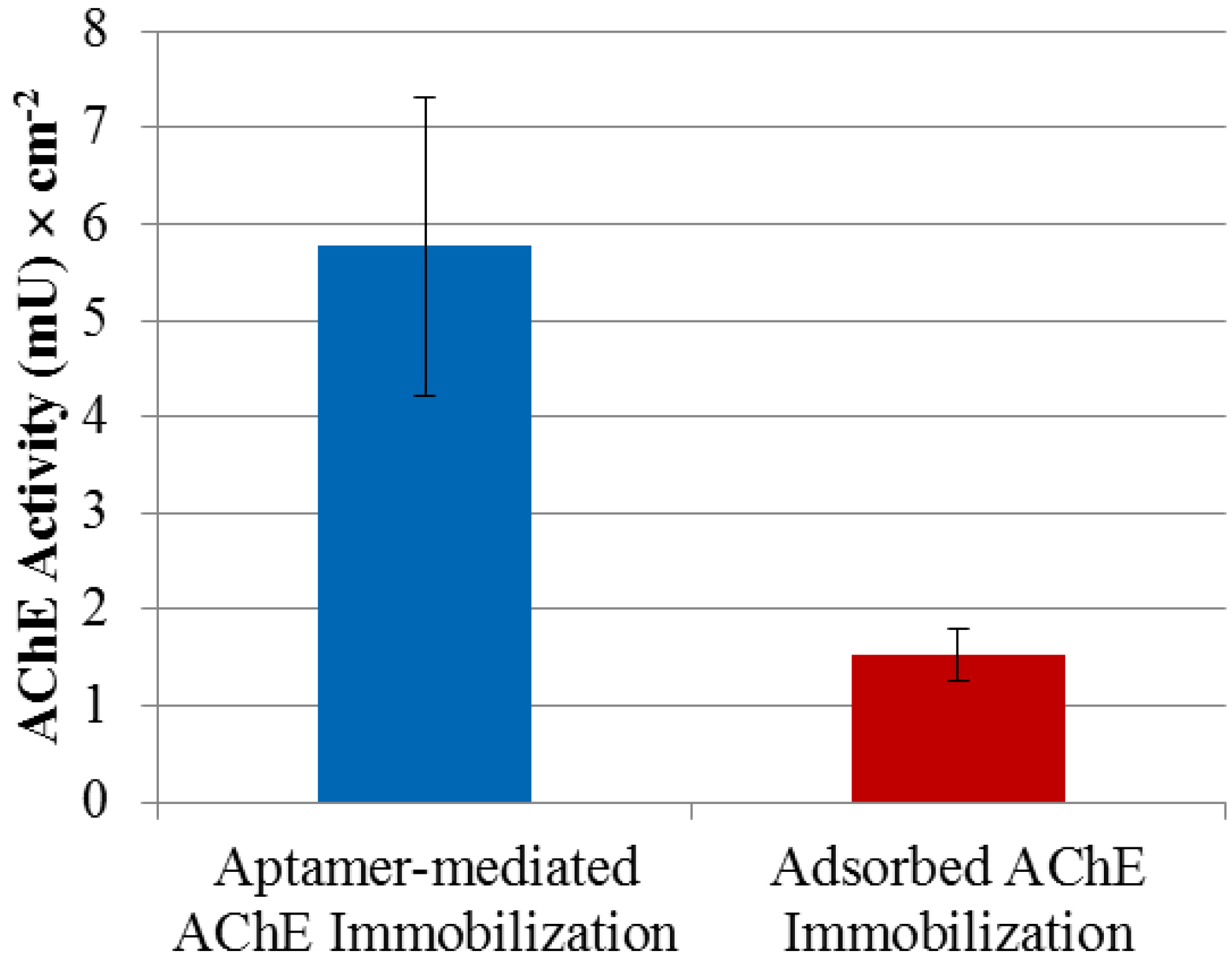
2.2. AChE Enzyme Activity

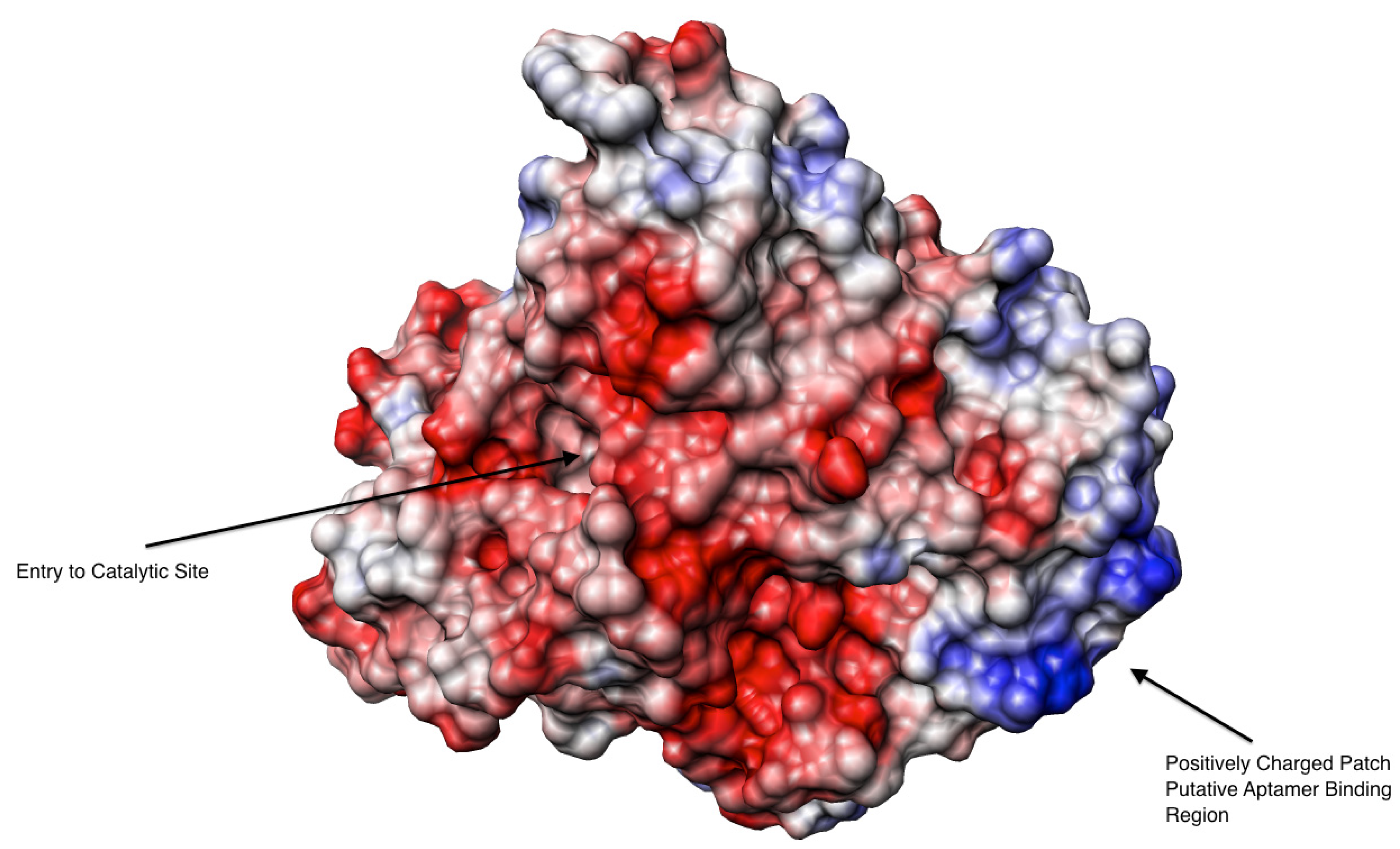
2.3. Binding Affinity of the Aptamer to AChE

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Binding buffer | Ionic strength (M) | Kd (pM) |
|---|---|---|
| Selection buffer (8 mM Na2HPO4, 2 mM NaH2PO4, 2.7 mM KCl, 137 mM NaCl, 4 mM MgCl2, pH 7.4) | ~0.15 | 157 ± 42 |
| Low ionic strength buffer (8 mM Na2HPO4, 2 mM NaH2PO4, 2.7 mM KCl, 4 mM MgCl2, pH 7.4) | ~0.02 | 243 ± 47 |
| High ionic strength buffer (8 mM Na2HPO4, 2 mM NaH2PO4, 2.7 mM KCl, 1,137 mM NaCl, 4 mM MgCl2, pH 7.4) | ~1.2 | (422 ± 69) × 103 |

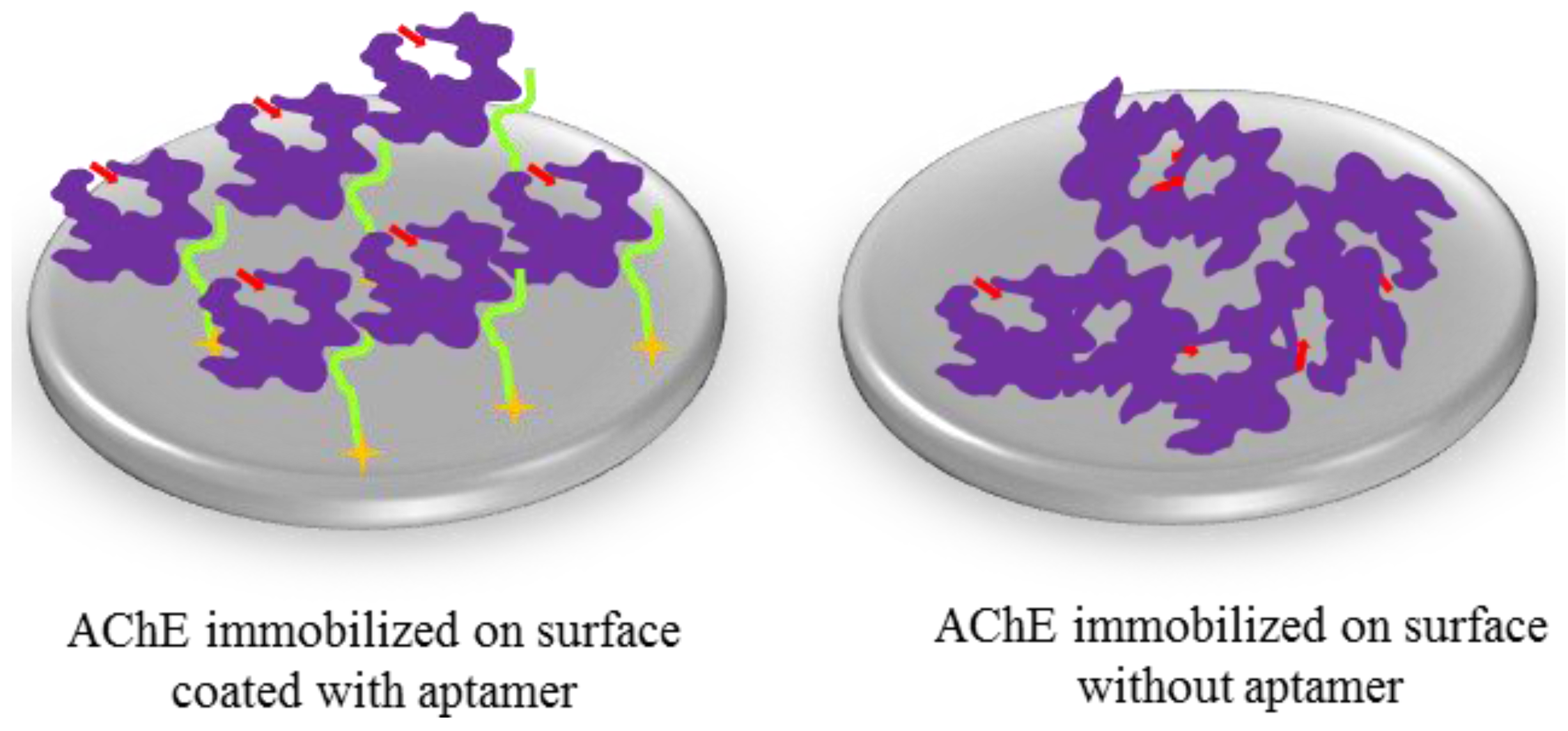
2.4. Immobilization of AChE Using R15/19

3. Experimental Section
3.1. Selection of the Aptamer
3.2. ELONA Assays
3.3. AChE Activity Assays
4. Conclusions
Abbreviations
| AChE | acetylcholinesterase |
| ELONA | enzyme-linked oligonucleotide assay |
| BSA | bovine serum albumin |
| PBST | phosphate buffer saline Tween |
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Keefe, A.D.; Pai, S.; Ellington, A. Aptamers as therapeutics. Nat. Rev. Drug Discov. 2010, 9, 537–550. [Google Scholar] [CrossRef]
- Cho, E.J.; Lee, J.-W.; Ellington, A.D. Applications of Aptamers as Sensors. Ann. Rev. Anal. Chem. 2009, 2, 241–264. [Google Scholar] [CrossRef]
- Jenison, R.; Gill, S.; Pardi, A.; Polisky, B. High-resolution molecular discrimination by RNA. Science 1994, 263, 1425–1429. [Google Scholar]
- Tasset, D.M.; Kubik, M.F.; Steiner, W. Oligonucleotide inhibitors of human thrombin that bind distinct epitopes. J. Mol. Biol. 1997, 272, 688–698. [Google Scholar] [CrossRef]
- Bock, L.C.; Griffin, L.C.; Latham, J.A.; Vermaas, E.H.; Toole, J.J. Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature 1992, 355, 564–566. [Google Scholar] [CrossRef]
- Srisawat, C.; Engelke, D.R. Streptavidin aptamers: Affinity tags for the study of RNAs and ribonucleoproteins. RNA 2001, 7, 632–641. [Google Scholar] [CrossRef]
- Tahiri-Alaoui, A.; Frigotto, L.; Manville, N.; Ibrahim, J.; Romby, P.; James, W. High affinity nucleic acid aptamers for streptavidin incorporated into bi-specific capture ligands. Nucl. Acid. Res. 2002, 30, e45. [Google Scholar] [CrossRef]
- Cass, T.; Ligler, F.S. Immobilized Biomolecules in Analysis: A Practical Approach; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Cooper, J.M.; Cass, A.E.G. Biosensors (A Practical Approach); Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Sheldon, R.A. Enzyme Immobilization: The Quest for Optimum Performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Miyazaki, M. Enzyme-immobilized reactors for rapid and efficient sample preparation in MS-based proteomic studies. Proteomics 2013, 13, 457–466. [Google Scholar] [CrossRef]
- Butler, J.E.; Ni, L.; Brown, W.R.; Joshi, K.S.; Chang, J.; Rosenberg, B.; Voss, E.W., Jr. The immunochemistry of sandwich elisas—VI. Greater than 90% of monoclonal and 75% of polyclonal anti-fluorescyl capture antibodies (CAbs) are denatured by passive adsorption. Mol. Immunol. 1993, 30, 1165–1175. [Google Scholar] [CrossRef]
- Paynter, S.; Russell, D.A. Surface plasmon resonance measurement of pH-induced responses of immobilized biomolecules: conformational change or electrostatic interaction effects? Anal. Biochem. 2002, 309, 85–95. [Google Scholar] [CrossRef]
- Hernandez, K.; Fernandez-Lafuente, R. Control of protein immobilization: Coupling immobilization and site-directed mutagenesis to improve biocatalyst or biosensor performance. Enzyme Microb. Technol. 2011, 48, 107–122. [Google Scholar] [CrossRef]
- Le, T.T.; Wilde, C.P.; Grossman, N.; Cass, A.E.G. A simple method for controlled immobilization of proteins on modified SAMs. Phys. Chem. Chem. Phys. 2011, 13, 5271–5278. [Google Scholar] [CrossRef]
- Takakura, Y.; Oka, N.; Kajiwara, H.; Tsunashima, M.; Usami, S.; Tsukamoto, H.; Ishida, Y.; Yamamoto, T. Tamavidin, a versatile affinity tag for protein purification and immobilization. J. Biotechnol. 2010, 145, 317–322. [Google Scholar] [CrossRef]
- Schlapschy, M.; Skerra, A. Purification and Analysis of Strep-tagged Antibody-Fragments. Antibody Engineering 2010, 317–330. [Google Scholar] [CrossRef]
- Lichty, J.J.; Malecki, J.L.; Agnew, H.D.; Michelson-Horowitz, D.J.; Tan, S. Comparison of affinity tags for protein purification. Protein Expres. Purif. 2005, 41, 98–105. [Google Scholar]
- Chapman-Smith, A.; Mulhern, T.D.; Whelan, F.; Cronan, J.E.; Wallace, J.C. The C-terminal domain of biotin protein ligase from E. coli is required for catalytic activity. Protein Sci. 2001, 10, 2608–2617. [Google Scholar] [CrossRef]
- Hochuli, E.; Bannwarth, W.; Dobeli, H.; Gentz, R.; Stuber, D. Genetic Approach to Facilitate Purification of Recombinant Proteins with a Novel Metal Chelate Adsorbent. Nat. Biotech. 1988, 6, 1321–1325. [Google Scholar] [CrossRef]
- Butler, J.E.; Ni, L.; Nessler, R.; Joshi, K.S.; Suter, M.; Rosenberg, B.; Chang, J.; Brown, W.R.; Cantarero, L.A. The physical and functional behavior of capture antibodies adsorbed on polystyrene. J. Immunol. Method. 1992, 150, 77–90. [Google Scholar] [CrossRef]
- Ng, E.W.M.; Shima, D.T.; Calias, P.; Cunningham, E.T.; Guyer, D.R.; Adamis, A.P. Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease. Nat. Rev. Drug Discov. 2006, 5, 123–132. [Google Scholar] [CrossRef]
- Walsh, M.K.; Wang, X.; Weimer, B.C. Optimizing the immobilization of single-stranded DNA onto glass beads. J. Biochem. Biophys. Methods 2001, 47, 221–231. [Google Scholar] [CrossRef]
- Kumar, A.; Larsson, O.; Parodi, D.; Liang, Z. Silanized nucleic acids: a general platform for DNA immobilization. Nucl. Acid. Res. 2000, 28, e71. [Google Scholar] [CrossRef]
- Herne, T.M.; Tarlov, M.J. Characterization of DNA Probes Immobilized on Gold Surfaces. J. Am. Chem. Soc. 1997, 119, 8916–8920. [Google Scholar] [CrossRef]
- Stoughton, R.B. Applications of DNA Microarrays in biology. Ann. Rev. Biochem. 2005, 74, 53–82. [Google Scholar] [CrossRef]
- Gresham, D.; Dunham, M.J.; Botstein, D. Comparing whole genomes using DNA microarrays. Nat. Rev. Genet. 2008, 9, 291–302. [Google Scholar] [CrossRef]
- Smith, L.; Greenfield, A. DNA microarrays and development. Hum. Mol. Genet. 2003, 12, R1–R8. [Google Scholar] [CrossRef]
- Sassolas, A.; Leca-Bouvier, B.D.; Blum, L.J. DNA Biosensors and Microarrays. Chem. Rev. 2007, 108, 109–139. [Google Scholar] [CrossRef]
- Gold, L.; Ayers, D.; Bertino, J.; Bock, C.; Bock, A.; Brody, E.N.; Carter, J.; Dalby, A.B.; Eaton, B.E.; Fitzwater, T.; et al. Aptamer-Based Multiplexed Proteomic Technology for Biomarker Discovery. PLoS One 2010, 5, e15004. [Google Scholar] [CrossRef]
- Petach, H.; Ostroff, R.; Greef, C.; Husar, G. Processing of Photoaptamer Microarrays. Protein Arrays 2004, 264, 101–110. [Google Scholar]
- Collett, J.R.; Cho, E.J.; Ellington, A.D. Production and processing of aptamer microarrays. Methods 2005, 37, 4–15. [Google Scholar] [CrossRef]
- Collett, J.R.; Cho, E.J.; Lee, J.F.; Levy, M.; Hood, A.J.; Wan, C.; Ellington, A.D. Functional RNA microarrays for high-throughput screening of antiprotein aptamers. Anal. Biochem. 2005, 338, 113–123. [Google Scholar] [CrossRef]
- The European Bioinformatics Institute Homepage. Available online: http://www.ebi.ac.uk (accessed on 25 February 2014).
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucl. Acid. Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef]
- Quinn, D.M. Acetylcholinesterase: Enzyme structure, reaction dynamics, and virtual transition states. Chem. Rev. 1987, 87, 955–979. [Google Scholar] [CrossRef]
- Nolte, H.J.; Rosenberry, T.L.; Neumann, E. Effective charge on acetylcholinesterase active sites determined from the ionic strength dependence of association rate constants with cationic ligands. Biochemistry 1980, 19, 3705–3711. [Google Scholar] [CrossRef]
- Bourne, Y.; Grassi, J.; Bougis, P.E.; Marchot, P. Conformational Flexibility of the Acetylcholinesterase Tetramer Suggested by X-ray Crystallography. J. Biol. Chem. 1999, 274, 30370–30376. [Google Scholar] [CrossRef]
- Vieregg, J.; Cheng, W.; Bustamante, C.; Tinoco, I. Measurement of the Effect of Monovalent Cations on RNA Hairpin Stability. J. Am. Chem. Soc. 2007, 129, 14966–14973. [Google Scholar] [CrossRef]
- Shiman, R.; Draper, D.E. Stabilization of RNA tertiary structure by monovalent cations. J. Mol. Biol. 2000, 302, 79–91. [Google Scholar] [CrossRef]
- Gunaratna, P.C.; Wilson, G.S. Use of monoclonal anti-enzyme antibodies for analytical purposes. Biotechnol. Progr. 1992, 8, 268–274. [Google Scholar] [CrossRef]
- Drolet, D.W.; Moon-McDermott, L.; Romig, T.S. An enzyme-linked oligonucleotide assay. Nat. Biotech. 1996, 14, 1021–1025. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chumphukam, O.; Le, T.T.; Cass, A.E.G. High Efficiency Acetylcholinesterase Immobilization on DNA Aptamer Modified Surfaces. Molecules 2014, 19, 4986-4996. https://doi.org/10.3390/molecules19044986
Chumphukam O, Le TT, Cass AEG. High Efficiency Acetylcholinesterase Immobilization on DNA Aptamer Modified Surfaces. Molecules. 2014; 19(4):4986-4996. https://doi.org/10.3390/molecules19044986
Chicago/Turabian StyleChumphukam, Orada, Thao T. Le, and Anthony E. G. Cass. 2014. "High Efficiency Acetylcholinesterase Immobilization on DNA Aptamer Modified Surfaces" Molecules 19, no. 4: 4986-4996. https://doi.org/10.3390/molecules19044986
APA StyleChumphukam, O., Le, T. T., & Cass, A. E. G. (2014). High Efficiency Acetylcholinesterase Immobilization on DNA Aptamer Modified Surfaces. Molecules, 19(4), 4986-4996. https://doi.org/10.3390/molecules19044986