
The Giant Adhesin SiiE of Salmonella enterica

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Salmonella Enterica Pathogenicity
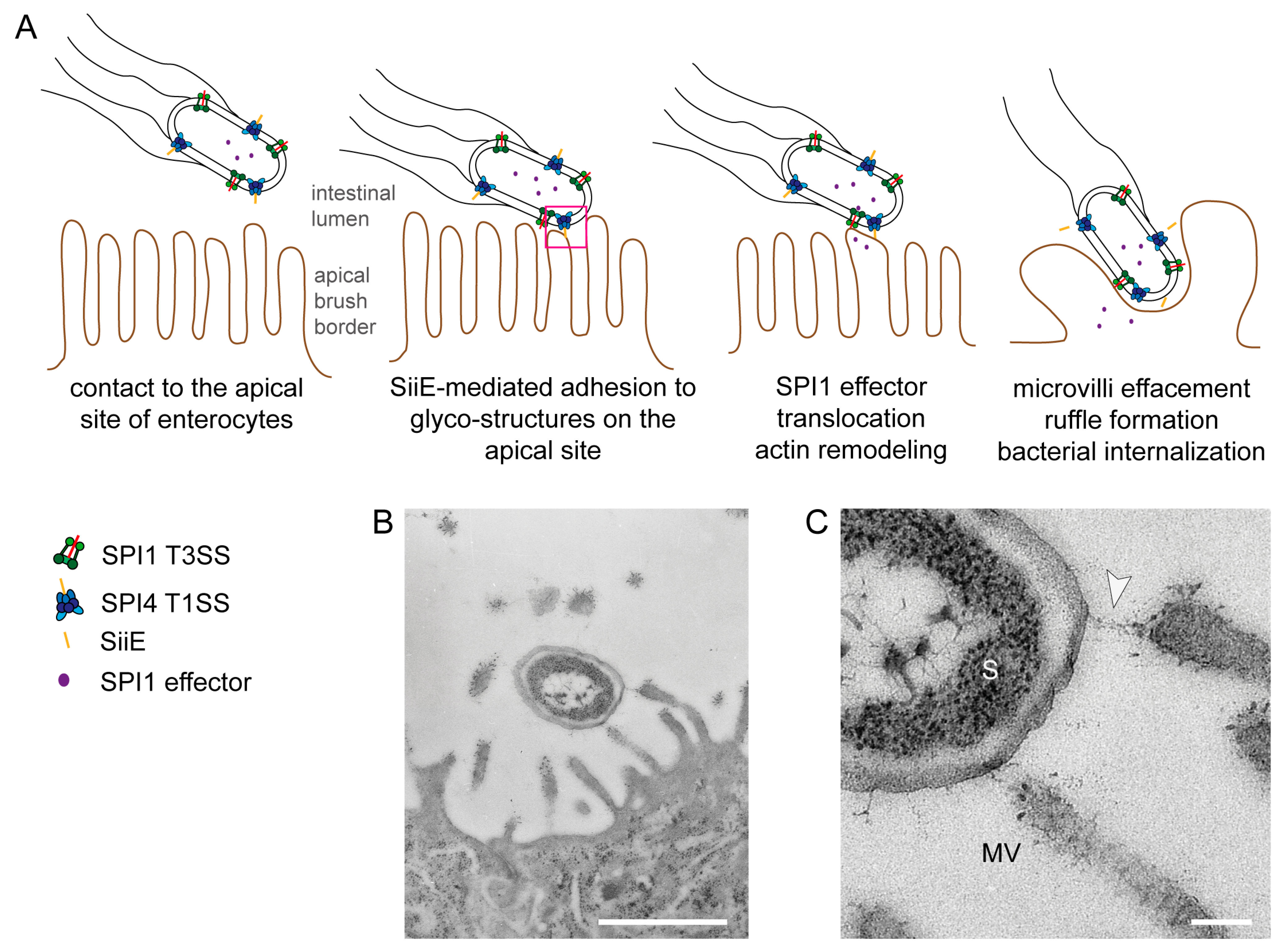
2. SiiE—A Non-Fimbrial Adhesin of Salmonella enterica
2.1. Role of Non-Fimbrial Adhesins in Salmonella Pathobiology
2.2. Non-Fimbrial Adhesins

2.3. The BIg Domain in Bacterial Proteins
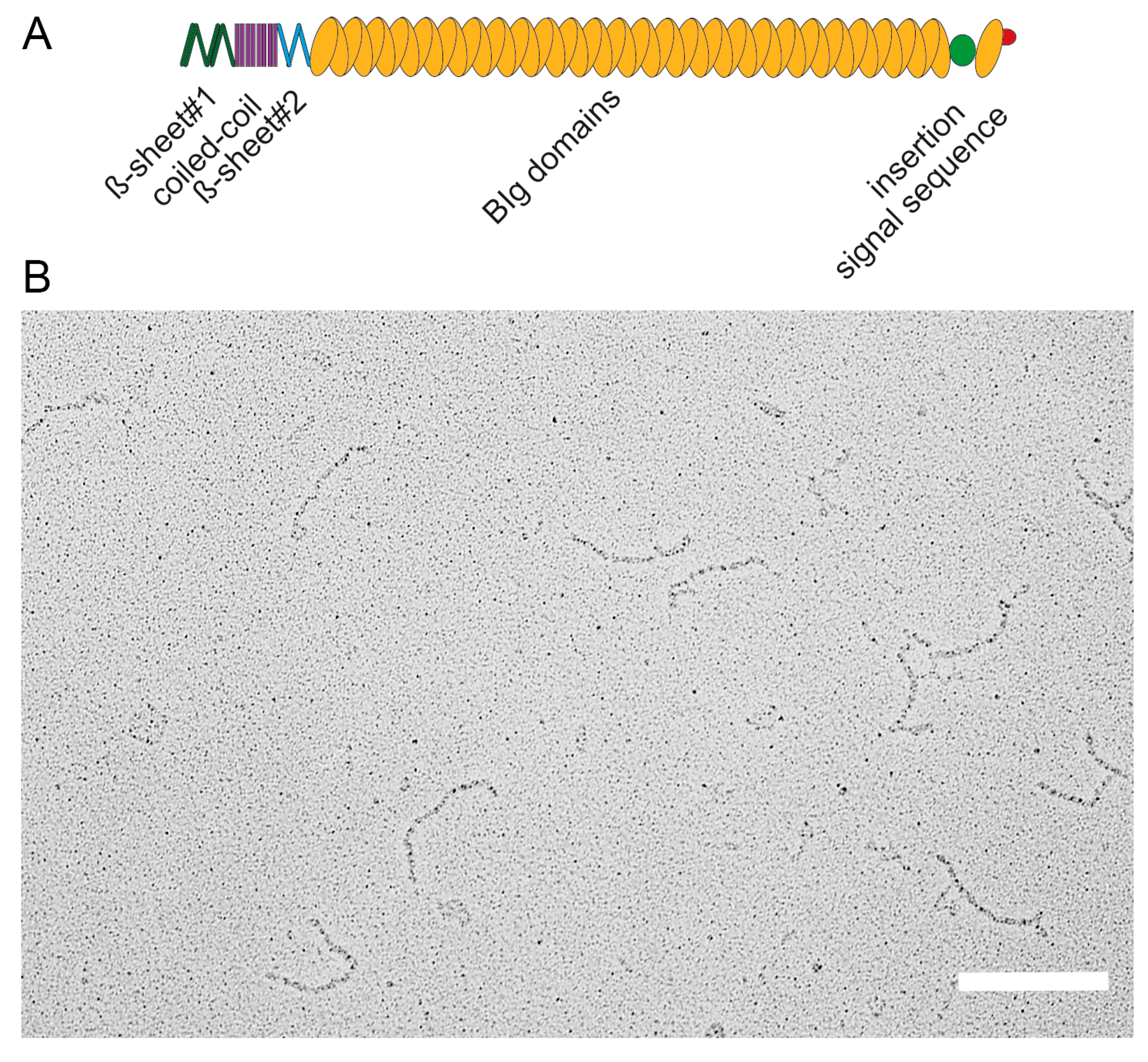
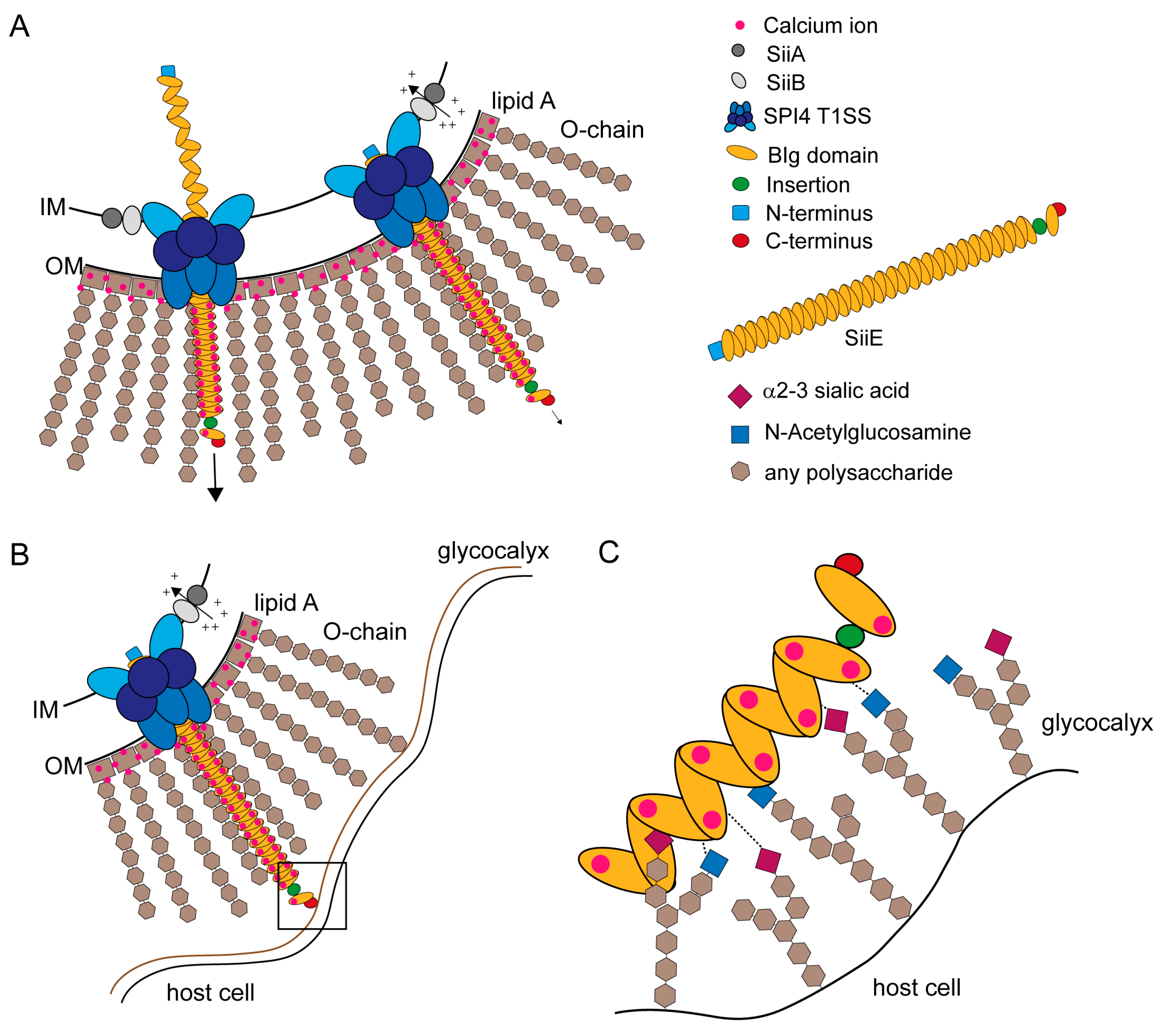
2.4. The Giant Adhesin SiiE

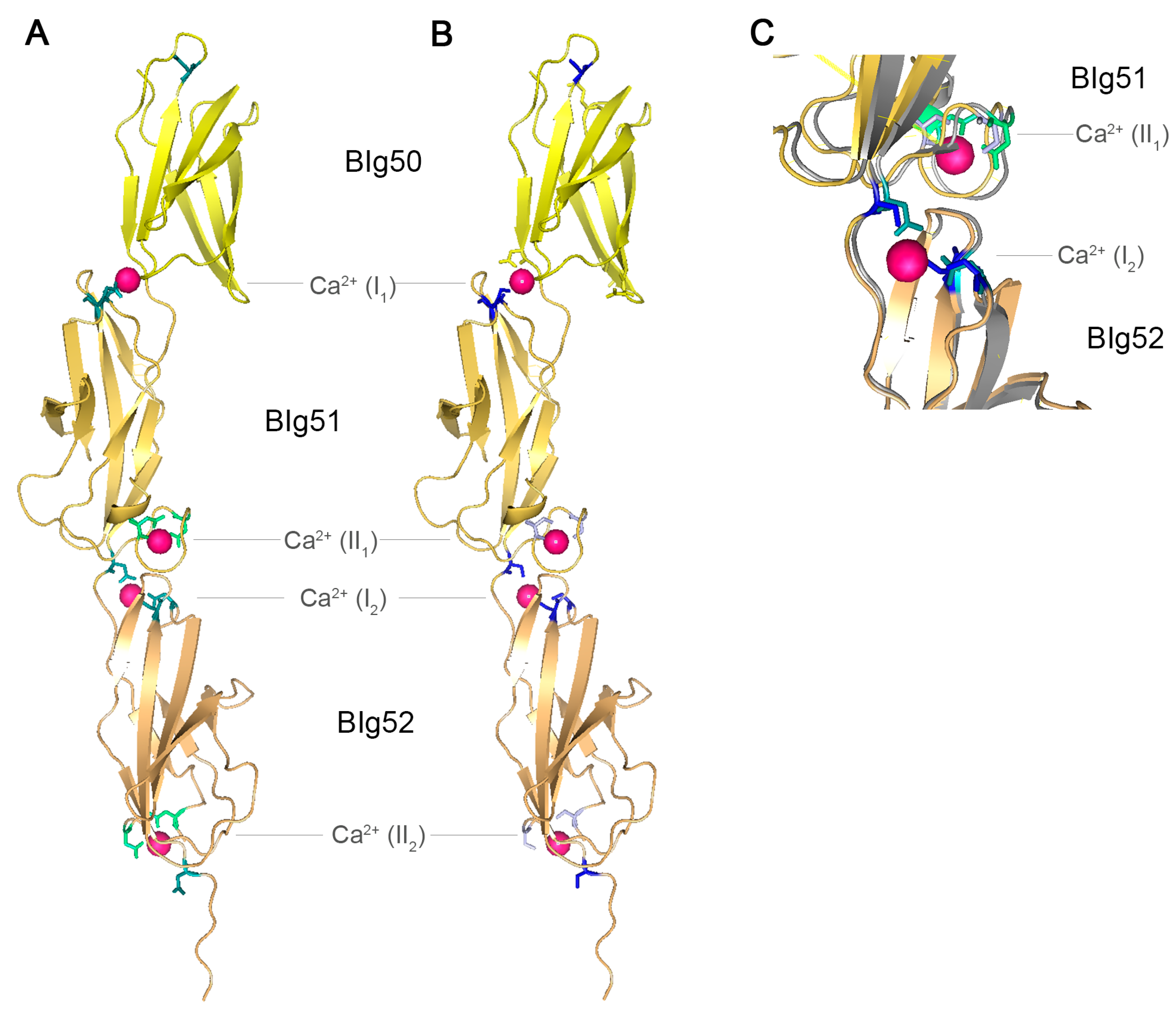
2.5. BIg Domains in SiiE


2.6. SiiE Binds in a Lectin-Like Manner
3. Conclusions and Outlook

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Marcus, S.L.; Brumell, J.H.; Pfeifer, C.G.; Finlay, B.B. Salmonella pathogenicity islands: Big virulence in small packages. Microbes Infect. 2000, 2, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Hensel, M. Evolution of pathogenicity islands of Salmonella enterica. Int. J. Med. Microbiol. 2004, 294, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Chakravortty, D.; Hansen-Wester, I.; Hensel, M. Salmonella pathogenicity island 2 mediates protection of intracellular Salmonella from reactive nitrogen intermediates. J. Exp. Med. 2002, 195, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, R.G.; Claudio, N.; Rohde, M.; Jackel, D.; Wagner, C.; Hensel, M. Cooperation of Salmonella pathogenicity islands 1 and 4 is required to breach epithelial barriers. Cell Microbiol. 2008, 10, 2364–2376. [Google Scholar] [CrossRef] [PubMed]
- Ahmer, B.M.; van Reeuwijk, J.; Watson, P.R.; Wallis, T.S.; Heffron, F. Salmonella SirA is a global regulator of genes mediating enteropathogenesis. Mol. Microbiol. 1999, 31, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, R.G.; Jackel, D.; Geymeier, N.; Hensel, M. Salmonella pathogenicity island 4-mediated adhesion is coregulated with invasion genes in Salmonella enterica. Infect. Immun. 2007, 75, 4697–4709. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Polke, M.; Gerlach, R.G.; Linke, D.; Stierhof, Y.D.; Schwarz, H.; Hensel, M. Functional dissection of SiiE, a giant non-fimbrial adhesin of Salmonella enterica. Cell. Microbiol. 2011, 13, 1286–1301. [Google Scholar] [CrossRef] [PubMed]
- Griessl, M.H.; Schmid, B.; Kassler, K.; Braunsmann, C.; Ritter, R.; Barlag, B.; Stierhof, Y.D.; Sturm, K.U.; Danzer, C.; Wagner, C.; et al. Structural insight into the giant Ca2+-binding adhesin SiiE: Implications for the adhesion of Salmonella enterica to polarized epithelial cells. Structure 2013, 21, 741–752. [Google Scholar]
- Wagner, C.; Hensel, M. Adhesive mechanisms of Salmonella enterica. Adv. Exp. Med. Biol. 2011, 715, 17–34. [Google Scholar] [PubMed]
- Latasa, C.; Roux, A.; Toledo-Arana, A.; Ghigo, J.M.; Gamazo, C.; Penades, J.R.; Lasa, I. BapA, a large secreted protein required for biofilm formation and host colonization of Salmonella enterica serovar Enteritidis. Mol. Microbiol. 2005, 58, 1322–1339. [Google Scholar] [CrossRef] [PubMed]
- Blanc-Potard, A.B.; Solomon, F.; Kayser, J.; Groisman, E.A. The SPI-3 pathogenicity island of Salmonella enterica. J. Bacteriol. 1999, 181, 998–1004. [Google Scholar] [PubMed]
- Kingsley, R.A.; van Amsterdam, K.; Kramer, N.; Baumler, A.J. The shdA gene is restricted to serotypes of Salmonella enterica subspecies I and contributes to efficient and prolonged fecal shedding. Infect. Immun. 2000, 68, 2720–2727. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, C.W.; Laarakker, M.C.; Humphries, A.D.; Weening, E.H.; Baumler, A.J. Salmonella enterica serotype Typhimurium MisL is an intestinal colonization factor that binds fibronectin. Mol. Microbiol. 2005, 57, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, R.A.; Santos, R.L.; Keestra, A.M.; Adams, L.G.; Baumler, A.J. Salmonella enterica serotype Typhimurium ShdA is an outer membrane fibronectin-binding protein that is expressed in the intestine. Mol. Microbiol. 2002, 43, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, R.G.; Jäckel, D.; Stecher, B.; Wagner, C.; Lupas, A.; Hardt, W.D.; Hensel, M. Salmonella Pathogenicity Island 4 encodes a giant non-fimbrial adhesin and the cognate type 1 secretion system. Cell. Microbiol. 2007, 9, 1834–1850. [Google Scholar] [CrossRef] [PubMed]
- Main-Hester, K.L.; Colpitts, K.M.; Thomas, G.A.; Fang, F.C.; Libby, S.J. Coordinate regulation of Salmonella pathogenicity island 1 (SPI1) and SPI4 in Salmonella enterica serovar Typhimurium. Infect. Immun. 2008, 76, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.; Campbell, J.D.; Rowe, S.C.; Bispham, J.; Stevens, M.P.; Bowen, A.J.; Barrow, P.A.; Maskell, D.J.; Wallis, T.S. Identification of host-specific colonization factors of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2004, 54, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Artsimovitch, I.; Landick, R. The transcriptional regulator RfaH stimulates RNA chain synthesis after recruitment to elongation complexes by the exposed nontemplate DNA strand. Cell 2002, 109, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Soto, G.E.; Hultgren, S.J. Bacterial adhesins: Common themes and variations in architecture and assembly. J. Bacteriol. 1999, 181, 1059–1071. [Google Scholar] [PubMed]
- Bouckaert, J.; Berglund, J.; Schembri, M.; de Genst, E.; Cools, L.; Wuhrer, M.; Hung, C.S.; Pinkner, J.; Slattegard, R.; Zavialov, A.; et al. Receptor binding studies disclose a novel class of high-affinity inhibitors of the Escherichia coli FimH adhesin. Mol. Microbiol. 2005, 55, 441–455. [Google Scholar]
- Lund, B.; Lindberg, F.; Marklund, B.I.; Normark, S. The PapG protein is the α-d-galactopyranosyl-(1->4)-β-d-galactopyranose-binding adhesin of uropathogenic Escherichia coli. Proc. Natl. Acad. Sci. USA 1987, 84, 5898–5902. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, R.G.; Hensel, M. Protein secretion systems and adhesins: The molecular armory of Gram-negative pathogens. Int. J. Med. Microbiol. 2007, 297, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Linke, D.; Riess, T.; Autenrieth, I.B.; Lupas, A.; Kempf, V.A. Trimeric autotransporter adhesins: Variable structure, common function. Trends Microbiol. 2006, 14, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Nummelin, H.; Merckel, M.C.; Leo, J.C.; Lankinen, H.; Skurnik, M.; Goldman, A. The Yersinia adhesin YadA collagen-binding domain structure is a novel left-handed parallel beta-roll. EMBO J. 2004, 23, 701–711. [Google Scholar] [CrossRef] [PubMed]
- Riess, T.; Andersson, S.G.; Lupas, A.; Schaller, M.; Schafer, A.; Kyme, P.; Martin, J.; Walzlein, J.H.; Ehehalt, U.; Lindroos, H.; et al. Bartonella adhesin a mediates a proangiogenic host cell response. J. Exp. Med. 2004, 200, 1267–1278. [Google Scholar]
- Delepelaire, P. Type I secretion in gram-negative bacteria. Biochim. Biophys. Acta 2004, 1694, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O’Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003, 49, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Walz, A.; Odenbreit, S.; Stuhler, K.; Wattenberg, A.; Meyer, H.E.; Mahdavi, J.; Boren, T.; Ruhl, S. Identification of glycoprotein receptors within the human salivary proteome for the lectin-like BabA and SabA adhesins of Helicobacter pylori by fluorescence-based 2-D bacterial overlay. Proteomics 2009, 9, 1582–1592. [Google Scholar] [CrossRef] [PubMed]
- Unemo, M.; Aspholm-Hurtig, M.; Ilver, D.; Bergstrom, J.; Boren, T.; Danielsson, D.; Teneberg, S. The sialic acid binding SabA adhesin of Helicobacter pylori is essential for nonopsonic activation of human neutrophils. J. Biol. Chem. 2005, 280, 15390–15397. [Google Scholar] [CrossRef] [PubMed]
- Parreira, P.; Shi, Q.; Magalhaes, A.; Reis, C.A.; Bugaytsova, J.; Boren, T.; Leckband, D.; Martins, M.C. Atomic force microscopy measurements reveal multiple bonds between Helicobacter pylori blood group antigen binding adhesin and Lewis b ligand. J. R. Soc. Interface 2014, 11. [Google Scholar] [CrossRef]
- Halaby, D.M.; Mornon, J.P. The immunoglobulin superfamily: An insight on its tissular, species, and functional diversity. J. Mol. Evol. 1998, 46, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.F.; Barclay, A.N. The immunoglobulin superfamily—Domains for cell surface recognition. Annu. Rev. Immunol. 1988, 6, 381–405. [Google Scholar] [CrossRef] [PubMed]
- Halaby, D.M.; Poupon, A.; Mornon, J. The immunoglobulin fold family: Sequence analysis and 3D structure comparisons. Protein Eng. 1999, 12, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Lasa, I.; Penades, J.R. Bap: A family of surface proteins involved in biofilm formation. Res. Microbiol. 2006, 157, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, Z.A.; Brown, M.S.; Isberg, R.R.; Bjorkman, P.J. Crystal structure of invasin: A bacterial integrin-binding protein. Science 1999, 286, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Dersch, P.; Isberg, R.R. An immunoglobulin superfamily-like domain unique to the Yersinia pseudotuberculosis invasin protein is required for stimulation of bacterial uptake via integrin receptors. Infect. Immun. 2000, 68, 2930–2938. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, D.A.; Tailford, L.E.; Hemmings, A.M.; Juge, N. Crystal structure of a mucus-binding protein repeat reveals an unexpected functional immunoglobulin binding activity. J. Biol. Chem. 2009, 284, 32444–32453. [Google Scholar] [CrossRef] [PubMed]
- Etzold, S.; Kober, O.I.; Mackenzie, D.A.; Tailford, L.E.; Gunning, A.P.; Walshaw, J.; Hemmings, A.M.; Juge, N. Structural basis for adaptation of lactobacilli to gastrointestinal mucus. Environ. Microbiol. 2014, 16, 888–903. [Google Scholar] [CrossRef] [PubMed]
- Raman, R.; Rajanikanth, V.; Palaniappan, R.U.; Lin, Y.P.; He, H.; McDonough, S.P.; Sharma, Y.; Chang, Y.F. Big domains are novel Ca2+-binding modules: Evidences from big domains of Leptospira immunoglobulin-like (Lig) proteins. PLoS One 2010, 5, e14377. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Paterson, N.G.; Gaspar, A.H.; Ton-That, H.; Baker, E.N. The Corynebacterium diphtheriae shaft pilin SpaA is built of tandem Ig-like modules with stabilizing isopeptide and disulfide bonds. Proc. Natl. Acad. Sci. USA 2009, 106, 16967–16971. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Bakkes, P.J.; Smits, S.H.; Schmitt, L. Equilibrium folding of pro-HlyA from Escherichia coli reveals a stable calcium ion dependent folding intermediate. Biochim. Biophys. Acta 2014, 1844, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Frey, E.A.; Pfuetzner, R.A.; Creagh, A.L.; Knoechel, D.G.; Haynes, C.A.; Finlay, B.B.; Strynadka, N.C. Crystal structure of enteropathogenic Escherichia coli intimin-receptor complex. Nature 2000, 405, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Barlag, B.; Gerlach, R.G.; Deiwick, J.; Hensel, M. The Salmonella enterica giant adhesin SiiE binds to polarized epithelial cells in a lectin-like manner. Cell. Microbiol. 2014, 16, 962–975. [Google Scholar] [CrossRef] [PubMed]
- Wille, T.; Wagner, C.; Mittelstadt, W.; Blank, K.; Sommer, E.; Malengo, G.; Dohler, D.; Lange, A.; Sourjik, V.; Hensel, M.; et al. SiiA and SiiB are novel type I secretion system subunits controlling SPI4-mediated adhesion of Salmonella enterica. Cell. Microbiol. 2014, 16, 161–178. [Google Scholar]
- Parkhill, J.; Dougan, G.; James, K.D.; Thomson, N.R.; Pickard, D.; Wain, J.; Churcher, C.; Mungall, K.L.; Bentley, S.D.; Holden, M.T.; et al. Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature 2001, 413, 848–852. [Google Scholar]
- Newell, P.D.; Yoshioka, S.; Hvorecny, K.L.; Monds, R.D.; O’Toole, G.A. Systematic analysis of diguanylate cyclases that promote biofilm formation by Pseudomonas fluorescens Pf0–1. J. Bacteriol. 2011, 193, 4685–4698. [Google Scholar] [CrossRef] [PubMed]
- Kisiela, D.I.; Chattopadhyay, S.; Libby, S.J.; Karlinsey, J.E.; Fang, F.C.; Tchesnokova, V.; Kramer, J.J.; Beskhlebnaya, V.; Samadpour, M.; Grzymajlo, K.; et al. Evolution of Salmonella enterica virulence via point mutations in the fimbrial adhesin. PLoS Pathog. 2012, 8, e1002733. [Google Scholar]
- Chessa, D.; Dorsey, C.W.; Winter, M.; Baumler, A.J. Binding specificity of Salmonella plasmid-encoded fimbriae assessed by glycomics. J. Biol. Chem. 2008, 283, 8118–8124. [Google Scholar] [CrossRef] [PubMed]
- Chessa, D.; Winter, M.G.; Jakomin, M.; Baumler, A.J. Salmonella enterica serotype Typhimurium Std fimbriae bind terminal α(1,2)fucose residues in the cecal mucosa. Mol. Microbiol. 2009, 71, 864–875. [Google Scholar] [CrossRef] [PubMed]
- Ishijima, N.; Suzuki, M.; Ashida, H.; Ichikawa, Y.; Kanegae, Y.; Saito, I.; Boren, T.; Haas, R.; Sasakawa, C.; Mimuro, H. BabA-mediated adherence is a potentiator of the Helicobacter pylori type IV secretion system activity. J. Biol. Chem. 2011, 286, 25256–25264. [Google Scholar] [CrossRef] [PubMed]
- Ilver, D.; Arnqvist, A.; Ogren, J.; Frick, I.M.; Kersulyte, D.; Incecik, E.T.; Berg, D.E.; Covacci, A.; Engstrand, L.; Boren, T. Helicobacter pylori adhesin binding fucosylated histo-blood group antigens revealed by retagging. Science 1998, 279, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi, J.; Sonden, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.A.; et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar]
- Aspholm, M.; Kalia, A.; Ruhl, S.; Schedin, S.; Arnqvist, A.; Linden, S.; Sjostrom, R.; Gerhard, M.; Semino-Mora, C.; Dubois, A.; et al. Helicobacter pylori adhesion to carbohydrates. Methods Enzymol. 2006, 417, 293–339. [Google Scholar]
- Aspholm, M.; Olfat, F.O.; Norden, J.; Sonden, B.; Lundberg, C.; Sjostrom, R.; Altraja, S.; Odenbreit, S.; Haas, R.; Wadstrom, T.; et al. SabA is the H. pylori hemagglutinin and is polymorphic in binding to sialylated glycans. PLoS Pathog. 2006, 2, e110. [Google Scholar]
- Comolli, J.C.; Waite, L.L.; Mostov, K.E.; Engel, J.N. Pili binding to asialo-GM1 on epithelial cells can mediate cytotoxicity or bacterial internalization by Pseudomonas aeruginosa. Infect. Immun. 1999, 67, 3207–3214. [Google Scholar] [PubMed]
- Ulloa, F.; Real, F.X. Differential distribution of sialic acid in α2,3 and α2,6 linkages in the apical membrane of cultured epithelial cells and tissues. J. Histochem. Cytochem. 2001, 49, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Pieters, R.J. Carbohydrate mediated bacterial adhesion. Adv. Exp. Med. Biol. 2011, 715, 227–240. [Google Scholar] [PubMed]
- De Greve, H.; Wyns, L.; Bouckaert, J. Combining sites of bacterial fimbriae. Curr. Opin. Struct. Biol. 2007, 17, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Dutzler, R.; Wang, Y.F.; Rizkallah, P.; Rosenbusch, J.P.; Schirmer, T. Crystal structures of various maltooligosaccharides bound to maltoporin reveal a specific sugar translocation pathway. Structure 1996, 4, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.; Bowen, A.J.; Carnell, S.C.; Wallis, T.S.; Stevens, M.P. SiiE is secreted by the Salmonella enterica serovar Typhimurium pathogenicity island 4-encoded secretion system and contributes to intestinal colonization in cattle. Infect. Immun. 2007, 75, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Morgan, E.; Nagy, G. Contribution of SPI-4 genes to the virulence of Salmonella enterica. FEMS Microbiol. Lett. 2007, 275, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Sturm, K.U.; Griessl, M.H.; Wagner, C.; Deiwick, J.; Hensel, M.; Muller, Y.A. Crystallization and preliminary crystallographic analysis of an Ig-domain-encompassing fragment of the giant adhesion protein SiiE from Salmonella enterica. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 1371–1374. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barlag, B.; Hensel, M. The Giant Adhesin SiiE of Salmonella enterica. Molecules 2015, 20, 1134-1150. https://doi.org/10.3390/molecules20011134
Barlag B, Hensel M. The Giant Adhesin SiiE of Salmonella enterica. Molecules. 2015; 20(1):1134-1150. https://doi.org/10.3390/molecules20011134
Chicago/Turabian StyleBarlag, Britta, and Michael Hensel. 2015. "The Giant Adhesin SiiE of Salmonella enterica" Molecules 20, no. 1: 1134-1150. https://doi.org/10.3390/molecules20011134
APA StyleBarlag, B., & Hensel, M. (2015). The Giant Adhesin SiiE of Salmonella enterica. Molecules, 20(1), 1134-1150. https://doi.org/10.3390/molecules20011134