
Molecular Progress in Research on Fruit Astringency

{kind=link}
Abstract
:1. Introduction
2. Tannins and the Generation of Fruit Astringency
2.1. Astringency and the Antioxidative Activity of Tannins
2.2. The Molecular Structure and Polymerization of Tannins and Their Relationship with the Intensity of Astringency in Fruit
2.3. The Relationship between Polysaccharides, Acids and the Intensity of Astringency in Fruit
2.4. The Localization of Tannins in Fruit
3. The Mechanism of the Biosynthesis of Astringent Substances
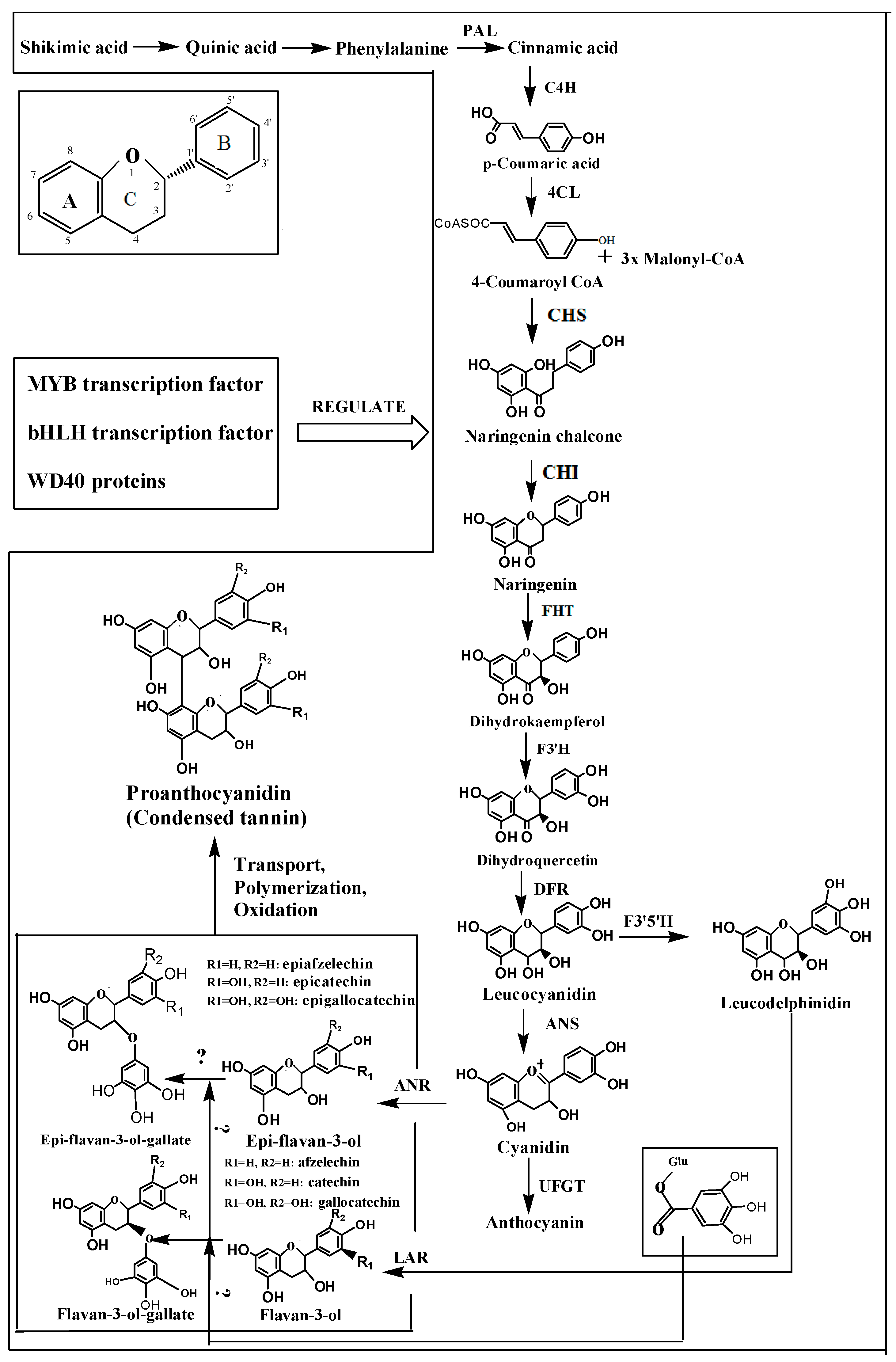
3.1. Phenylalanine Ammonia-Lyase
3.2. Cinnamate 4-Hydroxylase
3.3. Flavanone Hydroxylase
3.4. Dihydroflavonol Reductase
3.5. Leucoanthocyanidin Reductase
3.6. Anthocyanin Synthase
3.7. Anthocyanin Reductase
3.8. Function of the Main Enzymes are Described in Detail
4. Regulation of the Content of Astringent Substances in Fruit
5. Prospects for Studying Astringency of Fruit
Acknowledgments
Author Contributions
Conflicts of Interest
References
- EI Hadi, M.A.M.; Zhang, F.J.; Wu, F.F.; Zhou, C.H.; Tao, J. Advances in fruit aroma volatile research. Molecules 2013, 18, 8200–8229. [Google Scholar] [CrossRef] [PubMed]
- Gawel, R.; Oberholster, A.; Francis, I.L. A “Mouth-feel Wheel”: Terminology for communicating the mouth-feel characteristics of red wine. Aust. J. Grape Wine Res. 2000, 6, 203–207. [Google Scholar] [CrossRef]
- Bate-sminth, E.C. Flavonoid compounds in foods. Adv. Food Res. 1954, 5, 261–300. [Google Scholar] [PubMed]
- Kennedy, J.A.; Saucier, C.; Glories, Y. Grape and wine phenolics: History and perspective. Am. J. Enol. Vitic. 2006, 57, 239–248. [Google Scholar]
- Brossaud, F.; Cheynier, V.; Noble, A.C. Bitterness and astringency of grape and wine polyphenols. Aust. J. Grape Wine Res. 2001, 7, 33–39. [Google Scholar] [CrossRef]
- Preys, S.; Mazerolles, G.; Courcoux, P.; Samson, A.; Fischer, U.; Hanafi, M.; Bertrand, D.; Cheynier, V. Relationship between polyphenolic composition and some sensory properties in red wines using multiway analyses. Anal. Chim. Acta 2006, 563, 126–136. [Google Scholar] [CrossRef]
- Guinoza Siraichi, J.T.; Felipe, D.F.; Serra Brambilla, L.Z.; Gatto, M.J.; Terra, V.A.; Cecchini, A.L.; Cortez, L.E.; Rodrigues-Filho, E.; Cortez, D.A. Antioxidant Capacity of the Leaf Extract Obtained from Arrabidaea chica Cultivated in Southern Brazil. PLoS One 2013, 8, e72733. [Google Scholar] [CrossRef] [PubMed]
- Besada, C.; Salvador, A.; Arnal, L. Martínez-Jávega, J.M. Optimization of the duration of deastringency treatment depending on persimmon maturity. Acta Hortic. 2010, 858, 69–74. [Google Scholar]
- Demiglio, P.; Pickering, G.J. The influence of ethanol and pH on the taste and mouthfeel sensations elicited by red wine. J. Food Agric. Environ. 2008, 6, 143–150. [Google Scholar]
- Dinnella, C.; Recchia, A.; Fia, G.; Bertuccioli, M.; Monteleone, E. Saliva characteristics and individual sensitivity to phenolic astringent stimuli. Chem. Senses 2009, 34, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Labieniec, M.; Gabryelak, T. Interactions of tannic acid and its derivatives (ellagic and gallic acid) with calf thymus DNA and bovine serum albumin using spectroscopic method. J. Photochem. Photobiol. B 2006, 82, 72–78. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Shi, B.; Yao, K. Interactions of gallotannins with proteins, amino acids, phospholipids and sugars. Food Chem. 2006, 95, 250–254. [Google Scholar] [CrossRef]
- Wang, B.H.; Foo, L.Y.; Polya, G.M. Differential inhibition of eukaryote protein kinases by condensed tannins. Phytochemistry 1996, 43, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.Y.; Wang, C.P.; Huang, S.S.; Hsu, F.L. The in-hibitory effect of tannins of lipid peroxidation of rat heart mitochondria. J. Pharm. Pharmacol. 1995, 47, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Gust, J.; Suwalski, J. Relationship between radical scavenging effects and anticorrosive properties of polyphe-nols. Corrosion 1995, 51, 37–44. [Google Scholar] [CrossRef]
- Salah, N.; Miller, N.J.; Paganga, G.; Tijburg, L.; Bolwell, G.P.; Rice-Evans, C. Polyphenolic flavanols as scavengers of aqueous phase radicals and as chain-breaking antioxi-dants. Arch. Biochem. Biophys 1995, 322, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Kimura, Y.; Yoshida, T.; Hatano, T.; Okuda, H.; Arichi, S. Studies on the activities of tannins and related compounds from medicinal plants and drugs. I. Inhibitory effects on lipid peroxidation in mitochondria and microsomes of liver. Chem. Pharm. Bull. 1983, 31, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Calixto, B.J.; Nicolau, N.; Rae, G.A. Pharmacological actions of tannic acid I. effects on isolated smooth and cardiac muscles and on blood pressure. Planta Med. 1986, 52, 32–35. [Google Scholar] [CrossRef]
- Hanlin, R.L.; Hrmova, M.; Harbertson, J.F.; Downey, M.O. Condensed tannin and grape cell wall interactions and their impact on tannin extractability into wine. Aust. J. Grape Wine Res. 2010, 16, 173–188. [Google Scholar] [CrossRef]
- Quijada-Morin, N.; Regueiro, J.; Simal-Gandara, J.; Tomas, E.; Rivas-Gonzalo, J.C.; Escribano-Bailon, M.T. Relationship between the sensory-determined astringency and the flavanolic composition of red wines. J. Agric. Food Chem. 2012, 60, 12355–12361. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Ferrier, J.; Harbertson, J.F.; Peyrot des Gachons, C. Analysis of tannins in red wine using multiple methods: Correlation with perceived astringency. Am. J. Enol. Vitic. 2006, 57, 481–485. [Google Scholar]
- Xie, D.; Dixon, R.A. Review—Proanthocyanidin biosynthesis—Still more questions than answers? Phytochemistry 2005, 66, 2127–2144. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O. Phenolics and ripening in grape berries. Am. J. Enol. Vitic. 2006, 57, 249–256. [Google Scholar]
- Arnold, R.A.; Noble, A.C.; Singleton, V.L. Bitterness and astringency of phenolic fractions in wine. J. Agric. Food Chem. 1980, 28, 675–678. [Google Scholar] [CrossRef]
- Peleg, H.; Gacon, K.; Schlich, P.; Noble, A.C. Bitterness and astringency of flavan-3-ol monomers, dimers and trimers. J. Sci. Food Agric. 1999, 79, 1123–1128. [Google Scholar] [CrossRef]
- Vidal, S.; Francis, L.; Guyot, S.; Marnet, N.; Kwiatkowski, M.; Gawel, R.; Véronique Cheynier, V.; Waters, E.L. The mouth-feel properties of grape and apple proanthocyanidins in a wine-like medium. J. Sci. Food Agric. 2003, 83, 564–573. [Google Scholar] [CrossRef]
- Symoneaux, R.; Baron, A.; Marnet, N.; Bauduin, R.; Chollet, S. Impact of apple procyanidins on sensory perception in model cider (part 1): Polymerisation Degree and concentration. Food Sci. Technol. 2014, 57, 22–27. [Google Scholar]
- Rustioni, L.; Fiori, S.; Failla, O. Evaluation of tannins interactions in grape (Vitis vinifera L.) skins. Food Chem. 2014, 159, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.; Povoas, M.J.; Mateus, N.; de Freitas, V. Application of flow nephelometry to the analysis of the influence of carbohydrates on protein–tannin interactions. J. Sci. Food Agric. 2006, 86, 891–896. [Google Scholar] [CrossRef]
- Escot, S.; Feuillat, M.; Dulau, L.; Charpentier, C. Release of polysaccharides by yeasts and the influence of released polysaccharides on colour stability and wine astringency. Aust. J. Grape Wine Res. 2001, 7, 153–159. [Google Scholar] [CrossRef]
- De Freitas, V.; Carvalho, E.; Mateus, N. Study of carbohydrate influence on protein–tannin aggregation by nephelometry. Food Chem. 2003, 81, 503–509. [Google Scholar] [CrossRef]
- Mateus, N.; Carvalho, E.; Luis, C.; de Freitas, V. Influence of the tannin structure on the disruption effect of carbohydrates on protein–tannin aggregates. Anal. Chim. Acta 2004, 513, 135–140. [Google Scholar] [CrossRef]
- Poncet-Legrand, C.; Doco, T.; Williams, P.; Vernhet, A. Inhibition of grape seed tannin aggregation by wine mannoproteins: Effect of polysaccharide molecular weight. Am. J. Enol. Vitic. 2007, 58, 87–91. [Google Scholar]
- Riou, V.; Vernhet, A.; Doco, T.; Moutounet, M. Aggregation of grape seed tannins in model wine–effect of wine polysaccharides. Food Hydrocolloids 2002, 16, 17–23. [Google Scholar] [CrossRef]
- Vernhet, A.; Pellerin, P.; Prieur, C.; Osmianski, J.; Moutounet, M. Charge properties of some grape and wine polysaccharide and polyphenolic fractions. Am. J. Enol. Vitic. 1996, 47, 25–30. [Google Scholar]
- Vidal, S.; Francis, L.; Williams, P.; Kwiatkowski, M.; Gawel, R.; Cheynier, V.; Waters, E. The mouth-feel properties of polysaccharides and anthocyanins in a wine like medium. Food Chem. 2004, 85, 519–525. [Google Scholar] [CrossRef]
- Quijada-Morín, N.; Williams, P.; Rivas-Gonzalo, J.C.; Doco, T.; Escribano-Bailón, M.T. Polyphenolic, polysaccharide and oligosaccharide composition of Tempranillo red wines and their relationship with the perceived astringency. Food Chem. 2014, 154, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Renard, C.M.G.C.; Baron, A.; Guyot, S.; Drilleau, J.F. Interactions between apple cell walls and native apple polyphenols: Quantification and some consequences. Int. J. Biol. Macromol. 2001, 29, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Hayasaka, Y.; Vidal, S.; Waters, E.J.; Jones, G.P. Composition of grape skin proanthocyanidins at different stages of berry development. J. Agric. Food Chem. 2001, 49, 5348–5355. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.; Boulton, R.B.; Noble, A.C. Physiological factors contributing to the variability of sensory assessments: Relationship between salivary flow rate and temporal perception of gustatory stimuli. Food Qual. Prefer. 1994, 5, 55–64. [Google Scholar] [CrossRef]
- Kallithraka, S.; Bakker, J.; Clifford, M.N. Red wine and model wine astringency as affected by malic and lactic acid. J. Food Sci. 1997, 62, 416–420. [Google Scholar] [CrossRef]
- Guinard, J.-X.; Pangborn, R.M.; Lewis, M.J. Preliminary studies on acidity-astringency interactions in model solutions and wines. J. Sci. Food Agric. 1986, 37, 811–817. [Google Scholar] [CrossRef]
- Hanna, P.; Keith, K.B.; Ann, C.N. The Influence of Acid on Astringency of Alum and Phenolic Compounds. Chem. Senses 1998, 23, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Peleg, H.; Nobleb, A.C. Effect of viscosity, temperature and pH on astringency in cranberry juice. Food Qual. Prefer. 1999, 10, 343–347. [Google Scholar] [CrossRef]
- Beecher, J.W.; Drake, M.A.; Luck, P.J.; Foegeding, E.A. Factors Regulating Astringency of Whey Protein Beverages. J. Dairy Sci. 2008, 91, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- Vardhanabhuti, B.; Kelly, M.A.; Luck, P.J.; Drake, M.A.; Foegeding, E.A. Roles of charge interactions on astringency of whey proteins at low pH. J. Dairy Sci. 2010, 93, 1890–1899. [Google Scholar] [CrossRef] [PubMed]
- Fontoin, H.; Saucier, C.; Teissedre, P.L.; Glories, Y. Effect of pH, ethanol and acidity on astringency and bitterness of grape seed tannin oligomers in model wine solution. Food Qual. Prefer. 2008, 19, 286–291. [Google Scholar] [CrossRef]
- De Freitas, V.; Glories, Y. Concentration and compositional changes of procyanidins in grape seeds and skins of white Vitis vinifera varieties. J. Sci. Food Agric. 1999, 79, 1601–1606. [Google Scholar] [CrossRef]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, Y.; Carde, J.P.; Merillon, J.M.; Hamdi, S. Characterization of a grapevine R2R3-MYB transcription factor that regulates the phenylpropanoid pathway. Plant Physiol. 2006, 140, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Roby, G.; Harbertson, J.F.; Adams, D.A.; Matthews, M.A. Berry size and wine water deficits as factors in winegrape composition: Anthocyanins and tannins. Aust. J. Grape Wine Res. 2004, 10, 100–107. [Google Scholar] [CrossRef]
- Yen, W.J.; Chang, L.W.; Duh, P.D. Antioxidant activity of peanut seed testa and its antioxidative component, ethyl protocatechuate. Food Sci. Technol. 2005, 38, 193–200. [Google Scholar]
- Nepote, V.; Grosso, N.R.; Guzman, C.A. Optimization of extraction of phenolic antioxidant from peanut skins. J. Sci. Food Agric. 2005, 85, 33–38. [Google Scholar] [CrossRef]
- Yu, J.; Ahmedna, M.; Goktepe, I. Effects of processing methods and extraction solvents on concentration and antioxidant activity of peanut skin phenolics. Food Chem. 2005, 90, 199–206. [Google Scholar] [CrossRef]
- Prieur, C.; Rigaud, J.; Cheynier, V.; Moutounet, M. Oligomeric and polymeric procyanidins from grape seeds. Phytochemistry 1994, 36, 781–784. [Google Scholar] [CrossRef]
- Souquet, J.-M.; Cheynier, V.; Brossaud, F.; Moutounet, M. Polymeric proanthocyanidins from grape skins. Phytochemistry 1996, 43, 509–512. [Google Scholar] [CrossRef]
- Tsujita, T.; Shintani, T.; Sato, H. Preparation and characterisation of peanut seed skin polyphenols. Food Chem. 2014, 151, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Diago, A.; Dizy, M.; Fernández-Zurbano, P. Contribution of low molecular weight phenols to bitter taste and mouthfeel properties in red wines. Food Chem. 2014, 154, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Li, P.M.; Du, G.R.; Ma, F.W. Phenolics concentration and antioxidant capacity of different fruit tissues of astringent versus non-astringent persimmons. Sci. Hortic. 2011, 129, 710–714. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Apple peels as a value-added food ingredient. J. Agric. Food Chem. 2003, 51, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.C.G.; Gil-Izquierdo, A.; Gil, M.I. Comparative study of six pear cultivars in terms of their phenolic and vitamin C contents and antioxidant capacity. J. Sci. Food Agric. 2003, 83, 995–1003. [Google Scholar] [CrossRef]
- Remorini, D.; Tavarini, S.; Degl’Innocenti, E.; Loreti, F.; Massai, R.; Guidi, L. Effect of rootstocks and harvesting time on the nutritional quality of peel and pulp of peach fruits. Food Chem. 2008, 110, 361–367. [Google Scholar] [CrossRef]
- Kim, H.; Moon, J.Y.; Kim, H.; Lee, D.S.; Cho, M.; Choi, H.K.; Kim, Y.S.; Mosaddik, A.; Cho, S.K. Antioxidant and antiproliferative activities of mango (Mangifera indica L.) pulp and peel. Food Chem. 2010, 12, 429–436. [Google Scholar] [CrossRef]
- Li, Y.; Guo, C.; Yang, J.; Wei, J.; Xu, J.; Cheng, S. Evaluation of antioxidant properties of pomegranate peel extract in comparison with pomegranate pulp extract. Food Chem. 2006, 96, 254–260. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Ferreres, F.; Domingues, A.L.; Seabra, R.M.; Ferreira, M.A. Phenolic profile of quince fruit (Cydonia oblonga Miller) (Pulp and Peel). J. Agric. Food Chem. 2002, 50, 4615–4618. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.W.; Conn, E.E. The cinnamic acid 4-hydroxylase of pea seedlings. Arch. Biochem. Biophys. 1967, 122, 256–258. [Google Scholar] [CrossRef] [PubMed]
- Holton, T.A.; Cornish, E.C. Genetics and biochemistry of anthocyanin biosynthesis. Plant Cell 1995, 7, 1071–1083. [Google Scholar] [CrossRef] [PubMed]
- Aida, R.; Yoshida, K.; Kondo, T.; Kishimoto, S.; Shibata, M. Copigmentation gives bluer flowers on transgenic torenia plants with the antisense dihydroflavonol-4-reductase gene. Plant Sci. 2000, 160, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Petit, P.; Granier, T.; d’Estaintot, B.L.; Manigand, C.; Bathany, K.; Schmitter, J.M.; Lauvergeat, V.; Hamdi, S.; Gallois, B. Crystal structure of grape dihydroflavonol 4-reductase, a key enzyme in flavonoid biosynthesis. J. Mol. Biol. 2007, 368, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Trabelsi, N.; Petit, P.; Manigand, C.; d’Estaintot, B.L.; Granier, T.; Chaudiere, J.; Gallois, B. Structural evidence for the inhibition of grape dihydroflavonol 4-reductase by flavonols. Acta Crystallogr. Sect. D: Biol. Crystallogr. 2008, 64, 883–891. [Google Scholar] [CrossRef]
- O’Reilly, C.; Shepherd, N.S.; Pereira, A.; Schwarz-Sommer, Z.; Bertram, I.; Robertson, D.S.; Peterson, P.A.; Saedler, H. Molecular cloning of the a1 locus of Zea mays using the transposable elements En and Mu1. EMBO J. 1985, 4, 877–882. [Google Scholar] [PubMed]
- Beld, M.; Martin, C.; Huits, H.; Stuitje, A.R.; Gerats, A.G. Flavonoid synthesis in Petunia hybrida: Partial charac-terization of dihydroflavonol-4-reductase genes. Plant Mol. Biol. 1989, 113, 491–502. [Google Scholar] [CrossRef]
- Lo Piero, A.R.; Puglisi, I.; Petrone, G. Gene characterization, analysis of expression and in vitro synthesis of dihydroflavonol 4-reductase from Citrus sinensis (L.) Osbeck. Phytochemistry 2006, 67, 684–695. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Li, Y.H.; Xu, Z.R.; Yan, H.F.; Homma, S.; Kawabata, S. Ultraviolet A-specific induction of anthocyanin biosynthesis in the swollen hypocotyls of turnip (Brassica rapa). J. Exp. Bot. 2007, 58, 1771–1781. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, J.; Kühnel, C.; Brandt, J.; Duy, D.; Punyasiri, P.A.N.; Forkmann, G.; Fischer, T.C. Biosynthesis of flavan 3-ols by leucoanthocyanidin 4-reductases and anthocyanidin reductases in leaves of grape (Vitis vinifera L.), apple (Malus 3 domestica Borkh.) and other crops. Plant Physiol. Biochem. 2006, 44, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Bogs, J.; Downey, M.O.; Harvey, J.S.; Ashton, A.R.; Tanner, G.J.; Robinson, S.P. Proanthocyanidin synthesis and expression of genes encoding leucoanthocyanidin reductase and anthocyanidin reductase in developing grape berries and grapevine leaves. Plant Physiol. 2005, 139, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Punyasiri, P.A.N.; Abeysinghe, I.S.; Kumar, V.; Treutter, D.; Duy, D.; Gosch, C.; Martens, S.; Forkmann, G.; Fischer, T.C. Flavonoid biosynthesis in the tea plant Camellia sinensis: Properties of enzymes of the prominent epicatechin and catechin pathways. Arch. Biochem. Biophys. 2004, 431, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.R.M.; D’Amico, E.; Preuss, A.; Carbone, F.; de Vos, C.H.; Deiml, B.; Mourgues, F.; Perrotta, G.; Fischer, T.C.; Bovy, A.G.; et al. Characterization of major enzymes and genes involved in flavonoid and proanthocyanidin biosynthesis during fruit development in strawberry (Fragari × ananassa). Arch. Biochem. Biophys. 2007, 465, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Hiraoka, K.; Kobayashi, S.; Honda, C.; Terahara, N. Changes in the expression of anthoc-yanin biosynthetic genes during apple development. J. Am. Soc. Hortic. Sci. 2002, 127, 971–976. [Google Scholar]
- Takos, A.M.; Ubi, B.E.; Robinson, S.P.; Walker, A.R. Condensed tannin biosynthesis genes are regulated separately from other flavonoid biosynthesis genes in apple fruit skin. Plant Sci. 2006, 170, 487–499. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Q.L.; Luo, Z.R. Isolation and expression of gene encoding leuc-oanthocyanidin reductase from Diospyros kaki during fruit development. Biol. Plant 2010, 54, 707–710. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, J.; Xin, X.P.; Xu, Y.L.; Liu, Y.; Wei, Y.J.; Fu, H.H. Bioinformatics Analysis of LDOX/ANS Gene in Different Plants. Genomics Appl. Biol. 2010, 29, 815–822. [Google Scholar]
- Xie, D.Y.; Sharma, S.B.; Paiva, N.L.; Ferreira, D.; Dixon, R.A. Role of anthocyanidin reductase, encoded by BAN-YULS in plant flavonoid biosynthesis. Science 2003, 299, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Gargouri, M.; Manigand, C.; Mauge, C.; Granier, T.; Langlois d’Estaintot, B.; Cala, O.; Pianet, I.; Bathany, K.; Chaudiere, J.; Gallois, B. Structure and epimerase activity of anthocyanidin reductase from Vitis vinifera. Acta Crystallogr. Sect. D Biol. Crystallogr. 2009, 65, 989–1000. [Google Scholar] [CrossRef]
- Pang, Y.Z.; Abeysinghe, I.S.B.; He, J.; He, X.Z.; Huhman, D.; Mewan, K.M.; Sumner, L.W.; Yun, J.F.; Dixon, R.A. Functional characterization proanthocyanidin pathway enzymes from tea and their application for metabolic engineering. Plant Physiol. 2013, 161, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.W.; Breen, P.J. Activity of phenylalanine ammonia-lyase and concentration of anthocyanins and Phenolics in developing strawberry fruit. J. Am. Soc. Hortic. 1991, 116, 865–869. [Google Scholar]
- Blankenship, S.M.; Vnrath, C.R. PAL and dethylene-content during maturation of red and golden delicious apples. Phytochemistry 1987, 27, 1001–1003. [Google Scholar] [CrossRef]
- Sun, F.; Zhang, P.Y.; Guo, M.R.; Yu, W.Q.; Chen, K.S. Burdock fructooligosaccharide induces fungal resistance in postharvest Kyoho grapes by activating the salicylic acid-dependent pathway and inhibiting browning. Food Chem. 2013, 138, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, M.M.; Hoysame, R.M. Biochemical and regulatory aspects of anthocyanin synthesis in apple and pears. J. South. Afr. Soc. Hortic. Sci. 1995, 5, 1–6. [Google Scholar]
- Lu, S.F.; Zhou, Y.H.; Li, L.G.; Chiang, V.L. Distinct Roles of Cinnamate 4-hydroxylase Genes in Populus. Plant Cell Physiol. 2006, 47, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, Y.B.; Kim, Y.; Lee, M.Y.; Park, S.U. Overexpression of Cinnamate 4-Hydroxylase and 4-Coumaroyl CoA Ligase Prompted Flavone Accumulation in Scutellaria baicalensis Hairy Roots. Nat. Prod. Commun. 2014, 9, 803–807. [Google Scholar] [PubMed]
- Ryan, K.G.; Swinny, E.E.; Winefield, C.; Markham, K.R. Flavonoids and UV photoprotection in Arabidopsis mutants. Z. Naturforschung 2001, 56, 745–754. [Google Scholar]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Gould, K.S. Nature’s swiss army knife: The diverse protective roles of anthocyanins in leaves. J. Biomed. Biotechnol. 2004, 5, 314–320. [Google Scholar] [CrossRef]
- Xie, D.Y.; Jackson, L.A.; Cooper, J.D.; Ferreira, D.; Paiva, N.L. Molecular and biochemical analysis of two cDNA clones encoding dihydroflavonol-4-reductase from Medicago truncatula. Plant Physiol. 2004, 134, 979–994. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Gou, J.; Jia, Z.; Yang, L.; Sun, Y.; Xiao, X.; Song, F.; Luo, K. Molecular cloning and characterization of two genes encoding dihydroflavonol-4-reductase from Populus trichocarpa. PLoS One 2012, 7, e30364. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, J.; Chen, F.; Lv, X.; Fu, C.; Zhao, D.; Hua, X.; Zhao, Q. Molecular characterization and expression analysis of dihydroflavonol 4-reductase (DFR) gene in Saussurea medusa. Mol. Biol. Rep. 2012, 39, 2991–2999. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Iwasaka, R.; Shimada, N.; Ayabe, S.; Aoki, T.; Sakuta, M. Transcriptional control of the dihydroflavonol 4-reductase multigene family in Lotus japonicus. J. Plant Res. 2010, 123, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.J.; Constabel, C.P. Molecular analysis of herbivore-induced condensed tannin synthesis: Cloning and expression of dihydroflavonol reductase from trembling aspen (Populus tremuloides). Plant J. 2002, 32, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Crifò, T.; Puglisi, I.; Petrone, G.; Recupero, G.R.; Lo Piero, A.R. Expression analysis in response to low temperature stress in blood oranges: Implication of the flavonoid biosynthetic pathway. Gene 2011, 476, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Koyama, K.; Ikeda, H.; Poudel, P.R.; Goto-Yamamoto, N. Light quality affects flavonoid biosynthesis in young berries of cabernet Sauvignon grape. Phytochemistry 2012, 78, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Ollé, D.; Guiraud, J.L.; Souquet, J.M.; Terrier, N.; Ageorges, A.; Cheynier, V.; Verries, C. Effect of pre- and post-veraison water deficit on proanthocyanidin and anthocy anin accumulation during Shiraz berry development. Aust. J. Grape Wine Res. 2011, 17, 90–100. [Google Scholar] [CrossRef]
- Moalem-Beno, D.; Tamari, G.; Leitner-Dagan, Y.; Borochov, A.; Weiss, D. Sugar-dependent gibberellin-induced chalcone synthase gene expression in petunia corollas. Plant Physiol. 1997, 113, 419–424. [Google Scholar] [PubMed]
- Nesi, N.; Debeaujon, I.; Jond, C.; Pelletier, G.; Caboche, M.; Lepiniec, L. The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in Arabidopsis siliques. Plant Cell 2000, 12, 1863–1878. [Google Scholar] [CrossRef] [PubMed]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verries, C.; Cheynier, V.; Romieu, C. Ectopic expression of VvMybPA2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol. 2009, 149, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Ikegami, A.; Tsujimoto, T.; Kobayashi, S.; Sato, A.; Kono, A.; Yonemori, K. DkMyb4 is a Myb transcription factor involved in proanthocyanidin biosynthesis in persimmon fruit. Plant Physiol. 2009, 151, 2028–2045. [Google Scholar] [CrossRef] [PubMed]
- Akagi, T.; Ikegami, A.; Yonemori, K. DkMyb2 wound-induced transcription factor of persimmon (Diospyros kaki Thunb.) contributes to proanthocyanidin regulation. Planta 2010, 232, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Hichri, I.; Heppel, S.C.; Pillet, J.; Léon, C.; Czemmel, S.; Delrot, S.; Lauvergeat, V.; Bogs, J. The basic helix-loop-helix transcription factor MYC1 is involved in the regulation of the flavonoid biosynthesis pathway in grapevine. Mol. Plant 2010, 3, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Matus, J.T.; Poupin, M.J.; Canon, P.; Bordeu, E.; Alcalde, J.A.; Arce-Johnson, R. Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.). Plant Mol. Biol. 2010, 72, 607–620. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, M.; Tian, H.; Luo, X.; Qi, X.; Chen, X. Molecular Progress in Research on Fruit Astringency. Molecules 2015, 20, 1434-1451. https://doi.org/10.3390/molecules20011434
He M, Tian H, Luo X, Qi X, Chen X. Molecular Progress in Research on Fruit Astringency. Molecules. 2015; 20(1):1434-1451. https://doi.org/10.3390/molecules20011434
Chicago/Turabian StyleHe, Min, Henglu Tian, Xiaowen Luo, Xiaohua Qi, and Xuehao Chen. 2015. "Molecular Progress in Research on Fruit Astringency" Molecules 20, no. 1: 1434-1451. https://doi.org/10.3390/molecules20011434
APA StyleHe, M., Tian, H., Luo, X., Qi, X., & Chen, X. (2015). Molecular Progress in Research on Fruit Astringency. Molecules, 20(1), 1434-1451. https://doi.org/10.3390/molecules20011434