
Oral Administration of Achyranthis radix Extract Prevents TMA-induced Allergic Contact Dermatitis by Regulating Th2 Cytokine and Chemokine Production in Vivo

Abstract
:
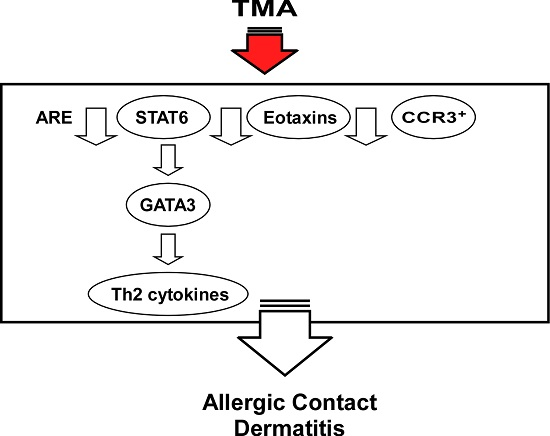
1. Introduction
2. Results and Discussion
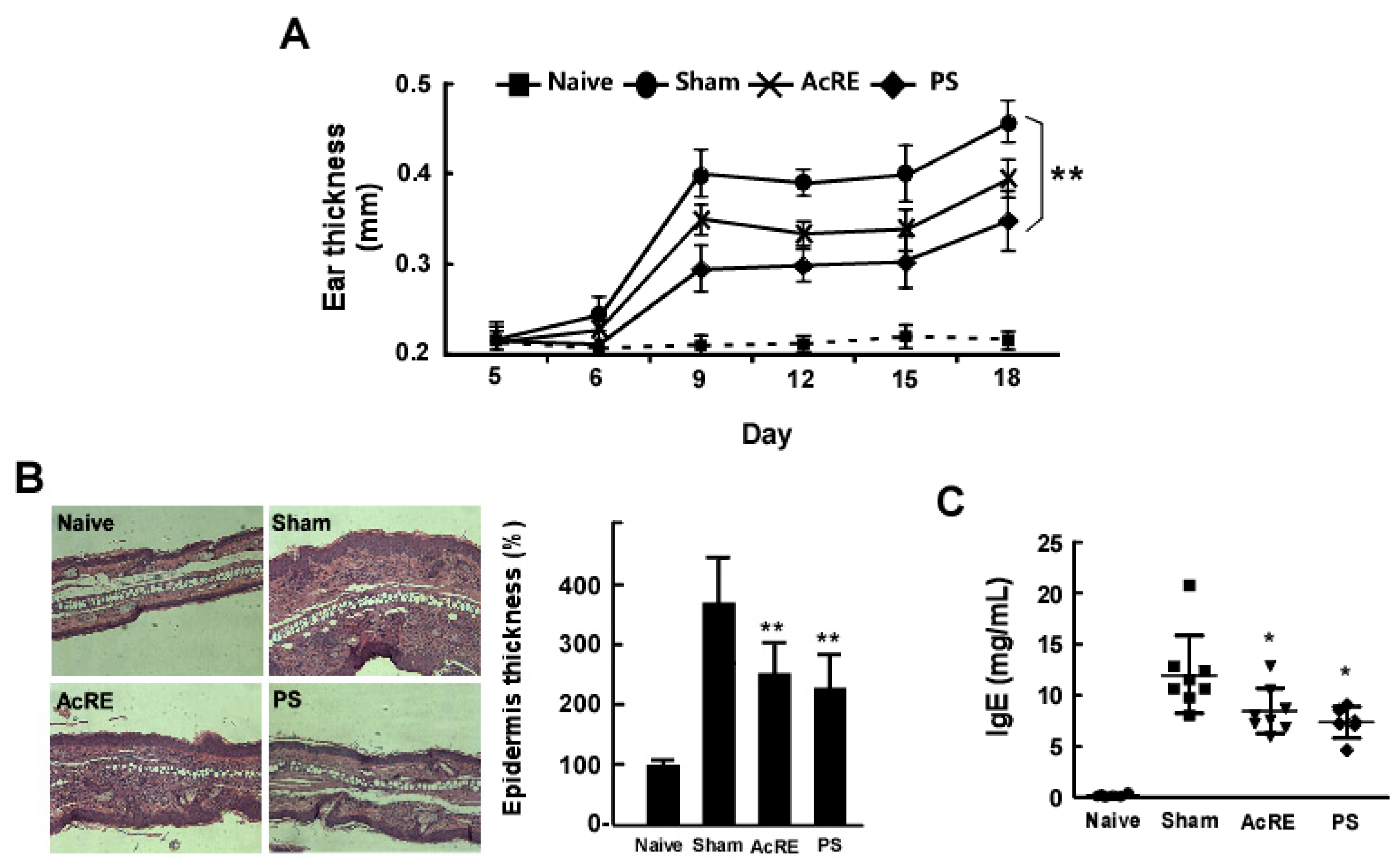
2.1. AcRE Attenuates TMA-Induced Ear Swelling and Increases in Epidermal Thickness and IgE Production in Mouse Ear Tissue

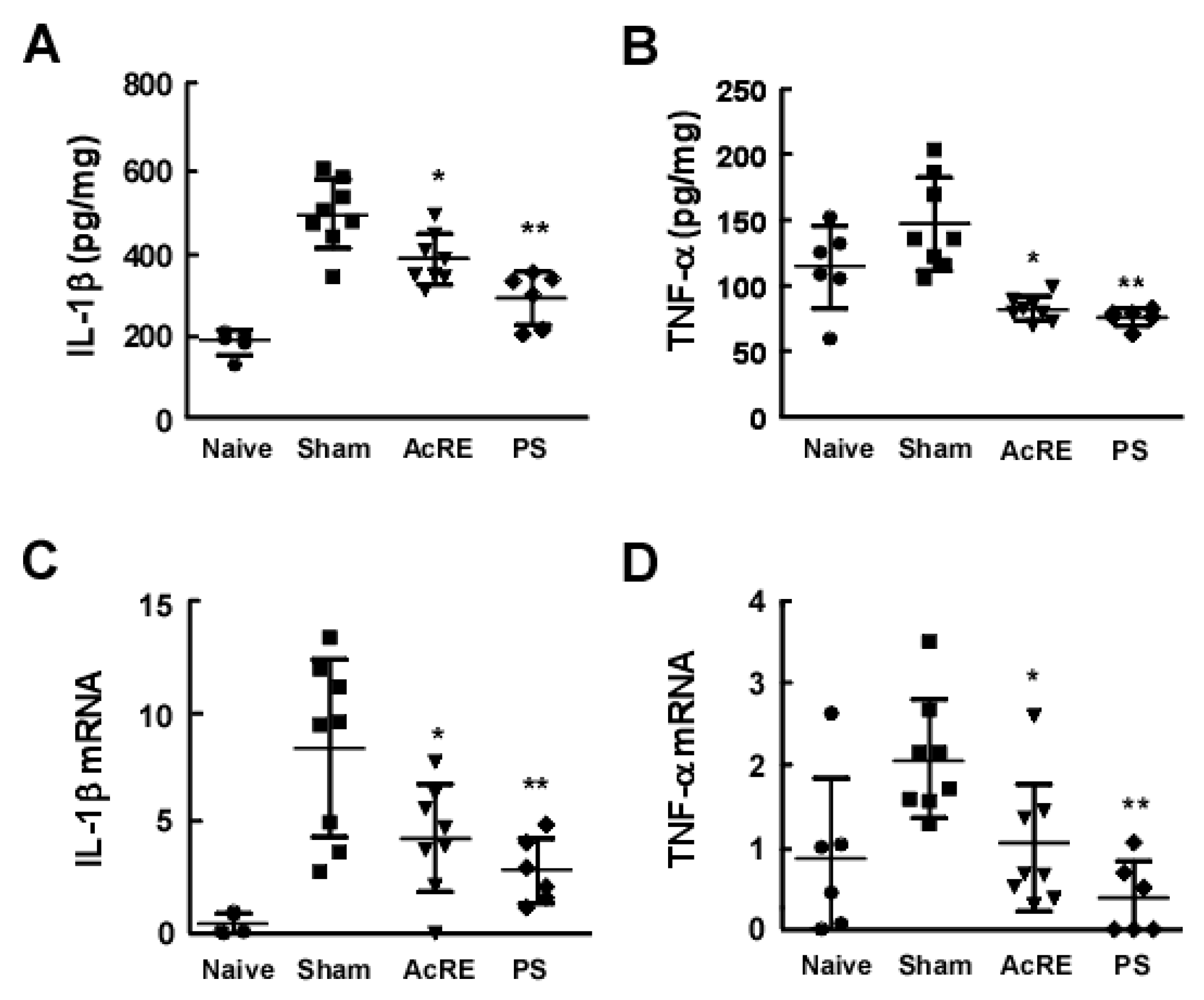
2.2. AcRE Suppresses TMA-Induced Pro-Inflammatory Cytokine Production and mRNA Expression in the Mouse Ear
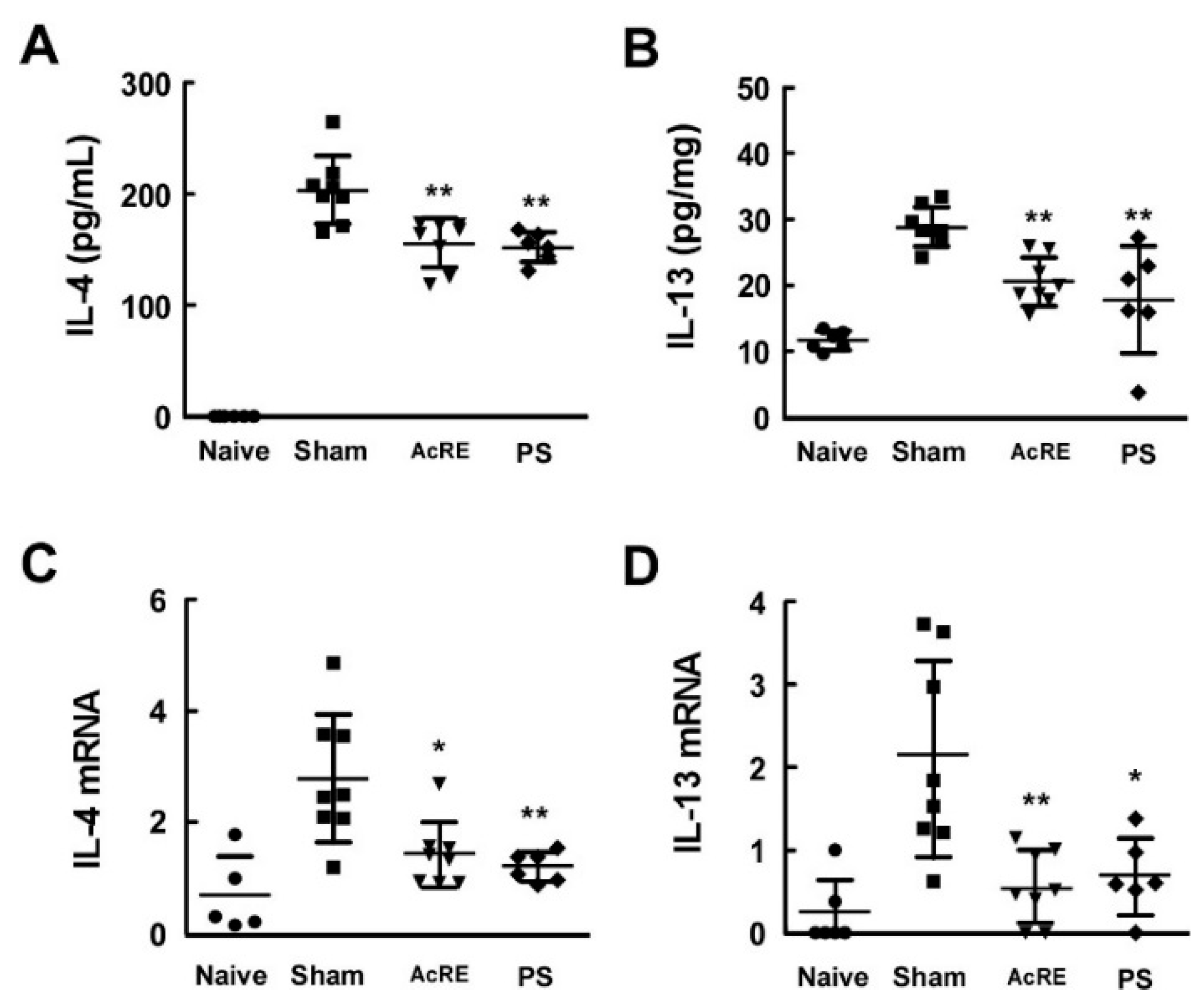
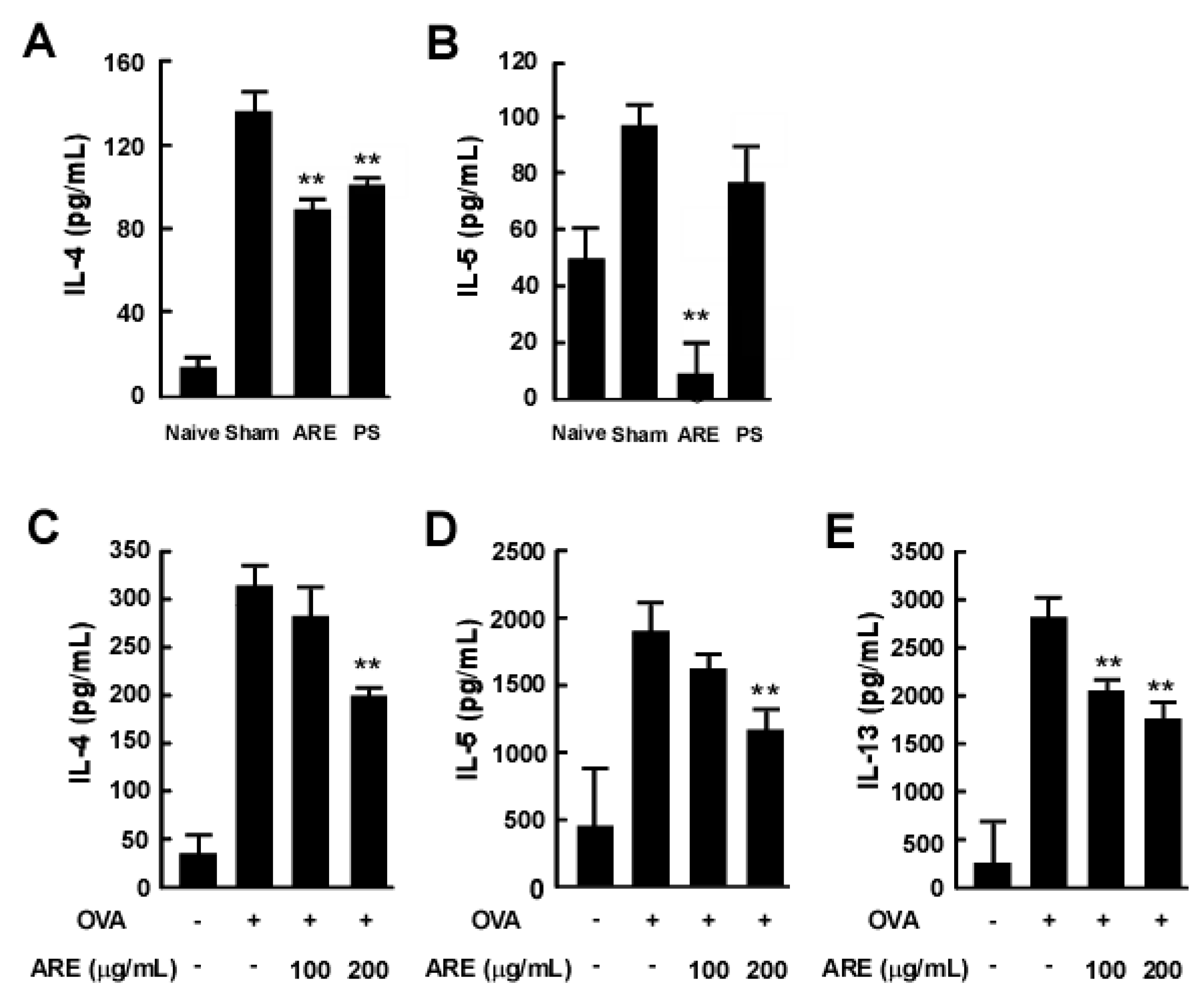
2.3. AcRE Suppresses TMA-Induced Th2 Cytokine Production in the Mouse Ear


2.4. AcRE Suppresses TMA-Induced Th2 Cytokine Production in the Mouse Ear

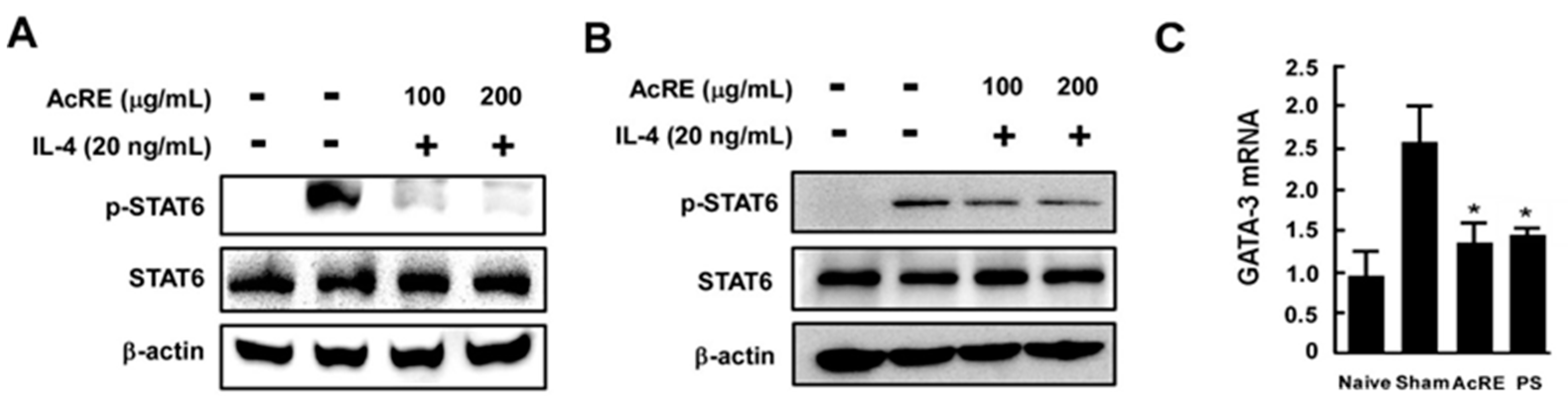
2.5. AcRE Suppresses IL-4-Induced STAT-6 Phosphorylation in Primary Cultured Mouse Splenocytes and Immortalized Human Keratinocytes
2.6. AcRE Affects TMA-Induced CCR3+ Cell Infiltration and CCL11 mRNA Production in Mouse Ear Tissue and IL-4-Induced CCL26 mRNA Expression in HaCaT Cells


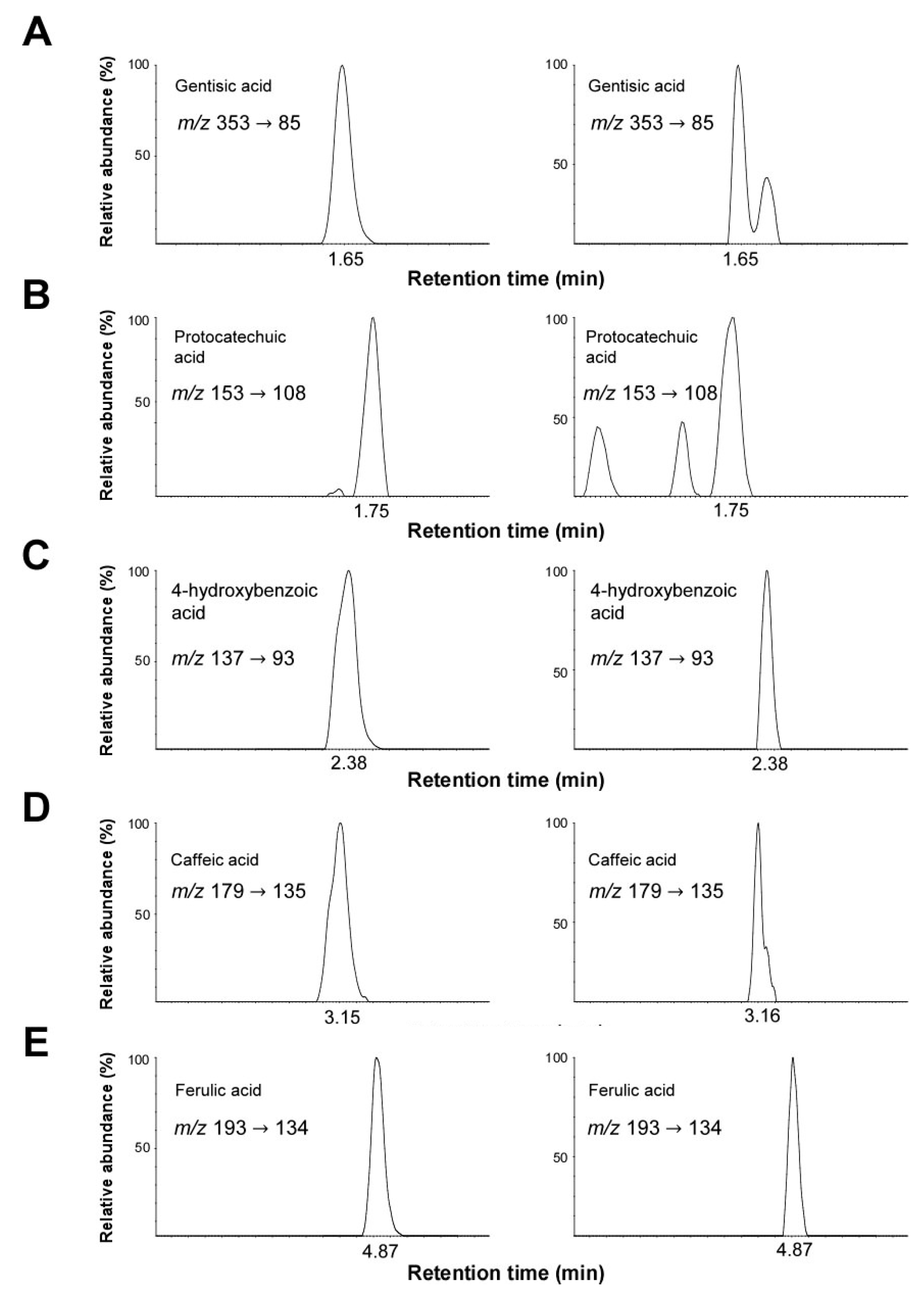
2.7. Identification and Quantification of Phenolic Compounds by UPLC-ESI-MS/MS Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RT a (min) | Content (μg/g) b | MRM c Transition (m/z) |
|---|---|---|---|
| Gentisic acid | 1.65 | 0.64 ± 0.2 | 353 → 93 |
| Protocatechuic acid | 1.75 | 2.69 ± 0.1 | 153 → 108 |
| 4-hydroxybenzoic acid | 2.38 | 5.59 ± 0.3 | 137 → 93 |
| Caffeic acid | 3.15 | 4.21 ± 0.1 | 179 → 135 |
| Feluric acid | 4.87 | 14.78 ± 0.4 | 193 → 134 |
2.8. Discussion
3. Experimental Section
3.1. Materials
3.2. Sample Preparationx
3.3. Animals
3.4. Induction of Allergic Contact Dermatitis and Experimental Schedule
3.5. Preparation of Mice Ears for ELISA Assay
3.6. Culture of Draining Lymph Node
3.7. Sensitization and Challenge with OVA and Preparation of Splenocyte Cultures
3.8. Cytokine Measurement by ELISA Assay
3.9. RNA Isolation and Quantitative Real-Time PCR Analyses
3.10. Western Blot Analysis
3.11. Measurement of Epidermal Thickness
3.12. Immunohistohemistry
3.13. UPLC-MS/MS Analysis
3.14. StatisticalAnalysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peiser, M.; Tralau, T.; Heidler, J.; Api, A.M.; Arts, J.H.; Basketter, D.A.; English, J.; Diepgen, T.L.; Fuhlbrigge, R.C.; Gaspari, A.A.; et al. Allergic contact dermatitis: Epidemiology, molecular mechanisms, in vitro methods and regulatory aspects. Current knowledge assembled at an international workshop at BfR, Germany. Cell Mol. Life Sci. 2012, 69, 763–781. [Google Scholar] [CrossRef] [PubMed]
- Dearman, R.J.; Kimber, I. Differential stimulation of immune function by respiratory and contact chemical allergens. Immunology 1991, 72, 563–570. [Google Scholar] [PubMed]
- Schneider, C.; Docke, W.D.F.; Zollner, T.M.; Rose, L. Chronic Mouse Model of TMA-Induced Contact Hypersensitivity. J. Investig. Dermatol. 2009, 129, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.H.; Igyarto, B.Z.; Gaspari, A.A. Early immune events in the induction of allergic contact dermatitis. Nat. Rev. Immunol. 2012, 12, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Honda, T.; Egawa, G.; Grabbe, S.; Kabashima, K. Update of immune events in the murine contact hypersensitivity model: Toward the understanding of allergic contact dermatitis. J. Investig. Dermatol. 2013, 133, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Cumberbatch, M.; Dearman, R.J.; Kimber, I. Langerhans cells require signals from both tumour necrosis factor-alpha and interleukin-1 beta for migration. Immunology 1997, 92, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Akdis, M.; Blaser, K.; Akdis, C.A. T regulatory cells in allergy: Novel concepts in the pathogenesis, prevention, and treatment of allergic diseases. J. Allergy Clin. Immunol. 2005, 116, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Roh, K.B.; Kim, I.H.; Kim, Y.S.; Lee, M.; Lee, J.A.; Jung, E.; Park, D. Synephrine inhibits eotaxin-1 expression via the STAT6 signaling pathway. Molecules 2014, 19, 11883–11895. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. Peripheral CD4+ T-cell differentiation regulated by networks of cytokines and transcription factors. Immunol. Rev. 2010, 238, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Flavell, R.A. The transcription factor GATA-3 is necessary and sufficient for Th2 cytokine gene expression in CD4 T cells. Cell 1997, 89, 587–596. [Google Scholar] [CrossRef]
- Onodera, A.; Yamashita, M.; Endo, Y.; Kuwahara, M.; Tofukuji, S.; Hosokawa, H.; Kanai, A.; Suzuki, Y.; Nakayama, T. STAT6-mediated displacement of polycomb by trithorax complex establishes long-term maintenance of GATA3 expression in T helper type 2 cells. J. Exp. Med. 2010, 207, 2493–2506. [Google Scholar] [CrossRef] [PubMed]
- Pivarcsi, A.; Homey, B. Chemokine networks in atopic dermatitis: Traffic signals of disease. Curr. Allergy Asthma Rep. 2005, 5, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Owczarek, W.; Paplinska, M.; Targowski, T.; Jahnz-Rozyk, K.; Paluchowska, E.; Kucharczyk, A.; Kasztalewicz, B. Analysis of eotaxin 1/CCL11, eotaxin 2/CCL24 and eotaxin 3/CCL26 expression in lesional and non-lesional skin of patients with atopic dermatitis. Cytokine 2010, 50, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Uratsuji, H.; Saeki, H.; Kagami, S.; Tsunemi, Y.; Komine, M.; Tamaki, K. CCR4 and CCR10 are expressed on epidermal keratinocytes and are involved in cutaneous immune reaction. Cytokine 2008, 44, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Pawankar, R.; Kimura, Y.; Kawana, S. Increased expression of RANTES, CCR3 and CCR5 in the lesional skin of patients with atopic eczema. Int. Arch. Allergy Immunol. 2006, 139, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Kagami, S.; Saeki, H.; Komine, M.; Kakinuma, T.; Tsunemi, Y.; Nakamura, K.; Sasaki, K.; Asahina, A.; Tamaki, K. Interleukin-4 and interleukin-13 enhance CCL26 production in a human keratinocyte cell line, HaCaT cells. Clin. Exp. Immunol. 2005, 141, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Banwell, M.E.; Tolley, N.S.; Williams, T.J.; Mitchell, T.J. Regulation of human eotaxin-3/CCL26 expression: Modulation by cytokines and glucocorticoids. Cytokine 2002, 17, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.H.; Ngoc, T.M.; Bae, K.; Kim, Y.S.; Kim, H.P. Inhibition of experimental atopic dermatitis by rhubarb (rhizomes of Rheum tanguticum) and 5-lipoxygenase inhibition of its major constituent, emodin. Phytother. Res. 2011, 25, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Cho, S.H. Effects of Korean red ginseng extract on the prevention of atopic dermatitis and its mechanism on early lesions in a murine model. J. Ethnopharmacol. 2013, 145, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jin, S.W.; Han, E.H.; Park, B.H.; Kim, H.G.; Khanal, T.; Hwang, Y.P.; Do, M.T.; Lee, H.S.; Chung, Y.C.; et al. Platycodon grandiflorum root-derived saponins attenuate atopic dermatitis-like skin lesions via suppression of NF-kappaB and STAT1 and activation of Nrf2/ARE-mediated heme oxygenase-1. Phytomedicine 2014, 21, 1053–1061. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kwon, H.K.; Lee, S.M. SHINBARO, a new herbal medicine with multifunctional mechanism for joint disease: First therapeutic application for the treatment of osteoarthritis. Arch. Pharm. Res. 2011, 34, 1773–1777. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Y.; Wang, R.T.; Tian, N.; Li, L.; Lin, N.; Chen, W.H. Effect of Huogu II Formula (II) with medicinal guide Radix Achyranthis Bidentatae on bone marrow stem cells directional homing to necrosis area after osteonecrosis of the femoral head in rabbit. Chin. J. Integr. Med. 2012, 18, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.T.; Kong, X.Y.; Tian, N.; Lin, N.; Chen, W.H. Effects of promoting blood circulation to remove meridian obstruction combined with medicinal guides on related osteogenesis factors in rabbits with femoral head necrosis. China J. Orthop. Traumatol. 2013, 26, 591–596. [Google Scholar]
- Siu, W.S.; Zhou, X.; Fung, C.H.; Shum, W.T.; Lau, C.B.; Leung, P.C.; Ko, C.H.; Hung, L.K. Preclinical evaluations on the efficacy of a topical Chinese herbal formula for swelling control and pain relief. J. Ethnopharmacol. 2015, 162, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Neis, M.M.; Peters, B.; Dreuw, A.; Wenzel, J.; Bieber, T.; Mauch, C.; Krieg, T.; Stanzel, S.; Heinrich, P.C.; Merk, H.F.; et al. Enhanced expression levels of IL-31 correlate with IL-4 and IL-13 in atopic and allergic contact dermatitis. J. Allergy Clin. Immunol. 2006, 118, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Piguet, P.F.; Grau, G.E.; Hauser, C.; Vassalli, P. Tumor necrosis factor is a critical mediator in hapten induced irritant and contact hypersensitivity reactions. J. Exp. Med. 1991, 173, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Na, K.M.; Oh, I.; Song, I.H.; Lee, Y.S.; Shin, J.; Kim, T.Y. Kurarinone regulates immune responses through regulation of the JAK/STAT and TCR-mediated signaling pathways. Biochem. Pharmacol. 2013, 85, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Homey, B.; Steinhoff, M.; Ruzicka, T.; Leung, D.Y. Cytokines and chemokines orchestrate atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 118, 178–89. [Google Scholar] [CrossRef] [PubMed]
- Korte, S.M. Corticosteroids in relation to fear, anxiety and psychopathology. Neurosci. Biobehav. Rev. 2001, 25, 117–142. [Google Scholar] [CrossRef]
- Vezzosi, D.; Cartier, D.; Regnier, C.; Otal, P.; Bennet, A.; Parmentier, F.; Plantavid, M.; Lacroix, A.; Lefebvre, H.; Caron, P. Familial adrenocorticotropin-independent macronodular adrenal hyperplasia with aberrant serotonin and vasopressin adrenal receptors. Eur. J. Endocrinol. 2007, 156, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Junghans, V.; Gutgesell, C.; Jung, T.; Neumann, C. Epidermal cytokines IL-1beta, TNF-alpha, and IL-12 in patients with atopic dermatitis: Response to application of house dust mite antigens. J. Investig. Dermatol. 1998, 111, 1184–1188. [Google Scholar] [CrossRef] [PubMed]
- Amerio, P.; Frezzolini, A.; Feliciani, C.; Verdolini, R.; Teofoli, P.; De Pita, O.; Puddu, P. Eotaxins and CCR3 receptor in inflammatory and allergic skin diseases: Therapeutical implications. Curr. Drug Targets Inflamm. Allergy 2003, 2, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, S.K.; Choi, D.W.; Kwon, D.-A.; Kim, M.J.; Seong, K.S.; Shon, D.-H. Oral Administration of Achyranthis radix Extract Prevents TMA-induced Allergic Contact Dermatitis by Regulating Th2 Cytokine and Chemokine Production in Vivo. Molecules 2015, 20, 21584-21596. https://doi.org/10.3390/molecules201219788
Jung SK, Choi DW, Kwon D-A, Kim MJ, Seong KS, Shon D-H. Oral Administration of Achyranthis radix Extract Prevents TMA-induced Allergic Contact Dermatitis by Regulating Th2 Cytokine and Chemokine Production in Vivo. Molecules. 2015; 20(12):21584-21596. https://doi.org/10.3390/molecules201219788
Chicago/Turabian StyleJung, Sung Keun, Dae Woon Choi, Da-Ae Kwon, Min Jung Kim, Ki Seung Seong, and Dong-Hwa Shon. 2015. "Oral Administration of Achyranthis radix Extract Prevents TMA-induced Allergic Contact Dermatitis by Regulating Th2 Cytokine and Chemokine Production in Vivo" Molecules 20, no. 12: 21584-21596. https://doi.org/10.3390/molecules201219788
APA StyleJung, S. K., Choi, D. W., Kwon, D.-A., Kim, M. J., Seong, K. S., & Shon, D.-H. (2015). Oral Administration of Achyranthis radix Extract Prevents TMA-induced Allergic Contact Dermatitis by Regulating Th2 Cytokine and Chemokine Production in Vivo. Molecules, 20(12), 21584-21596. https://doi.org/10.3390/molecules201219788