
Inhibitory Effect of Duabanga grandiflora on MRSA Biofilm Formation via Prevention of Cell-Surface Attachment and PBP2a Production


Abstract
:1. Introduction
2. Results and Discussion
2.1. Results




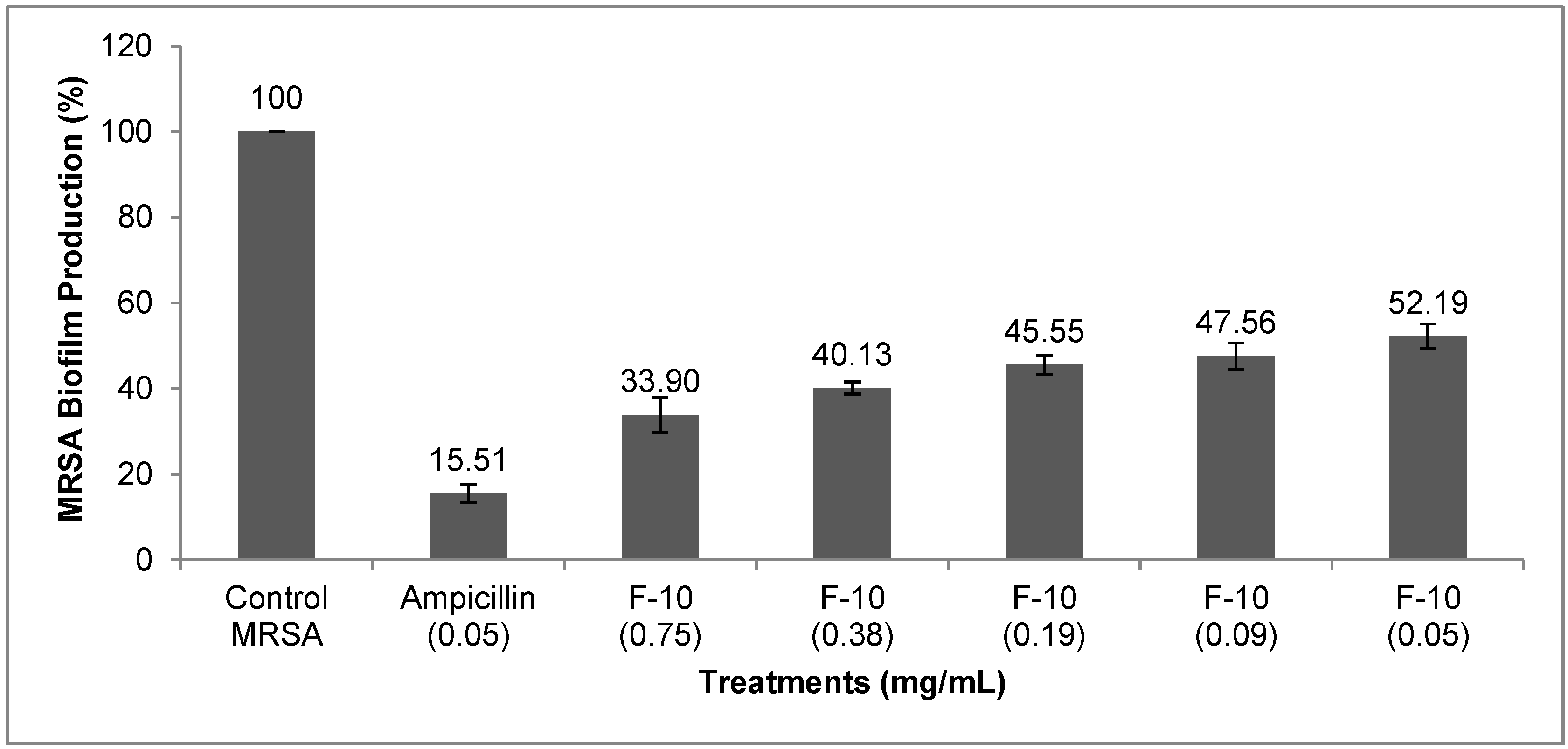{kind=link}
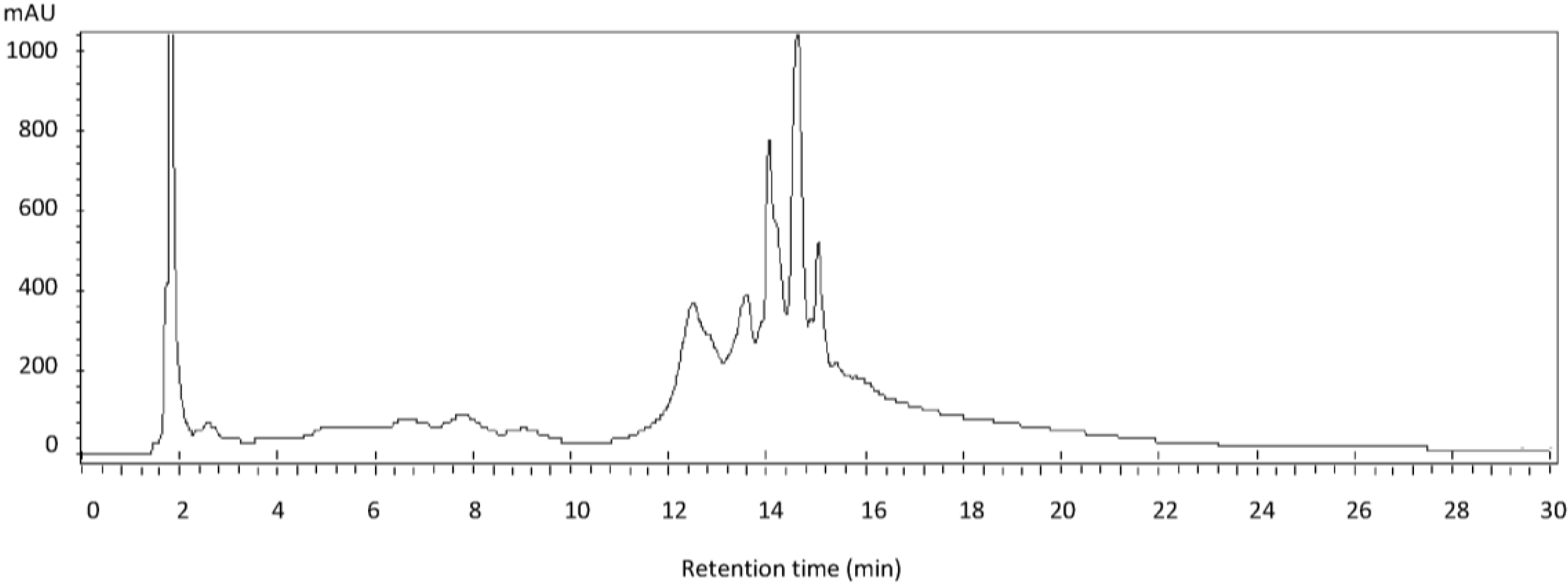{kind=link}
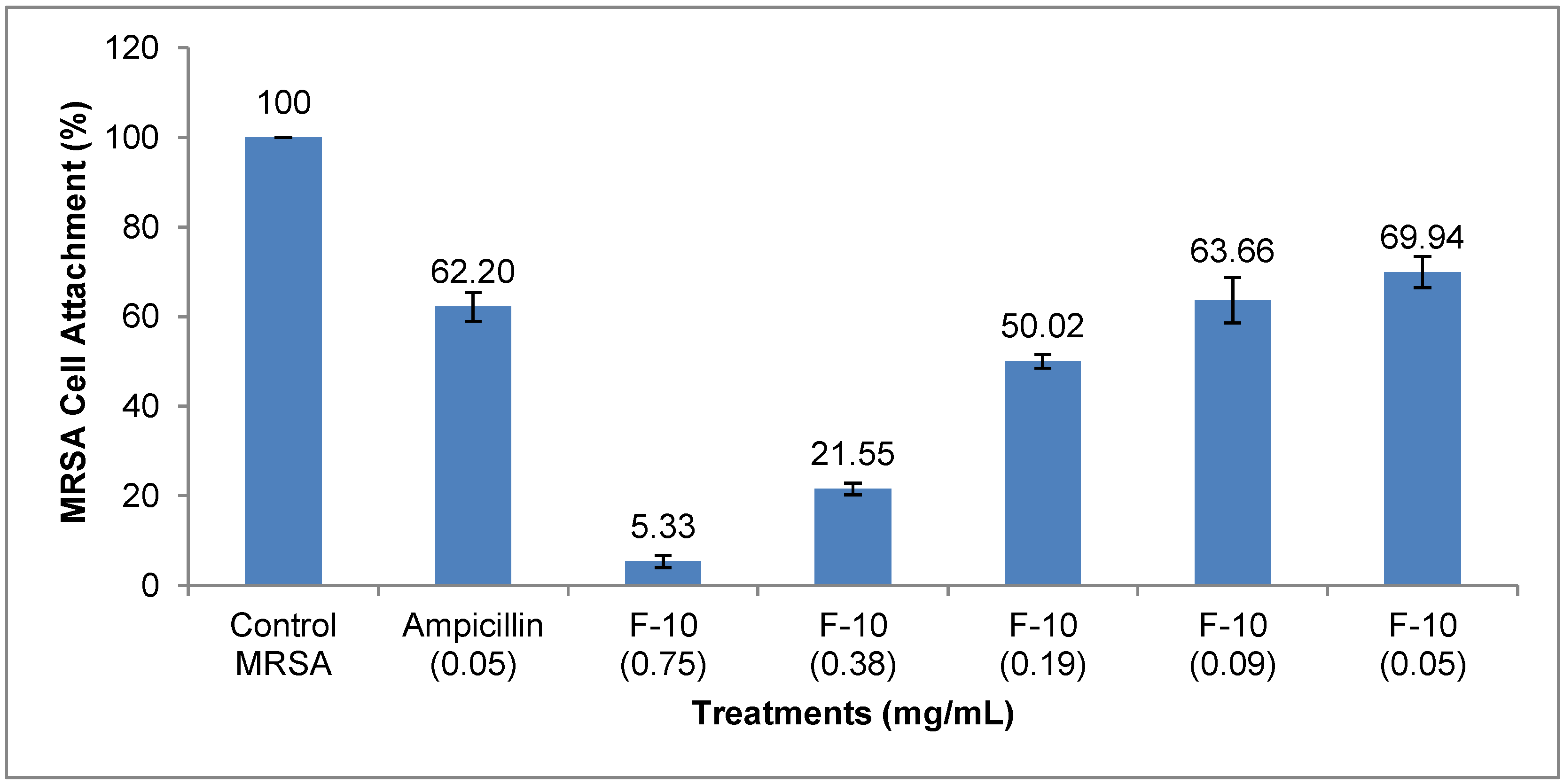{kind=link}
{kind=link}
| Treatments (mg/mL) | Intensity of PBP2a Agglutination |
|---|---|
| Control MRSA | + + |
| Ampicillin (0.05) | + + + |
| F-10 (0.75) | + |
| F-10 (0.38) | + |
| F-10 (0.19) | + |
| F-10 (0.09) | + |
| F-10 (0.05) | + |

2.2. Discussion
3. Experimental Section
3.1. Plant Material
3.2. Extraction and Isolation
3.3. Phytochemical and HPLC Analysis
3.4. Bacterial Strain, Growth Conditions and Determination of MIC Ampicillin
3.5. Inhibition of Biofilm Assay
3.6. Microtiter Attachment Assay
3.7. PBP2a Latex Agglutination Test on MRSA Biofilm
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, J.L.; Patel, R. The challenge of treating biofilm-associated bacterial infections. Clin. Pharmacol. Ther. 2007, 82, 204–209. [Google Scholar]
- Petrelli, D.; Repetto, A.; D’Ercole, S.; Rombini, S.; Ripa, S.; Prenna, M.; Vitali, L.A. Analysis of meticillin-susceptible and meticillin-resistant biofilm-forming Staphylococcus aureus from catheter infections isolated in a large italian hospital. J. Med. Microbiol. 2008, 57, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Antimicrobial resistance in gram-positive bacteria. Am. J. Med. 2006, 34, S11–S19. [Google Scholar] [CrossRef]
- Mack, D. Molecular mechanisms of Staphylococcus epidermidis biofilm formation. J. Hosp. Infect. 1999, 43, S113–S125. [Google Scholar] [CrossRef] [PubMed]
- Götz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.; Becker, P.; Chatterjee, I.; Dobinsky, S.; Knobloch, J.K.M.; Peters, G.; Rohde, H.; Herrmann, M. Mechanisms of biofilm formation in Staphylococcus epidermidis and Staphylococcus aureus: Functional molecules, regulatory circuits, and adaptive responses. Int. J. Med. Microbiol. 2004, 294, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Berger-Bächi, B.; Rohrer, S. Factors influencing methicillin resistance in staphylococci. Arch. Microbiol. 2002, 178, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, C.; Waters, E.M.; Rudkin, J.K.; Schaeffer, C.R.; Lohan, A.J.; Tong, P.; Loftus, B.J.; Pier, G.B.; Fey, P.D.; Massey, R.C.; et al. Methicillin resistance alters the biofilm phenotype and attenuates virulence in Staphylococcus aureus device-associated infections. PLoS Pathog. 2012, 8, e1002626. [Google Scholar] [CrossRef] [PubMed]
- Shankar, R.; Devalla, R.B. Conservation of folk healing practices and commercial medicinal plants with special reference to nagaland. Int. J. Biodivers. Conserv. 2012, 4, 155–163. [Google Scholar] [CrossRef]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Muñoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.C.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus δ-toxin induces allergic skin disease by activating mast cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Othman, M.; Genapathy, S.; Liew, P.S.; Ch’ng, Q.T.; Loh, H.S.; Khoo, T.J.; Wiart, C.; Ting, K.N. Search for antibacterial agents from malaysian rainforest and tropical plants. Nat. Prod. Res. 2011, 25, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Othman, M.; Loh, H.S.; Wiart, C.; Khoo, T.J.; Lim, K.H.; Ting, K.N. Optimal methods for evaluating antimicrobial activities from plant extracts. J. Microbiol. Methods 2011, 84, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Santiago, C.; Pang, E.L.; Lim, K.H.; Loh, H.S.; Ting, K.N. Inhibition of penicillin-binding protein 2a (PBP2a) in methicillin resistant Staphylococcus aureus (MRSA) by combination of ampicillin and a bioactive fraction from Duabanga grandiflora. BMC Complement. Altern. Med. 2014. submitted. [Google Scholar]
- Saleem, M.; Nazir, M.; Ali, M.S.; Hussain, H.; Lee, Y.S.; Riaz, N.; Jabbar, A. Antimicrobial natural products: An update on future antibiotic drug candidates. Nat. Prod. Rep. 2010, 27, 238–254. [Google Scholar] [CrossRef] [PubMed]
- Ozçelik, B.; Orhan, I.; Toker, G. Antiviral and antimicrobial assessment of some selected flavonoids. Z. Naturforsch. 2006, 61, 632–638. [Google Scholar]
- Cushnie, T.P.T.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Overhage, J.; Campisano, A.; Bains, M.; Torfs, E.C.W.; Rehm, B.H.A.; Hancock, R.E.W. Human host defense peptide LL-37 prevents bacterial biofilm formation. Infect. Immun. 2008, 76, 4176–4182. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm Formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- Mataraci, E.; Dosler, S. In vitro Activities of antibiotics and antimicrobial cationic peptides alone and in combination against methicillin-resistant Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 2012, 56, 6366–6371. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.F.J. Detection of methicillin/oxacillin resistance in staphylococci. J. Antimicrob. Chemother. 2001, 48, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.P.; Kinghorn, A.D. Natural Products Isolation; Sarker, S.D., Latif, Z., Gray, A.I., Eds.; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar]
- Santiago, C.; Pang, E.L.; Lim, K.-H.; Loh, H.-S.; Ting, K.N. Reversal of ampicillin resistance in mrsa via inhibition of penicillin-binding protein 2a by Acalypha wilkesiana. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Durham-Colleran, M.W.; Verhoeven, A.B.; van Hoek, M.L. Francisella novicida forms in vitro biofilms mediated by an orphan response regulator. Microb. Ecol. 2010, 59, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.H.; Hu, Z.Q.; Okubo, S.; Hara, Y.; Shimamura, T. Mechanism of synergy between epigallocatechin gallate and beta-lactams against methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2001, 45, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago, C.; Lim, K.-H.; Loh, H.-S.; Ting, K.N. Inhibitory Effect of Duabanga grandiflora on MRSA Biofilm Formation via Prevention of Cell-Surface Attachment and PBP2a Production. Molecules 2015, 20, 4473-4482. https://doi.org/10.3390/molecules20034473
Santiago C, Lim K-H, Loh H-S, Ting KN. Inhibitory Effect of Duabanga grandiflora on MRSA Biofilm Formation via Prevention of Cell-Surface Attachment and PBP2a Production. Molecules. 2015; 20(3):4473-4482. https://doi.org/10.3390/molecules20034473
Chicago/Turabian StyleSantiago, Carolina, Kuan-Hon Lim, Hwei-San Loh, and Kang Nee Ting. 2015. "Inhibitory Effect of Duabanga grandiflora on MRSA Biofilm Formation via Prevention of Cell-Surface Attachment and PBP2a Production" Molecules 20, no. 3: 4473-4482. https://doi.org/10.3390/molecules20034473