
Strophalloside Induces Apoptosis of SGC-7901 Cells through the Mitochondrion-Dependent Caspase-3 Pathway

Abstract
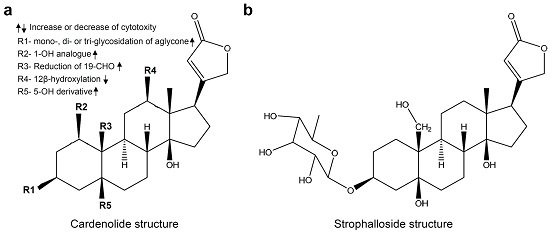
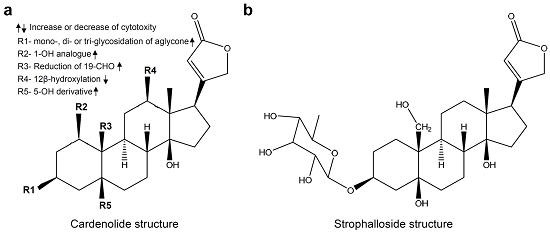
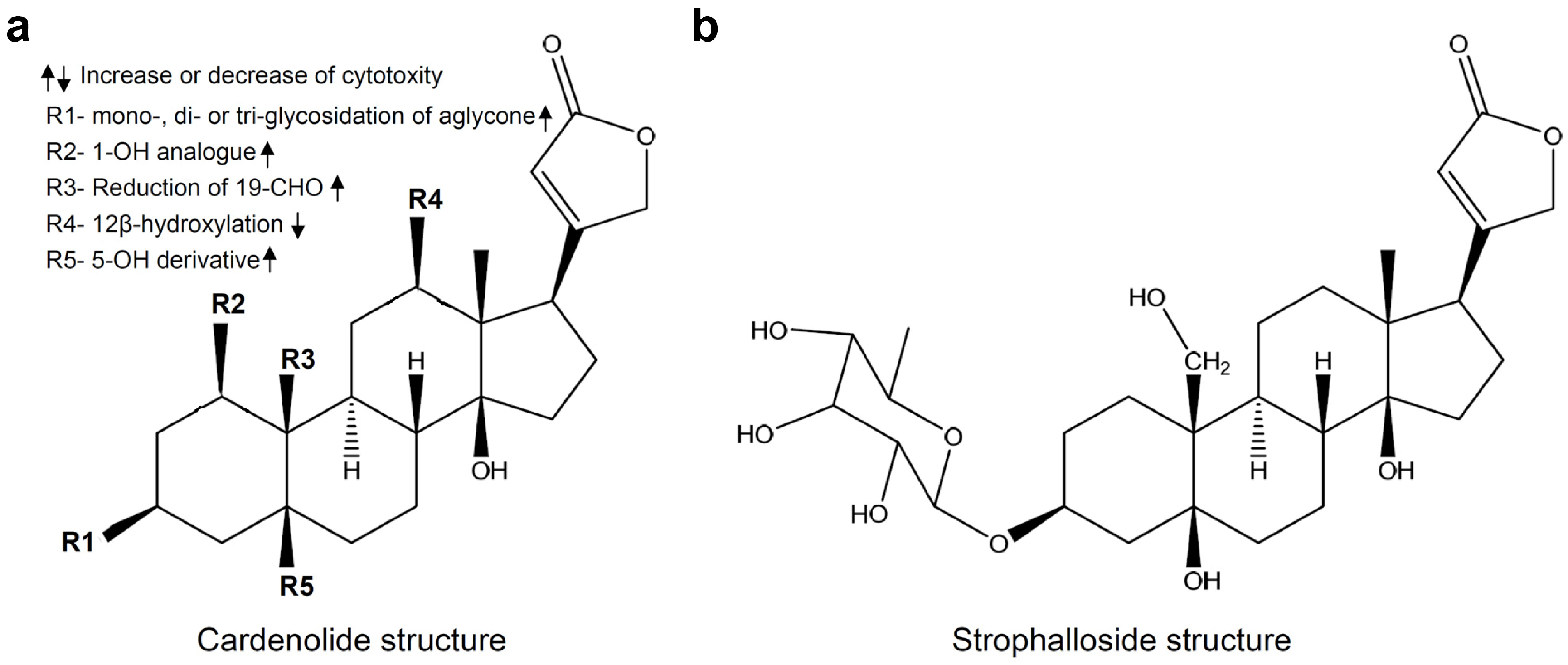
:
1. Introduction

2. Results and Discussion
2.1. Inhibition of SGC-7901 Cell Proliferation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Dose (nM/mL) | 24 h | 48 h | ||
|---|---|---|---|---|---|
| A Value | Inhibition (%) | A Value | Inhibition (%) | ||
| Control | 0.0 | 0.92 ± 0.027 | 00.00 ± 00.00 | 0.975 ± 0.034 * | 00.00 ± 00.00 |
| Strophalloside | 0.12 | 0.855 ± 0.029 * | 7.06 ± 3.15 * | 0.890 ± 0.056 * | 8.71 ± 5.80 * |
| 0.24 | 0.782 ± 0.015 * | 15.00 ± 1.60 * | 0.747 ± 0.040 * | 23.38 ± 4.23 * | |
| 0.47 | 0.675 ± 0.009 * | 26.63 ± 1.02 * | 0.356 ± 0.012 * | 62.56 ± 1.31 * | |
| 0.93 | 0.465 ± 0.005 * | 49.46 ± 0.50 * | 0.104 ± 0.005 * | 89.33 ± 0.57 * | |
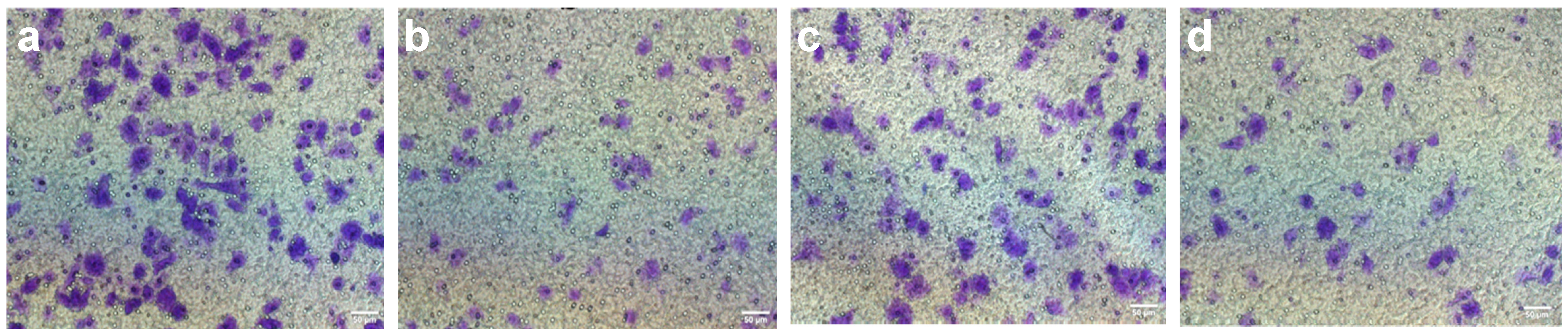
2.2. Inhibition of SGC-7901 Cell Migration and Invasion
| Groups | Dose (nM/mL) | Migration | Invasion | ||
|---|---|---|---|---|---|
| Cell Number | Inhibition (%) | Cell Number | Inhibition (%) | ||
| Control | 0.0 | 153.1 ± 8.74 | 00.00 ± 00.00 | 140.8 ± 7.66 | 00.00 ± 00.00 |
| Strophalloside | 0.12 | 127.4 ± 6.52 * | 16.79 ± 7.42 * | 108.0 ± 5.47 * | 23.30 ± 5.30 * |
| 0.24 | 76.7 ± 7.31 * | 49.90 ± 6.31 * | 64.2 ± 6.06 * | 54.40 ± 4.29 * | |
| 0.47 | 49.3 ± 6.47 * | 67.80 ± 8.47 * | 41.2 ± 5.97 * | 70.74 ± 4.24 * | |
| 0.93 | 30.7 ± 7.54 * | 79.95 ± 5.75 * | 22.4 ± 4.77 * | 84.10 ± 3.39 * | |

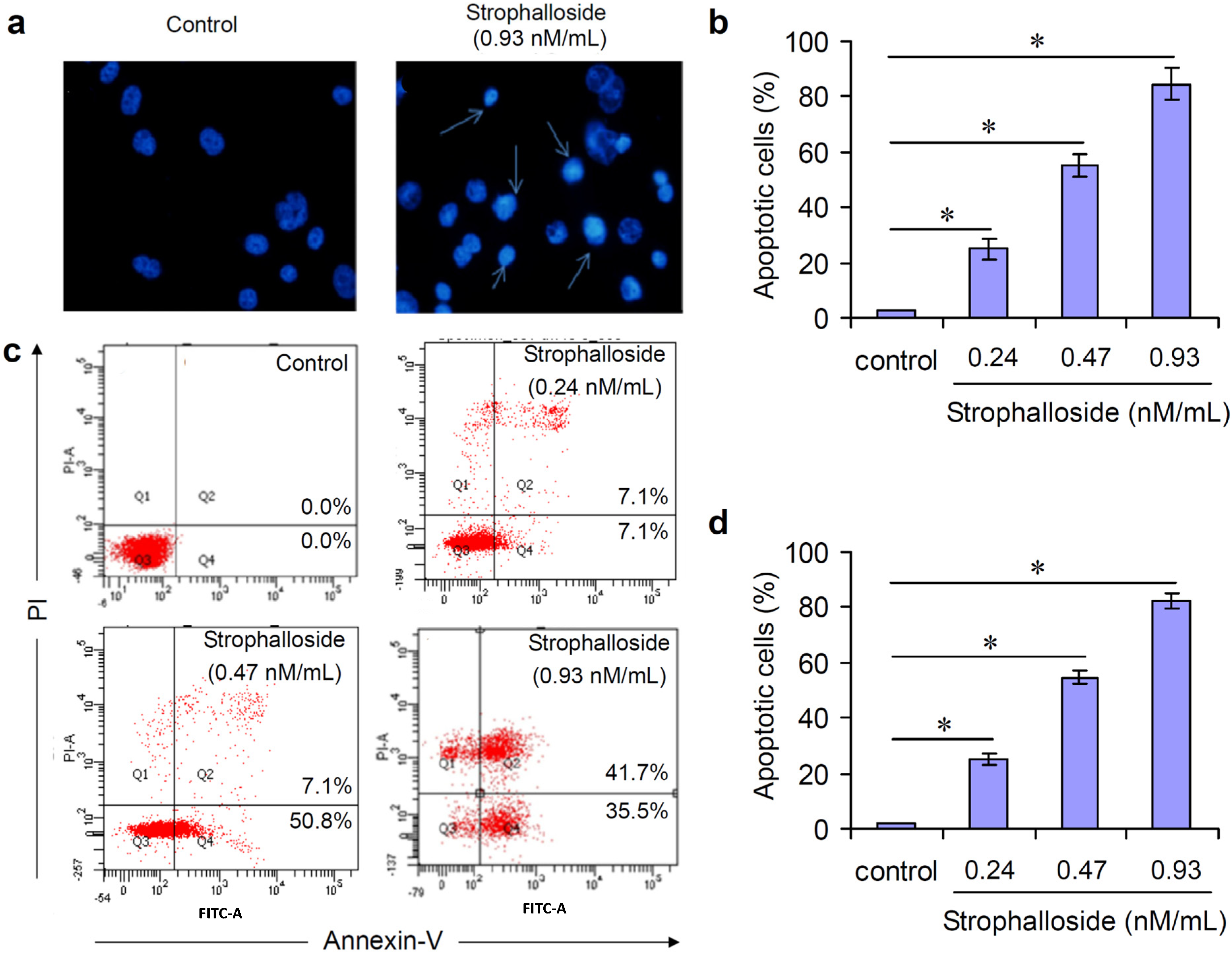
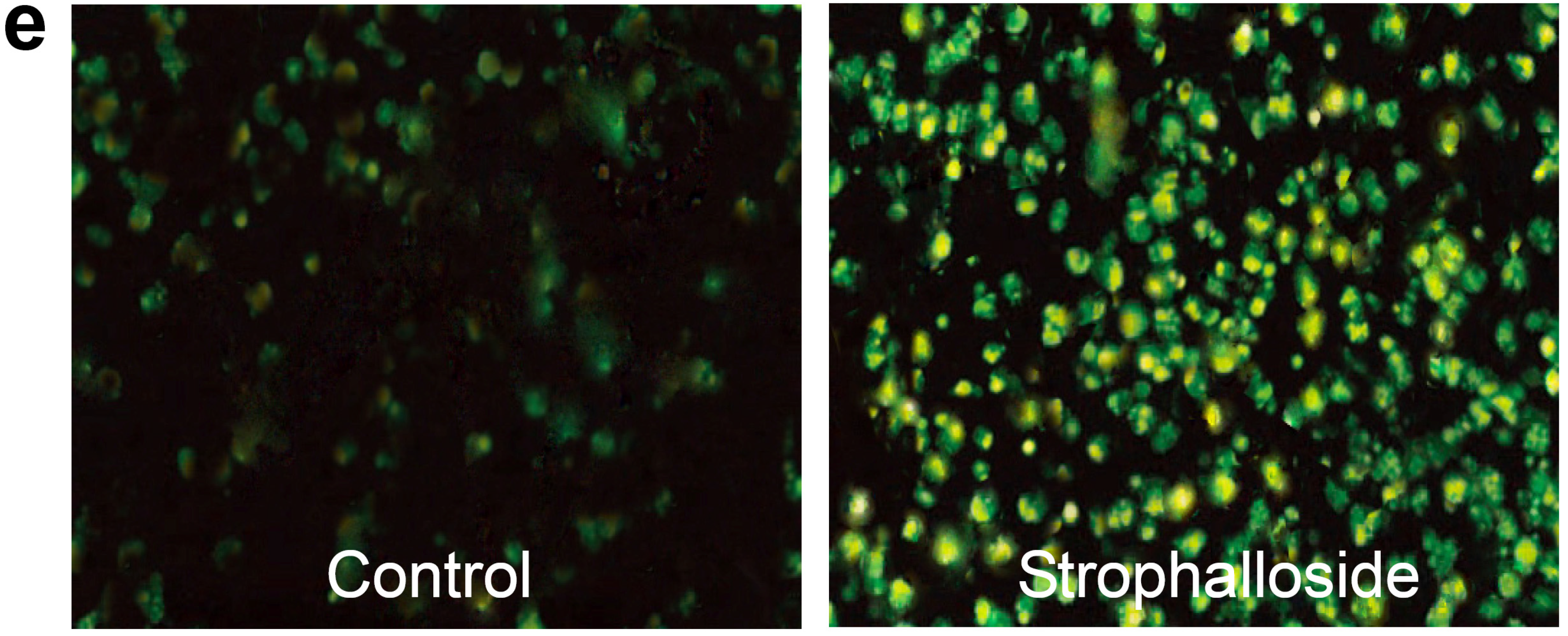
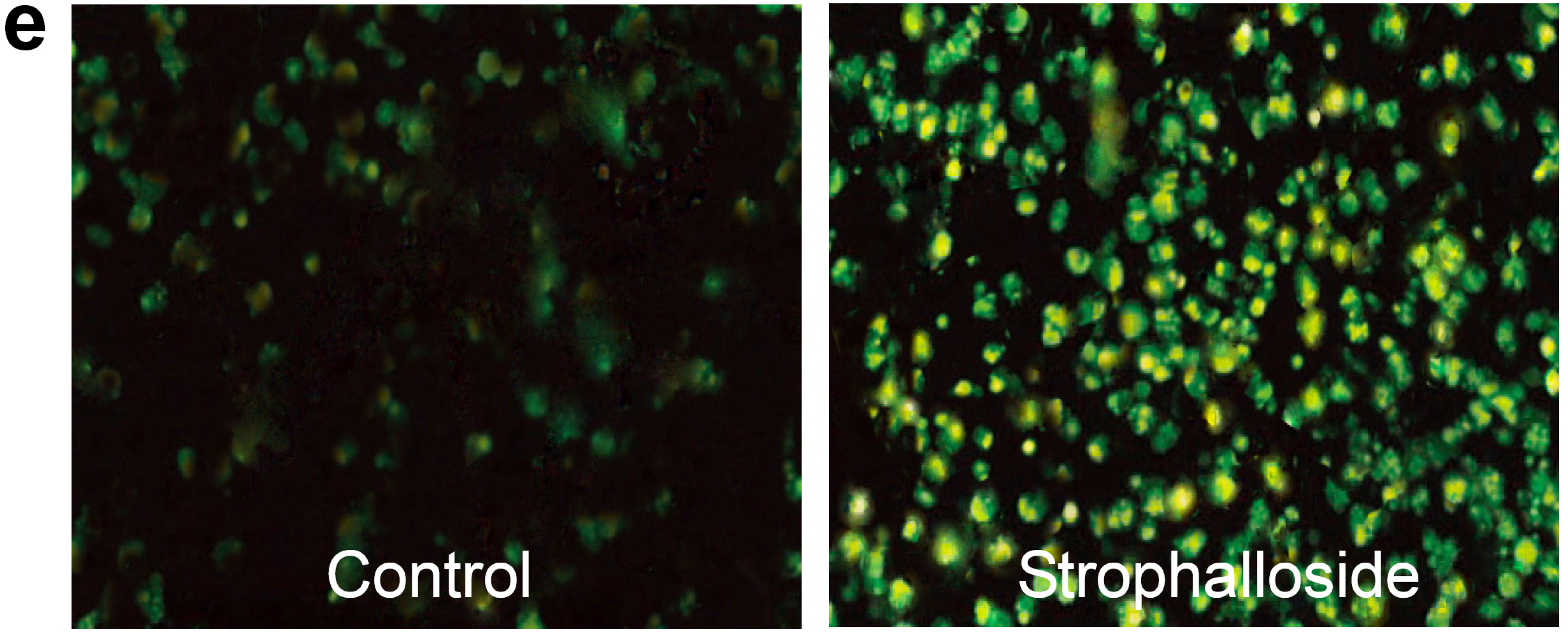
2.3. Strophalloside Induced SGC-7901 Cell Death in Vitro

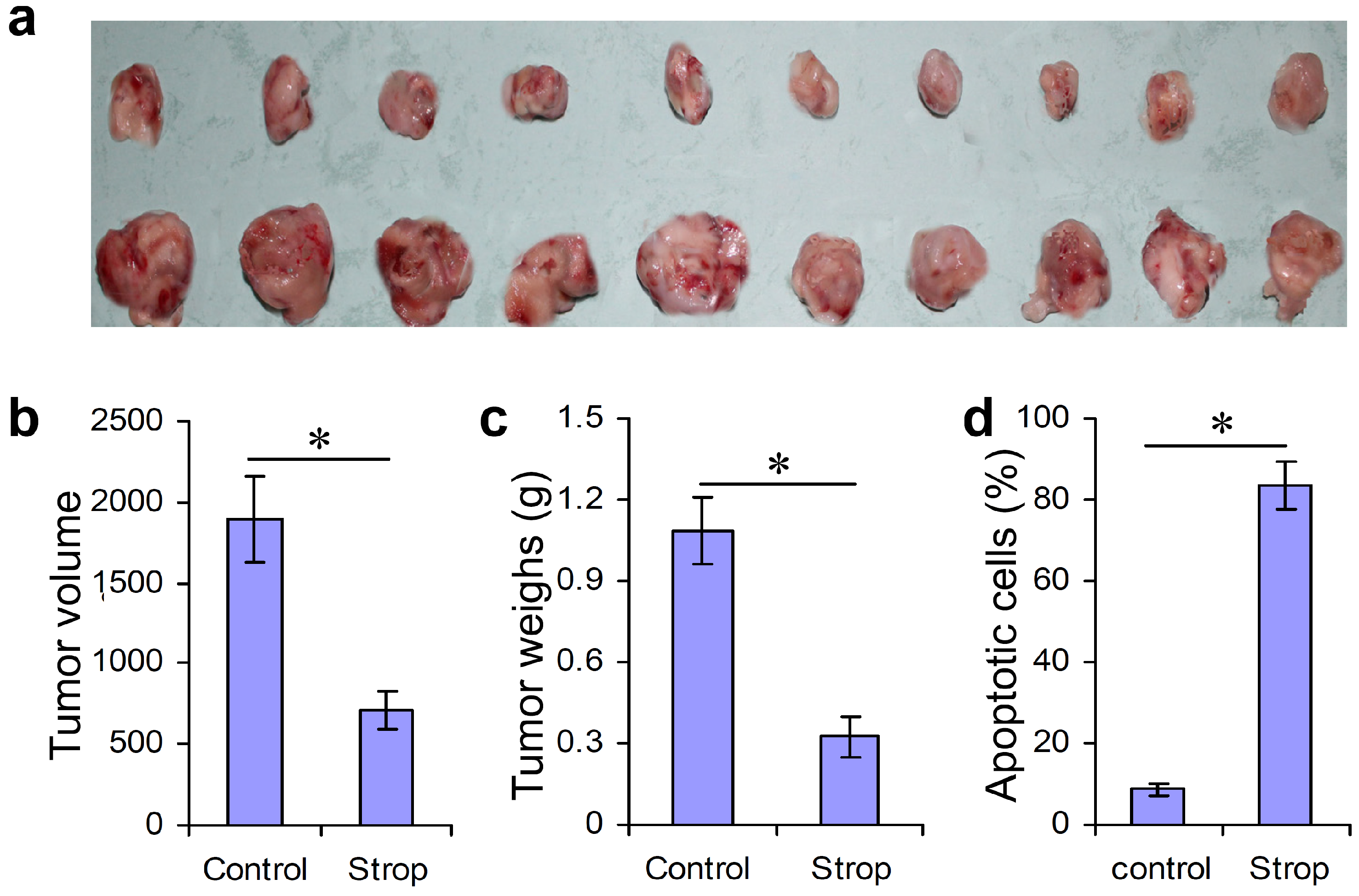
2.4. Strophalloside Inhibited Tumor Growth and Induced SGC-7901 Cell Apoptosis in Vivo

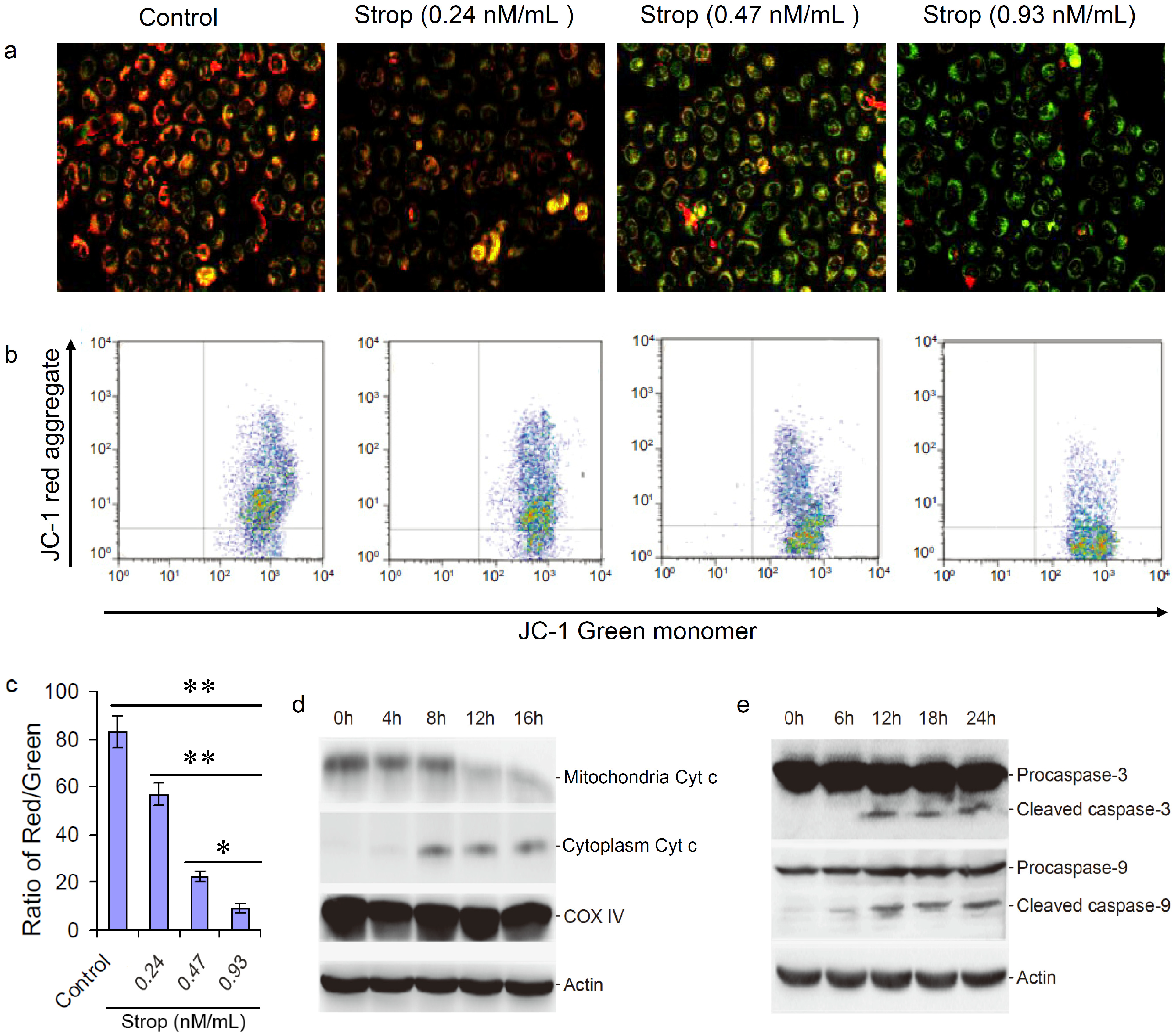
2.5. Strophalloside Activated the Mitochondrion-Mediated Apoptosis Pathway

3. Experimental Section
3.1. Isolation and Purification of Strophalloside
3.2. Cell Culture
3.3. Measurement of Cell Viability
3.4. Invasion and Migration Assay
3.5. Nuclear Morphology Analysis
3.6. Flow Cytometric Analysis of Apoptotic Cell
3.7. Establishment of SGC-7901 Tumor Model and Strophalloside Treatment
3.8. Detection of Tumor Cell Apoptosis in Vivo
3.9. Mitochondrial Membrane Potentials Assay
3.10. Preparation of Cytosolic and Mitochondrial Extracts
3.11. Western Blot Analysis
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.M. A review of complementary and alternative medicine practices among cancer survivors. Clin. J. Oncol. Nurs. 2010, 14, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Lu, J.; Huang, M.; Li, Y.; Chen, M.; Wu, G.; Gong, J.; Zhong, Z.; Xu, Z.; Dang, Y.; et al. Anti-cancer natural products isolated from chinese medicinal herbs. Chin. Med. 2011, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, T.; Kopp, B.; Bisset, N.G. The Moraceae-based dart poisons of South America. Cardiac glycosides of Maquira and Naucleopsis species. J. Ethnopharmacol. 1992, 37, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.F.; Gan, Y.J.; Que, D.M.; Wu, J.; Wen, Z.C.; Mei, W.L. A new cytotoxic 19-nor-cardenolide from the latex of Antiaris toxicaria. Molecules 2009, 14, 3694–3699. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.F.; Gan, Y.J.; Que, D.M.; Wu, J.; Wen, Z.C.; Mei, W.L. Two new cytotoxic cardenolides from the latex of Antiaris toxicaria. J. Asian Nat. Prod. Res. 2009, 11, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.H.; Mei, W.L.; Zhao, Y.X.; Zeng, Y.B.; Wang, H.; Dai, H.F. A new drimane sesquiterpenoid glycoside from the seeds of Antiaris toxicaria. J. Asian Nat. Prod. Res. 2011, 13, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.H.; Mei, W.L.; Zhao, Y.X.; Zeng, Y.B.; Zuo, W.J.; Wang, H.; Li, X.N.; Dai, H.F. Cytotoxic cardenolide glycosides from the seeds of Antiaris toxicaria. Planta Med. 2011, 77, 1730–1734. [Google Scholar] [CrossRef] [PubMed]
- Gheorghiade, M.; van Veldhuisen, D.J.; Colucci, W.S. Contemporary use of digoxin in the management of cardiovascular disorders. Circulation 2006, 113, 2556–2564. [Google Scholar] [CrossRef] [PubMed]
- Hamad, E.; Mather, P.J.; Srinivasan, S.; Rubin, S.; Whellan, D.J.; Feldman, A.M. Pharmacologic therapy of chronic heart failure. Am. J. Cardiovasc. Drugs 2007, 7, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.S.; Liao, Y.R.; Su, M.J.; Lee, A.S.; Kuo, P.C.; Damu, A.G.; Kuo, S.C.; Sun, H.D.; Lee, K.H.; Wu, T.S.; et al. Cardiac glycosides from Antiaris toxicaria with potent cardiotonic activity. J. Nat. Prod. 2010, 73, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- McConkey, D.J.; Lin, Y.; Nutt, L.K.; Ozel, H.Z.; Newman, R.A. Cardiac glycosides stimulate Ca2+ increases and apoptosis in androgen-independent, metastatic human prostate adenocarcinoma cells. Cancer Res. 2000, 60, 3807–3812. [Google Scholar] [PubMed]
- Choedon, T.; Mathan, G.; Arya, S.; Kumar, V.L.; Kumar, V. Anticancer and cytotoxic properties of the latex of Calotropis procera in a transgenic mouse model of hepatocellular carcinoma. World J. Gastroenterol. 2006, 12, 2517–2522. [Google Scholar] [PubMed]
- Frese, S.; Frese-Schaper, M.; Andres, A.C.; Miescher, D.; Zumkehr, B.; Schmid, R.A. Cardiac glycosides initiate Apo2L/TRAIL-induced apoptosis in non-small cell lung cancer cells by up-regulation of death receptors 4 and 5. Cancer Res. 2006, 66, 5867–5874. [Google Scholar] [CrossRef] [PubMed]
- Prassas, I.; Diamandis, E.P. Novel therapeutic applications of cardiac glycosides. Nat. Rev. Drug Discov. 2008, 7, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Denmeade, S.; Carducci, M.A. HIF-1alpha and calcium signaling as targets for treatment of prostate cancer by cardiac glycosides. Curr. Cancer Drug Tar. 2009, 9, 881–887. [Google Scholar] [CrossRef]
- Wang, Z.; Zheng, M.; Li, Z.; Li, R.; Jia, L.; Xiong, X.; Southall, N.; Wang, S.; Xia, M.; Austin, C.P.; et al. Cardiac glycosides inhibit p53 synthesis by a mechanism relieved by Src or MAPK inhibition. Cancer Res. 2009, 69, 6556–6564. [Google Scholar] [CrossRef] [PubMed]
- Juncker, T.; Cerella, C.; Teiten, M.H.; Morceau, F.; Schumacher, M.; Ghelfi, J.; Gaascht, F.; Schnekenburger, M.; Henry, E.; Dicato, M.; et al. UNBS1450, a steroid cardiac glycoside inducing apoptotic cell death in human leukemia cells. Biochem. Pharmacol. 2011, 81, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.L.; Zheng, S.J.; Li, Y.N.; Jie, W.; Hao, X.B.; Li, T.F.; Xia, L.P.; Mei, W.L.; Huang, F.Y.; Kong, Y.Q.; et al. Toxicarioside A inhibits SGC-7901 proliferation, migration and invasion via NF-kappaB/bFGF signaling. World J. Gastroenterol. 2012, 18, 1602–1609. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, V.; Pirker, C.; Martin de Lassalle, E.; Vernier, M.; Mijatovic, T.; DeNeve, N.; Gaussin, J.F.; Dehoux, M.; Lefranc, F.; Berger, W.; et al. The sodium pump alpha1 sub-unit: A disease progression-related target for metastatic melanoma treatment. J. Cell. Mol. Med. 2009, 13, 3960–3972. [Google Scholar] [CrossRef] [PubMed]
- Juncker, T.; Schumacher, M.; Dicato, M.; Diederich, M. UNBS1450 from Calotropis procera as a regulator of signaling pathways involved in proliferation and cell death. Biochem. Pharmacol. 2009, 78, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E.; Heufelder, A.E.; Hengartner, M.O. Advances in apoptosis research. Proc. Natl. Acad. Sci. USA 1997, 94, 12736–12737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradzka, I. Mechanisms and regulation of the programmed cell death. Postepy Biochem. 2006, 52, 157–165. [Google Scholar] [PubMed]
- White, B.C.; Sullivan, J.M. Apoptosis. Acad. Emerg. Med. 1998, 5, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Kim, W.J. Involvement of mitochondrial- and Fas-mediated dual mechanism in CoCl2-induced apoptosis of rat PC12 cells. Neurosci. Lett. 2004, 371, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Orrenius, S. Mitochondrial regulation of apoptotic cell death. Toxicol. Lett. 2004, 149, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Nalepa, G.; Zukowska-Szczechowska, E. Caspases and apoptosis: Die and let live. Wiad Lek 2002, 55, 100–106. [Google Scholar] [PubMed]
- Pirnia, F.; Schneider, E.; Betticher, D.C.; Borner, M.M. Mitomycin C induces apoptosis and caspase-8 and -9 processing through a caspase-3 and Fas-independent pathway. Cell Death Differ. 2002, 9, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Czerski, L.; Nunez, G. Apoptosome formation and caspase activation: Is it different in the heart? J. Mol. Cell. Cardiol. 2004, 37, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Crow, M.T.; Mani, K.; Nam, Y.J.; Kitsis, R.N. The mitochondrial death pathway and cardiac myocyte apoptosis. Circ. Res. 2004, 95, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Youle, R.J. The role of mitochondria in apoptosis. Annu. Rev. Genet. 2009, 43, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Norberg, E.; Orrenius, S.; Zhivotovsky, B. Mitochondrial regulation of cell death: Processing of apoptosis-inducing factor (AIF). Biochem. Biophys. Res. Commun. 2010, 396, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Scorrano, L.; Korsmeyer, S.J. Mechanisms of cytochrome c release by proapoptotic BCL-2 family members. Biochem. Biophys. Res. Commun. 2003, 304, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome C-mediated apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.G.; Choi, E.J. Apoptotic signaling pathways: Caspases and stress-activated protein kinases. J. Biochem. Mol. Biol. 2002, 35, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.Y.; Mei, W.L.; Li, Y.N.; Tan, G.H.; Dai, H.F.; Guo, J.L.; Wang, H.; Huang, Y.H.; Zhao, H.G.; Zhou, S.L.; et al. Toxicarioside A inhibits tumor growth and angiogenesis: Involvement of TGF-beta/endoglin signaling. PLoS ONE 2012, 7, e50351. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.Y.; Mei, W.L.; Tan, G.H.; Dai, H.F.; Li, Y.N.; Guo, J.L.; Huang, Y.H.; Zhao, H.G.; Wang, H.; Zhou, S.L.; et al. Cytochalasin D promotes pulmonary metastasis of B16 melanoma through expression of tissue factor. Oncol. Rep. 2013, 30, 478–484. [Google Scholar] [PubMed]
- Sample Availability: Samples of the strophalloside is available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.-J.; Mei, W.-L.; Tan, G.-H.; Wang, C.-C.; Zhou, S.-L.; Huang, F.-R.; Chen, B.; Dai, H.-F.; Huang, F.-Y. Strophalloside Induces Apoptosis of SGC-7901 Cells through the Mitochondrion-Dependent Caspase-3 Pathway. Molecules 2015, 20, 5714-5728. https://doi.org/10.3390/molecules20045714
Zhang X-J, Mei W-L, Tan G-H, Wang C-C, Zhou S-L, Huang F-R, Chen B, Dai H-F, Huang F-Y. Strophalloside Induces Apoptosis of SGC-7901 Cells through the Mitochondrion-Dependent Caspase-3 Pathway. Molecules. 2015; 20(4):5714-5728. https://doi.org/10.3390/molecules20045714
Chicago/Turabian StyleZhang, Xue-Jiao, Wen-Li Mei, Guang-Hong Tan, Cai-Chun Wang, Song-Lin Zhou, Feng-Ru Huang, Bin Chen, Hao-Fu Dai, and Feng-Ying Huang. 2015. "Strophalloside Induces Apoptosis of SGC-7901 Cells through the Mitochondrion-Dependent Caspase-3 Pathway" Molecules 20, no. 4: 5714-5728. https://doi.org/10.3390/molecules20045714