
Antimycobacterial and Anti-Inflammatory Activities of Substituted Chalcones Focusing on an Anti-Tuberculosis Dual Treatment Approach

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results

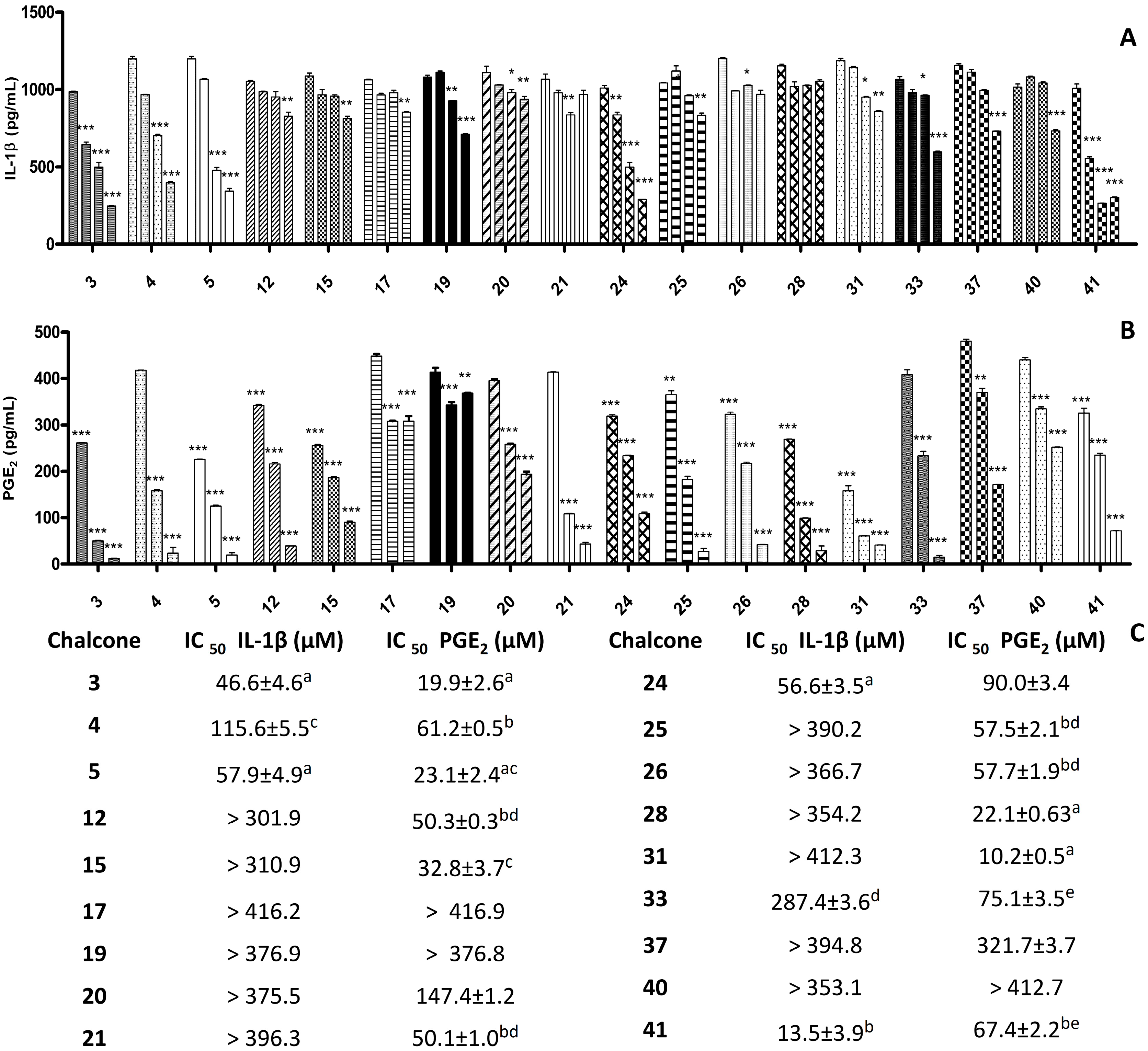{kind=link}
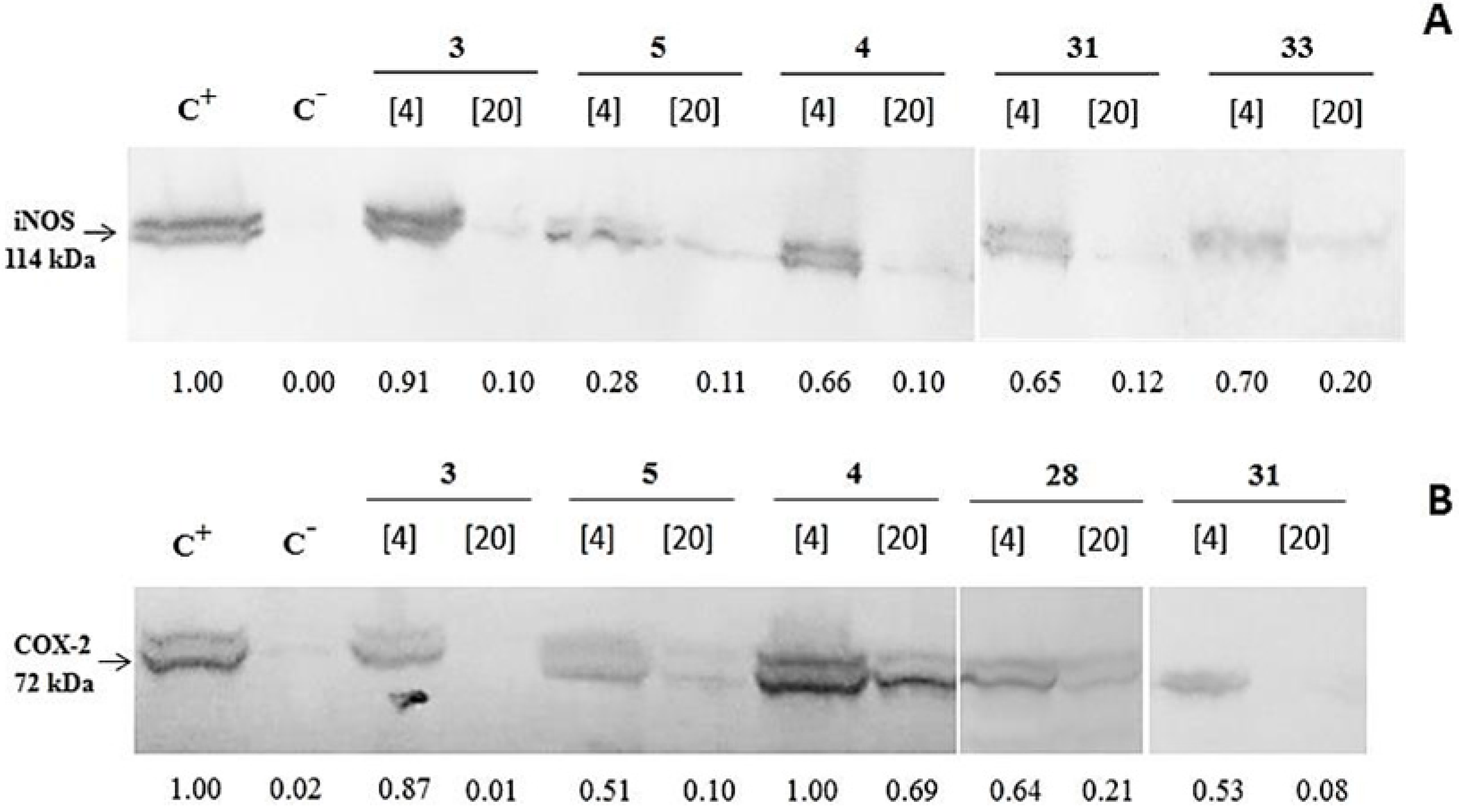{kind=link}
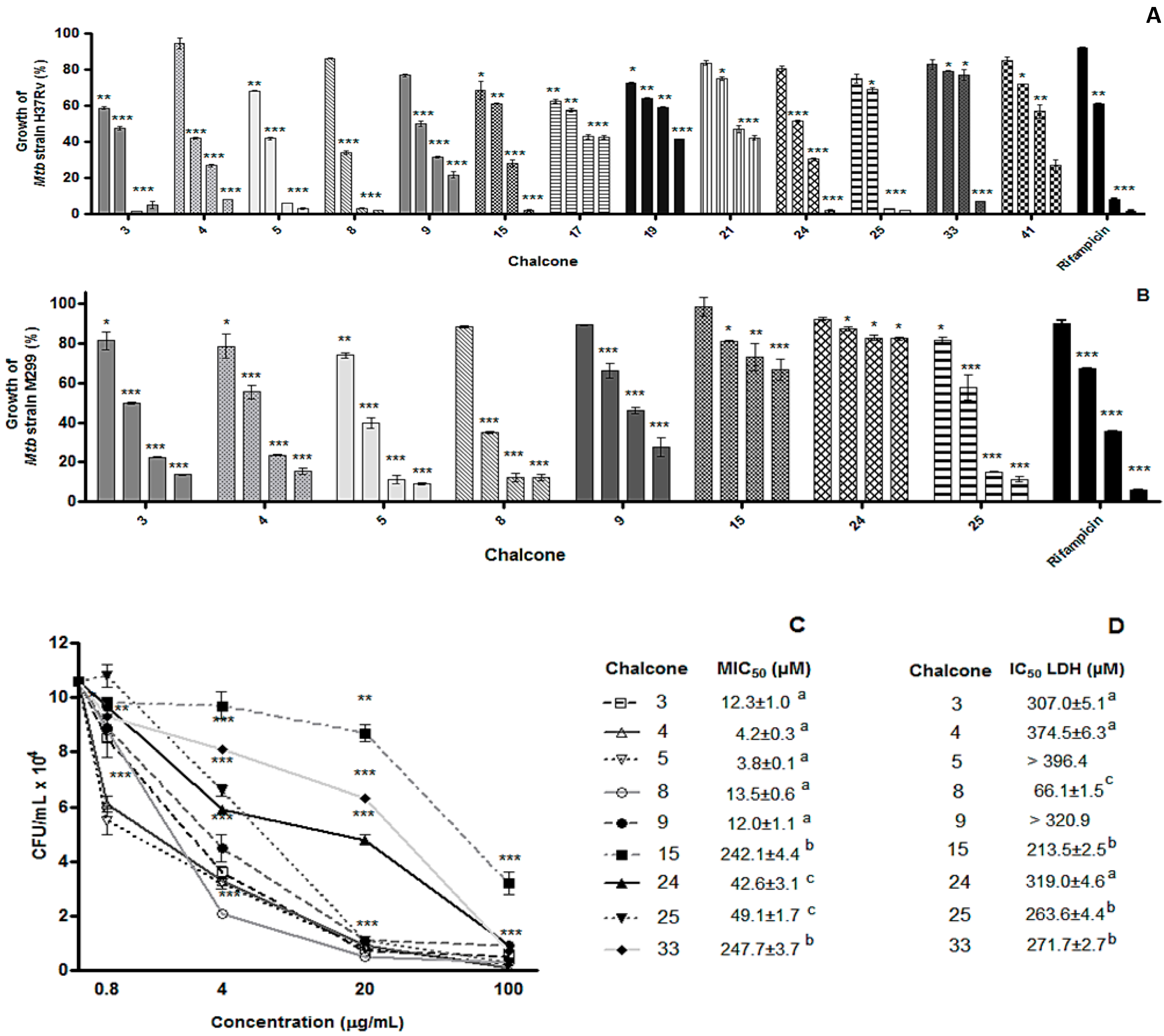{kind=link}

| Chalcone | B Ring | A Ring | ||||
|---|---|---|---|---|---|---|
| 2' | 3' | 4' | 2 | 3 | 4 | |
| 1 | H | H | H | H | H | H |
| 2 | H | H | H | H | H | F |
| 3 | H | H | H | H | H | Cl |
| 4 | H | H | H | H | H | N(CH3)2 |
| 5 | H | H | H | H | -O(CH2)O- | |
| 6 | H | H | H | H | H | OCH3 |
| 7 | H | H | H | N (pyridine ring) | H | H |
| 8 | H | H | Br | H | H | H |
| 9 | H | H | Br | H | H | F |
| 10 | H | H | Br | H | H | Cl |
| 11 | H | H | Br | H | H | N(CH3)2 |
| 12 | H | H | Br | H | -O(CH2)O- | |
| 13 | H | H | Br | H | H | OCH3 |
| 14 | H | H | Br | H | OCH3 | OCH3 |
| 15 | H | H | Br | Cl | H | H |
| 16 | H | H | CH3 | H | H | H |
| 17 | H | H | CH3 | H | H | F |
| 18 | H | H | CH3 | H | H | Cl |
| 19 | H | H | CH3 | H | H | N(CH3)2 |
| 20 | H | H | CH3 | H | -O(CH2)O- | |
| 21 | H | H | CH3 | H | H | OCH3 |
| 22 | H | H | CH3 | H | OCH3 | OCH3 |
| 23 | H | H | CH3 | Cl | H | H |
| 24 | H | H | OCH3 | H | H | H |
| 25 | H | H | OCH3 | H | H | F |
| 26 | H | H | OCH3 | H | H | Cl |
| 27 | H | H | OCH3 | H | H | N(CH3)2 |
| 28 | H | H | OCH3 | H | -O(CH2)O- | |
| 29 | H | H | OCH3 | H | H | OCH3 |
| 30 | H | H | OCH3 | Cl | H | H |
| 31 | Cl | H | H | H | H | H |
| 32 | Cl | H | H | H | H | F |
| 33 | Cl | H | H | H | H | Cl |
| 34 | Cl | H | H | H | H | OCH3 |
| 35 | Cl | H | H | H | -O(CH2)O- | |
| 36 | Cl | H | H | H | OCH3 | OCH3 |
| 37 | H | H | H | H | H | NO2 |
| 38 | H | H | Br | H | H | NO2 |
| 39 | H | H | CH3 | H | H | NO2 |
| 40 | H | H | OCH3 | H | H | NO2 |
| 41 | Cl | H | H | H | H | NO2 |
| Chalcone | IC50 (µM) | MIC50 (µM) | MIC90 (µM) | ||
|---|---|---|---|---|---|
| NO | TNF-α | LDH | M. bovis BCG | ||
| 1 | 41.6 ± 2.0 | >480.2 | 177.3 ± 9.5 | 137.3 ± 4.7 | 383.2 ± 9.1 |
| 2 | >442.0 | >442.0 | >442.0 | >442.0 | >442.0 |
| 3 | 31.9 ± 4.6 | 60.5 ± 4.7 | 399.7 ± 4.9 | 20.7 ± 6.4 | 169.9 ± 5.4 |
| 4 | 12.0 ± 2.2 | 66.9 ± 4.3 | 348.4 ± 5.1 | 14.3 ± 5.8 | 137.3 ± 7.9 |
| 5 | 2.1 ± 2.7 | 48.1 ± 7.6 | >396.4 | 14.5 ± 6.1 | 75.3 ± 8.4 |
| 6 | >419.7 | >419.7 | >419.7 | 255.9 ± 4.8 | 249.2 ± 6.2 |
| 7 | >477.9 | >477.9 | >477.9 | 35.8 ± 7.2 | 440.8 ± 9.5 |
| 8 | 46.4 ± 2.3 | 66.9 ± 4.5 | 191.0 ± 5.2 | 13.4 ± 4.0 | 33.7 ± 4.6 |
| 9 | >327.7 | >327.7 | >327.7 | 35.8 ± 3.9 | 308.8 ± 5.3 |
| 10 | 68.9 ± 4.3 | >310.9 | 113.4 ± 0.5 | 22.3 ± 6.3 | 287.8 ± 3.2 |
| 11 | 41.3 ± 3.5 | >302.8 | 132.0 ± 2.9 | 67.8 ± 3.8 | 242.1 ± 2.6 |
| 12 | 19.6 ± 3.3 | >302.0 | >302.0 | 84.9 ± 4.7 | 229.6 ± 2.2 |
| 13 | 52.9 ± 3.6 | >315.3 | >315.3 | 220.6 ± 3.6 | >315.3 |
| 14 | 58.9 ± 6.8 | >288.0 | >288.0 | >288.0 | >288.0 |
| 15 | 36.7+3.8 | >311.0 | 256.0 ± 3.8 | 13.8 ± 4.4 | 265.1 ± 4.5 |
| 16 | >449.9 | >449.9 | >449.9 | >449.9 | >449.9 |
| 17 | 30.8 ± 4.5 | >416.2 | 289.5 ± 4.3 | 61.5 ± 7.6 | 388.4 ± 7.3 |
| 18 | 150.3 ± 3.8 | >389.5 | 304.3 ± 4.2 | 71.3 ± 4.8 | 323.2 ± 7.1 |
| 19 | 27.8 ± 3.8 | >376.9 | >376.9 | 56.4 ± 4.4 | 322.7 ± 1.7 |
| 20 | 30.1 ± 1.2 | >375.5 | >375.5 | >375.5 | >375.5 |
| 21 | 30.6 ± 4.7 | >396.3 | >396.3 | 33.3 ± 5.4 | 362.2 ± 5.9 |
| 22 | >354.2 | >354.2 | >354.2 | >354.2 | >354.2 |
| 23 | >389.5 | >389.5 | >389.5 | >389.5 | >389.5 |
| 24 | 21.3 ± 4.5 | 78.8 ± 6.5 | 244.8 ± 4.5 | 31.5 ± 5.9 | 399.1 ± 6.9 |
| 25 | 41.2 ± 4.6 | >390.2 | >390.2 | 15.8 ± 5.2 | 228.3 ± 5.8 |
| 26 | 25.3 ± 3.9 | >366.7 | >366.7 | 91.2 ± 5.0 | 286.7 ± 7.8 |
| 27 | 44.6 ± 5.2 | >355.4 | 226.4 ± 3.5 | 308.8 ± 5.0 | >355.4 |
| 28 | 14.9 ± 4.1 | >354.2 | >354.2 | >354.2 | >354.2 |
| 29 | 13.1 ± 4.2 | >372.7 | 93.9 ± 4.9 | 32.5 ± 5.4 | 339.5 ± 3.2 |
| 30 | 23.9 ± 4.8 | >366.7 | 100.0 ± 5.1 | 37.8 ± 6.4 | 328.7 ± 3.6 |
| 31 | 8.4 ± 6.8 | >412,0 | 267.0 ± 4.5 | 90.9 ± 5.5 | 330.2 ± 4.2 |
| 32 | >383.6 | >383.6 | >383.6 | >383.6 | >383.6 |
| 33 | 3.1 ± 0.1 | 165.5 ± 5.2 | 351.2 ± 5.0 | 45.7 ± 5.4 | >360.8 |
| 34 | >366.7 | >366.7 | >366.7 | >366.7 | >366.7 |
| 35 | 160.3 ± 4.7 | >348.8 | >348.8 | >348.8 | >348.8 |
| 36 | 187.8 ± 4.1 | >330.3 | >330.3 | 269.6 ± 3.9 | >330.3 |
| 37 | 58.6 ± 4.5 | >394.9 | >394.9 | >394.9 | >394.9 |
| 38 | >301.1 | >301.1 | >301.1 | 97.4 ± 4.2 | 220.7 ± 5.6 |
| 39 | 113.7 ± 4.8 | >373.2 | 328.6 ± 3.9 | 28.1 ± 5.4 | 344.1 ± 6.3 |
| 40 | 13.5 ± 5.5 | >353.0 | >353.0 | 14.5 ± 3.1 | >353.0 |
| 41 | 11.2 ± 5.0 | 68.54 ± 3.3 | >347.6 | 30.5 ± 4.7 | 246.9 ± 7.2 |
| L-NMMA 1 | 78.3 ± 6.5 | XX | XX | XX | XX |
| Rifampicin 2 | XX | XX | XX | 0.01 ± 0.03 | 0.2 ± 0.01 |


| Chalcone | MIC50 (µM) | MIC90 (µM) | MIC50 (µM) | MIC90 (µM) |
|---|---|---|---|---|
| M. tuberculosis H37Rv | M. tuberculosis M299 | |||
| 3 | 13.0 ± 3.3 a | 372.7 ± 3.6 a | 18.37 ± 3.9 a | 373.9 ± 1.4 |
| 4 | 19.7 ± 1.9 a | 320.1 ± 4.8 b | 20.1 ± 1.0 a | 351.5 ± 1.8 |
| 5 | 12.7 ± 1.9 a | 303.2 ± 5.4 | 11.7 ± 3.4 a | 289.6 ± 0.7 |
| 7 | >477.9 | >477.9 | XX | XX |
| 8 | 10.5 ± 3.8 a | 325.9 ± 3.3 b | 10.2 ± 4.9 a | 344.7 ± 0.4 |
| 9 | 24.4 ± 4.5 a | 252.1 ± 2.7 c | 53.58 ± 1.9 | 313.0 ± 1.4 |
| 15 | 17.1 ± 3.8 a | 287.8 ± 5.5 c | >311.0 | >311.0 |
| 17 | 63.8 ± 6.2 b | >416.2 | XX | XX |
| 19 | 112.4 ± 6.4 c | >376.8 | XX | XX |
| 21 | 66.9 ± 6.2 b | 368.2 ± 5.2 a | XX | XX |
| 24 | 25.3 ± 5.2 a | 374.6 ± 6.8 a | >419.7 | >419.7 |
| 25 | 25.1 ± 5.2 a | 366.1 ± 3.6 a | 18.5 ± 3.6 a | 366.9 ± 0.3 |
| 33 | 36.9 ± 4.5 a | 355.8 ± 4.9 a | XX | XX |
| 39 | >373.2 | >373.2 | XX | XX |
| 40 | 131.7 ± 1.3 c | 351.8 ± 5.9 a | XX | XX |
| 41 | 82.1 ± 4.3 b | 275.3 ± 6.7 c | XX | XX |
| Rifampicin | 0.11 ± 0.02 | 0.15 ± 0.08 | 0.2 ± 0.18 | 3.3 ± 0.16 |


| # | O1 | C7 | C8 | C9 | C8, C7, C2, C3 | C8, C9, C11, C14 |
|---|---|---|---|---|---|---|
| 3 | −0.591 | 0.583 | −0.307 | −0.169 | 38.51 | −48.75 |
| 4 | −0.608 | 0.645 | −0.434 | −0.079 | 31.08 | 27.78 |
| 5 | −0.591 | 0.564 | −0.405 | −0.036 | −38.86 | 46.97 |
| 8 | −0.589 | 0.610 | −0.364 | −0.184 | 39.72 | −47.79 |
| 9 | −0.586 | 0.582 | −0.362 | −0.142 | 39.81 | −47.43 |
| 15 | −0.581 | 0.562 | −0.314 | −0.116 | 39.27 | −54.30 |
| 17 | −0.606 | 0.633 | −0.349 | −0.129 | 38.56 | −47.76 |
| 19 | −0.617 | 0.646 | −0.385 | −0.113 | 41.46 | −39.66 |
| 21 | −0.614 | 0.646 | −0.377 | −0.103 | 40.34 | −43.67 |
| 24 | −0.613 | 0.651 | −0.32 | −0.195 | 36.6 | −48.40 |
| 25 | −0.613 | 0.650 | −0.341 | −0.148 | 36.55 | −48.18 |
| 33 | −0.593 | 0.654 | −0.374 | −0.124 | 53.44 | −45.43 |
| 40 | −0.603 | 0.66 | −0.289 | −0.233 | 35.21 | −49.64 |
| 41 | −0.585 | 0.659 | −0.319 | −0.199 | −48.82 | 48.58 |
| Chalcone | MIC50 (µM) | MW | LogP | HBD | HBA |
|---|---|---|---|---|---|
| 3 | 13.0 ± 3.3 a | 242.70 | 4.32 | 0 | 1 |
| 4 | 19.7 ± 1.9 a | 251.33 | 4.04 | 0 | 2 |
| 5 | 12.7 ± 1.9 a | 252.27 | 3.54 | 0 | 3 |
| 8 | 10.5 ± 3.8 a | 287.16 | 4.59 | 0 | 1 |
| 9 | 24.4 ± 4.5 a | 305.15 | 4.75 | 0 | 1 |
| 15 | 17.1 ± 3.8 a | 321.60 | 5.15 | 0 | 1 |
| 17 | 63.8 ± 6.2 b | 240.28 | 4.4 | 0 | 1 |
| 19 | 112.4 ± 6.4 c | 265.36 | 4.53 | 0 | 2 |
| 21 | 66.9 ± 6.2 b | 252.31 | 4.12 | 0 | 2 |
| 24 | 25.3 ± 5.2 a | 238.29 | 3.63 | 0 | 2 |
| 25 | 25.1 ± 5.2 a | 256.28 | 3.79 | 0 | 2 |
| 33 | 36.9 ± 4.5 a | 277.14 | 4.88 | 0 | 1 |
| 40 | 131.7 ± 1.3 c | 283.28 | 3.67 | 0 | 5 |
| 41 | 82.1 ± 4.3 b | 287.70 | 4.35 | 0 | 4 |
2.2. Discussion
3. Experimental Section
3.1. Reagents
3.2. Synthesis of Substituted Chalcones
Characterization of the Chalcones
3.3. Cell Culture and Treatments
3.4. Quantification of Proinflammatory Mediators (TNF-α, IL-1β, PGE2 and NO)
3.5. Macrophage Cytotoxicity Assay
3.6. Detection of iNOS and COX-2 by Western Blot
3.7. Mycobacterial Culture and Evaluation of Bacterial Growth
3.8. Infection of Macrophage Cultures and Quantification of Intracellular Growth
3.9. Molecular Modelling and in Silico ADMET Studies
3.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Duque, G.A.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [PubMed]
- Medzhitov, R. Origin and physiological roles of inflammation. Nat. Rev. 2008, 454, 428–435. [Google Scholar]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Ashley, N.T.; Zachary, M.W.; Nelson, R.J. Inflammation: Mechanisms, costs, and natural variation. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 385–406. [Google Scholar] [CrossRef]
- Thwaites, G.; Fisher, M.; Hemingway, C.; Scott, G.; Solomon, T.; Innes, J.Innes. British Infection Society guidelines for the diagnosis and treatment of tuberculosis of the central nervous system in adults and children. J. Infect. 2009, 59, 167–187. [Google Scholar] [CrossRef] [PubMed]
- Meintjes, G.; Wilkinson, R.J.; Morroni, C.; Pepper, D.J.; Rebe, K.; Rangaka, M.X.; Oni, T.; Maartens, G. Randomized placebo-controlled trial of prednisone for paradoxical tuberculosis-associated immune reconstitution inflammatory syndrome. AIDS 2010, 24, 2381–2390. [Google Scholar] [PubMed]
- Tobin, D.M.; Roca, F.J.; Oh, S.F.; McFarland, R.; Vickery, T.W.; Ray, J.P.; Ko, D.C.; Zou, Y.; Bang, N.D.; Chau, T.T.; et al. Host genotype-specific therapies can optimize the inflammatory response to mycobacterial infections. Cell 2012, 148, 434–446. [Google Scholar] [CrossRef] [PubMed]
- Critchley, J.A.; Young, F.; Orton, L.; Garner, P. Corticosteroids for prevention of mortality in people with tuberculosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Cholo, M.C.; Steel, H.C.; Fourie, P.B.; Germishuizen, W.A.; Anderson, R. Clofazimine: Current status and future prospects. J. Antimicrob. Chemother. 2012, 67, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Jantan, I.; Bukhari, S.N.; Adekoya, O.A.; Sylte, I. Studies of synthetic chalcone derivatives as potential inhibitors of secretory phospholipase A2, cyclooxygenases, lipoxygenase and pro-inflammatory cytokines. Drug Des. Dev. Ther. 2014, 16, 1405–1418. [Google Scholar] [CrossRef]
- Nowakowska, Z. A review of anti-infective and anti-inflammatory chalcones. Eur. J. Med. Chem. 2007, 42, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, S.N.A.; Jantan, I.; Jasamai, M. Anti-inflammatory trends of 1,3-diphenyl-2-propen-1-one derivatives. Mini Rev. Med. Chem. 2013, 13, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.; Javed, K.; Ahmad, S.; Rathish, I.G.; Singh, S.; Chaitanya, M.; Arunasree, K.M.; Alama, M.S. Synthesis of some novel chalcones, flavanones and flavones and evaluation of their anti-inflammatory activity. Eur. J. Med. Chem. 2013, 65, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-M.; Zhou, Y.; Flavin, M.T.; Zhou, L.-M.; Niea, W.; Chen, F.-C. Chalcones and flavonoids as anti-tuberculosis agents. Bioorg. Med. Chem. 2002, 10, 2795–2802. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A. Chalcone: A valuable insight into the recent advances and potential pharmacological activities. Chem. Sci. J. 2011, 2011, 1–16. [Google Scholar]
- Moodley, S.; Koorbanally, N.A.; Moodley, T.; Ramjugernath, D.; Pillay, M. The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay is a rapid, cheap, screening test for the in vitro anti-tuberculous activity of chalcones. J. Microbiol. Methods 2014, 104, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, P.M.; Kumar, V.; Seenivasan, S.P.; Mohanapriya, J.; Doble, M. Experimental and theoretical approaches to enhance anti tubercular activity of chalcones. WSEAS Trans. Biol. Biomed. 2010, 2, 51–61. [Google Scholar]
- Ávila, H.P.; Fátima, E.; Smânia, A.; Monache, F.D.; Smânia, A., Jr. Structure-activity relationship of antibacterial chalcones. Bioorg. Med. Chem. 2008, 16, 9790–9794. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, P.M.; Seenivasan, S.P.; Kumar, V.; Doble, M. Synthesis, antimycobacterial activity evaluation, and QSAR studies of chalcone derivatives. Bioorg. Med. Chem. Lett. 2007, 17, 1695–1700. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.C.M.; Gomes, L.L.; Amaral, E.P.; Andrade, M.R.M.; Almeida, F.M.; Rezende, A.L.; Lanes, V.R.; Carvalho, E.C.Q.; Suffys, P.N.; Mokrousov, I.; et al. Mycobacterium tuberculosis strains of the modern sublineage of the Beijing family are more likely to display increased virulence than strains of the ancient sublineage. J. Clin. Microbiol. 2014, 52, 2615–2624. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Van de Waterbeemd, H.; Gifford, E. ADMET in silico modelling: Towards prediction paradise? Nat. Rev. Drug Discov. 2003, 2, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Bourré-Tessier, J.; Arino-Torregrosa, M.; Choquette, D. Increased incidence of liver enzymes abnormalities in patients treated with isoniazid in combination with disease modifying and/or biologic agents. Clin. Rheumatol. 2014, 33, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Garlanda, C.; Di Liberto, D.; Annunciata, V.; La Manna, M.P.; Buracchi, C.; Caccamo, N.; Salerno, A.; Dieli, F.; Mantovan, A. Damping excessive inflammation and tissue damage in Mycobacterium tuberculosis infection by toll IL-1 receptor 8/single Ig IL-1-related receptor, a negative regulator of IL-1/TLR signaling. J. Immunol. 2007, 179, 3119–3125. [Google Scholar] [CrossRef] [PubMed]
- Buonsenso, D.; Serranti, D.; Valentini, P. Management of central nervous system tuberculosis in children: Light and shade. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 845–853. [Google Scholar] [PubMed]
- Prasad, K.; Singh, M.B. Corticosteroids for managing tuberculous meningitis. Cochrane Database Syst. Rev. 2010, 23. [Google Scholar] [CrossRef]
- Gao, X.; He, J.; Wu, X.-D.; Peng, L.-Y.; Dong, L.-B.; Deng, X.; Li, Y.; Cheng, X.; Zhao, Q.-S. Further lignans from Saururus chinensis. Planta Med. 2014, 79, 1720–1723. [Google Scholar]
- Lee, J.Y.; Park, W. Anti-inflammatory effect of myristicin on RAW 264.7 macrophages stimulated with polyinosinic-polycytidylic acid. Molecules 2011, 16, 7132–7142. [Google Scholar] [CrossRef] [PubMed]
- Kontogiorgis, C.; Mantzanidou, M.; Hadjipavlou-Litina, D. Chalcones and their potential role in inflammation. Mini Rev. Med. Chem. 2008, 8, 1224–1242. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.A.; Andrade, C.K.Z.; Napolitano, H.B.; Vencato, I.; Lariucci, C.; de Castro, M.R.C.; Camargo, A.J. Biological and structure-activity evaluation of chalcone derivatives against bacteria and fungi. J. Braz. Chem. Soc. 2013, 24, 133–144. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, J.; Park, H.; Kim, H.P. Anti-inflammatory activity of the synthetic chalcone derivatives: Inhibition of inducible nitric oxide synthase-catalyzed nitric oxide production from lipopolysaccharide-treated RAW 264.7 cells. Biol. Pharm. Bull. 2007, 30, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Downs, L.E.; Wolfe, D.M.; Schreiner, P.R. Organic base-mediated condensation of ppridinecarboxaldehydes to azachalcones. Adv. Synth. Catal. 2005, 347, 235–238. [Google Scholar] [CrossRef]
- Hsieh, C.T.; Hsieh, T.J.; El-Shazly, M.; Chuang, D.W.; Tsai, Y.H.; Yen, C.T.; Wu, S.F.; Wu, Y.C.; Chang, F.R. Synthesis of chalcone derivatives as potential anti-diabetic agents. Bioorg. Med. Chem. Lett. 2012, 22, 3912–3915. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, G.; Veena, K.; Narayana, B.; Yathirajan, H.S.; Kubicki, M. (2E)-1-(4-Bromophenyl)-3-(4-fluorophenyl)prop-2-en-1-one. Acta Crystallogr. Sect. E: Struct. Rep. Online 2010, 66, o1243–o1244. [Google Scholar] [CrossRef]
- Ahirwar, M.K.; Shrivastava, S.P.; Mehta, P. Synthesis and antimicrobial activity of some pyrazoline derivatives. J. Pharm. Chem. 2010, 4, 75–79. [Google Scholar]
- Patil, P.S.; Chantrapromma, S.; Fun, H.-K.; Dharmaprakash, S.M. 1-(3-Bromophenyl)-3-[4-(dimethylamino)phenyl]prop-2-en-1-one. Acta Crystallogr. Sect. E: Struct. Rep. Online 2007, 63, o1738–o1740. [Google Scholar] [CrossRef]
- Vital, D.G.; Arribas, M.; Trossini, G.H.G. Molecular modeling and docking application to evaluate cruzain inhibitory activity by chalcones and hydrazides. Lett. Drug Des. Discov. 2014, 11, 249–255. [Google Scholar] [CrossRef]
- Hayat, F.; Salahuddin, A.; Umar, S.; Azam, A. Synthesis, characterization, antiamoebic activity and cytotoxicity of novel series of pyrazoline derivatives bearing quinoline tail. Eur. J. Med. Chem. 2010, 45, 4669–4675. [Google Scholar] [CrossRef] [PubMed]
- Joseph, L.; Arunsasi, B.S.; Sajan, D.; Shettigar, V. Synthesis, crystal growth, thermal, electronic and vibrational spectral studies of 1-(4-bromophenyl)-3-(3,4-dimethoxyphenyl)prop-2-en-1-one: A density functional theory study. J. Mol. Struct. 2014, 1076, 687–697. [Google Scholar] [CrossRef]
- Kistayya, C.; Rao, N.G.R.; Rao, A.V.; Srinath, N.; Nayak, B.S.; Chawda, B. Synthesis and biological evaluation of some new chalcones as potential cytotoxic agents. Int. J. Pharm. 2014, 4, 397–406. [Google Scholar]
- Schmink, J.R.; Holcomb, J.L.; Leadbeater, N.E. Testing the validity of microwave-interfaced, in situ raman spectroscopy as a tool for kinetic studies. Org. Lett. 2009, 11, 365–368. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-F.; Neumann, H.; Spannenberg, A.; Schulz, T.; Jiao, H.; Beller, M. Development of a general palladium-catalyzed carbonylative heck reaction of aryl halides. J. Am. Chem. Soc. 2010, 132, 14596–14602. [Google Scholar] [CrossRef] [PubMed]
- Alegaon, S.G.; Alagawadi, K.R.; Dadwe, D.H. Synthesis and anti-tubercular activity of novel 3, 5-diaryl-4,5-dihydro-1H-pyrazole derivatives. Drug Res. 2014, 64, 553–558. [Google Scholar] [CrossRef]
- Sonmez, F.; Sevmezler, S.; Atahan, A.; Ceylan, M.; Demir, D.; Gencer, N.; Arslan, O.; Kucukislamoglu, M. Evaluation of new chalcone derivatives as polyphenol oxidase inhibitors. Bioorg. Med. Chem. Lett. 2011, 21, 7479–7482. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Kumari, N.; Joshi, Y.C.; Joshi, P.; Shekhawat, S.S. Solvent free microwave assisted synthesis of chalcones and their antifungal activities. Asian J. Chem. 2007, 19, 4483–4486. [Google Scholar]
- Lopez, S.; Castelli, M.V.; Zacchino, S.; Dominguez, J.N.; Lobo, G.; Charris-Charris, J.; Coetrs, J.C.C.; Ribas, J.C.; Devia, C.; Rodrigues, A.M.; et al. In vitro antifungal evaluation and structure-activity relationships of a new series of chalcone derivatives and synthetic analogues, with inhibitory properties against polymers of the fungal cell wall. Bioorg. Med. Chem. 2001, 9, 1999–2013. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Malik, O.P. Synthesis and antimicrobial activity of 1-(4-chlorophenyl)-3(4-methoxy/3,4-dimethoxyphenyl)propan-1,3-diones and their 2-[(2-nitro/4-chloro or bromophenyl) azo]analogs. Indian J. Chem. 1994, 34, 455–459. [Google Scholar]
- Ananthnag, G.S.; Adhikari, A.; Balakrishna, M.S. Iron-catalyzed aerobic oxidative aromatization of 1,3,5-trisubstituted pyrazolines. Catal. Commun. 2014, 43, 240–243. [Google Scholar] [CrossRef]
- Sharma, N.; Sharma, A.; Kumar, R.; Shard, A.; Sinha, A.K. One-pot two-step oxidative cleavage of 1,2-arylalkenes to aryl ketones instead of arylaldehydes in an aqueous medium: A complementary approach to ozonolysis. Eur. J. Org. Chem. 2010, 31, 6025–6032. [Google Scholar] [CrossRef]
- Mitch, C.H.; Quimby, S.J.; Diaz, N.; Pedregal, C.; Torre, M.G.; Jimenez, A.; Shi, Q.; Canada, E.J.; Kahl, S.D.; Statnick, M.A.; et al. Discovery of aminobenzyloxyarylamides as κ opioid receptor selective antagonists: Application to preclinical development of a κ opioid receptor antagonist receptor occupancy tracer. J. Med. Chem. 2011, 54, 8000–8012. [Google Scholar] [CrossRef] [PubMed]
- Ju, J.; Zhang, L.; Hua, R. Palladium-catalyzed direct intramolecular double a-C–H arylation of 1,5-diketone: A strategy for synthesis of Tröger’s base analogues. Tetrahedron Lett. 2014, 55, 3374–3376. [Google Scholar] [CrossRef]
- Upadhyay, A.; Madhuban, G.; Prasad, D. Synthesis and nematicidal activity of pyrazole derivatives. Pestic. Res. J. 2012, 24, 65–70. [Google Scholar]
- Zhao, F.; Zhao, Q.-J.; Zhao, J.-X.; Zhang, D.-Z.; Wu, Q.-Y.; Jin, Y.-S. Synthesis and cdc25B inhibitory activity evaluation of chalcones. Chem. Nat. Comp. 2013, 49, 206–214. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Gao, S.; Ma, M.; Ren, G.; Liu, H.; Chen, X. Synthesis and antifungal activity of chalcone derivatives. Nat. Prod. Res. 2015, 12, 1–7. [Google Scholar] [CrossRef]
- Montes-Avila, J.; Díaz-Camacho, S.P.; Sicairos-Félix, J.; Delgado-Vargas, F.; Rivero, I.A. Solution-phase parallel synthesis of substituted chalcones and their antiparasitary activity against Giardia lamblia. Bioorg. Med. Chem. 2009, 17, 6780–6785. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, J.; Pandeya, S.N.; Selvakumar, D. Synthesis and biological evaluation of 4,6-diaryl substituted-4,5-dihydro-2-amino pyrimidines. Indian J. Pharm. Sci. 2007, 69, 586–589. [Google Scholar] [CrossRef]
- Park, P.-H.; Kim, H.S.; Jin, X.Y.; Jin, F.; Hur, J.; Ko, G.; Sohn, D.H. KB-34, a newly synthesized chalcone derivative, inhibits lipopolysaccharide-stimulated nitric oxide production in RAW 264.7 macrophages via heme oxygenase-1 induction and blockade of activator protein-1. Eur. J. Pharmacol. 2009, 606, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Muzitano, M.F.; Cruz, E.A.; de Almeida, A.P.; Da Silva, S.A.; Kaiser, C.R.; Guette, C.; Rossi-Bergmann, B.; Costa, S.S. Quercitrin: An antileishmanial flavonoid glycoside from Kalanchoe pinnata. Planta Med. 2006, 72, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, J.L.; Varughese, J.F.; Zhou, Z.; Bartolotti, L.J.; Li, Y. Computational screening and design of S100B ligand to block S100B–p53 interaction. J. Mol. Graph. Model. 2009, 27, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds 1–41 are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ventura, T.L.B.; Calixto, S.D.; Abrahim-Vieira, B.D.A.; Souza, A.M.T.d.; Mello, M.V.P.; Rodrigues, C.R.; Miranda, L.S.d.M.e.; De Souza, R.O.M.A.; Leal, I.C.R.; Lasunskaia, E.B.; et al. Antimycobacterial and Anti-Inflammatory Activities of Substituted Chalcones Focusing on an Anti-Tuberculosis Dual Treatment Approach. Molecules 2015, 20, 8072-8093. https://doi.org/10.3390/molecules20058072
Ventura TLB, Calixto SD, Abrahim-Vieira BDA, Souza AMTd, Mello MVP, Rodrigues CR, Miranda LSdMe, De Souza ROMA, Leal ICR, Lasunskaia EB, et al. Antimycobacterial and Anti-Inflammatory Activities of Substituted Chalcones Focusing on an Anti-Tuberculosis Dual Treatment Approach. Molecules. 2015; 20(5):8072-8093. https://doi.org/10.3390/molecules20058072
Chicago/Turabian StyleVentura, Thatiana Lopes Biá, Sanderson Dias Calixto, Bárbara De Azevedo Abrahim-Vieira, Alessandra Mendonça Teles de Souza, Marcos Vinícius Palmeira Mello, Carlos Rangel Rodrigues, Leandro Soter de Mariz e Miranda, Rodrigo Octavio Mendonça Alves De Souza, Ivana Correa Ramos Leal, Elena B. Lasunskaia, and et al. 2015. "Antimycobacterial and Anti-Inflammatory Activities of Substituted Chalcones Focusing on an Anti-Tuberculosis Dual Treatment Approach" Molecules 20, no. 5: 8072-8093. https://doi.org/10.3390/molecules20058072
APA StyleVentura, T. L. B., Calixto, S. D., Abrahim-Vieira, B. D. A., Souza, A. M. T. d., Mello, M. V. P., Rodrigues, C. R., Miranda, L. S. d. M. e., De Souza, R. O. M. A., Leal, I. C. R., Lasunskaia, E. B., & Muzitano, M. F. (2015). Antimycobacterial and Anti-Inflammatory Activities of Substituted Chalcones Focusing on an Anti-Tuberculosis Dual Treatment Approach. Molecules, 20(5), 8072-8093. https://doi.org/10.3390/molecules20058072