
Structure and Antibacterial Activity of Ambobactin, a New Telomycin-Like Cyclic Depsipeptide Antibiotic Produced by Streptomyces ambofaciens F3

Abstract
:1. Introduction
2. Results and Discussion
2.1. Structural Elucidation

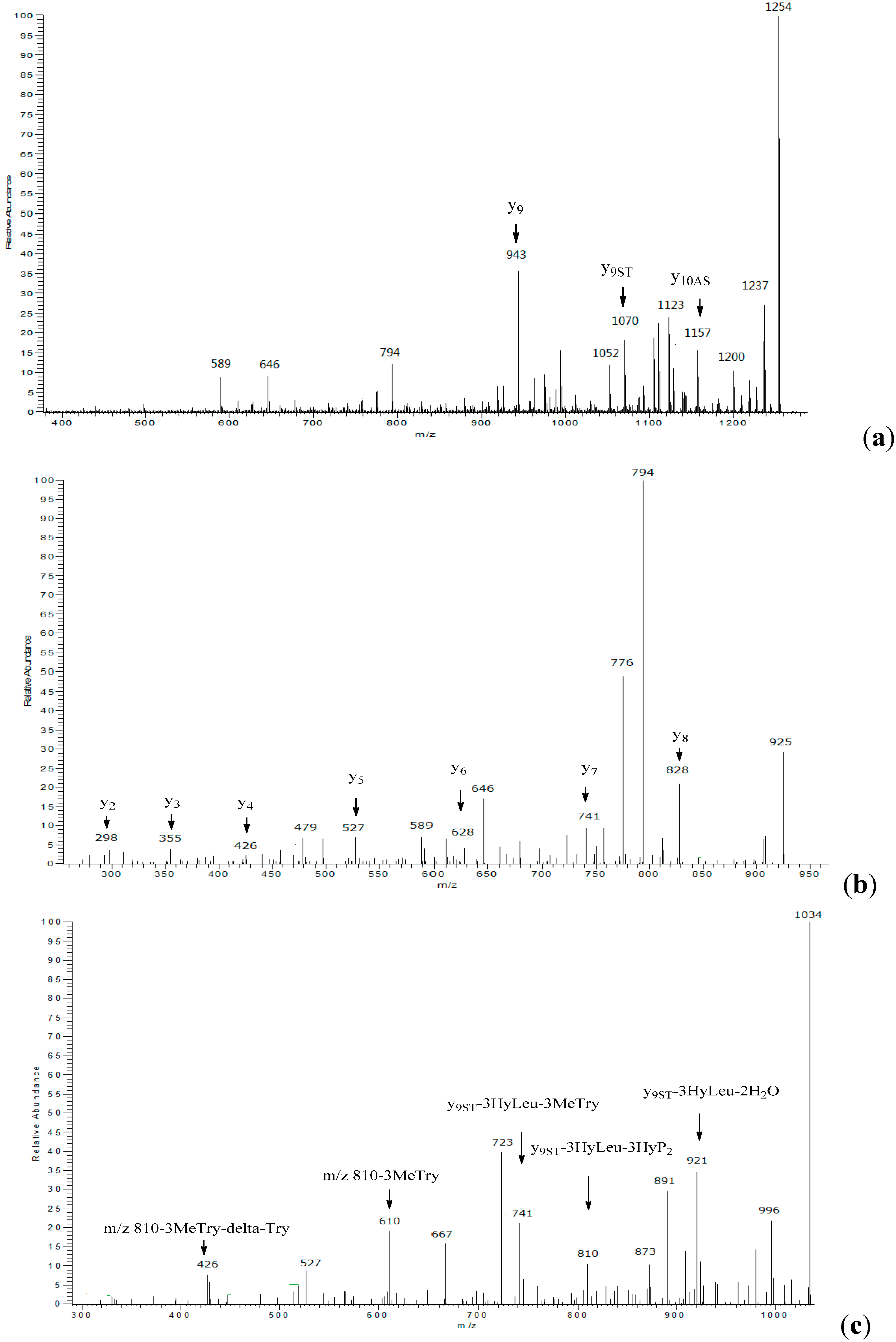{kind=link}
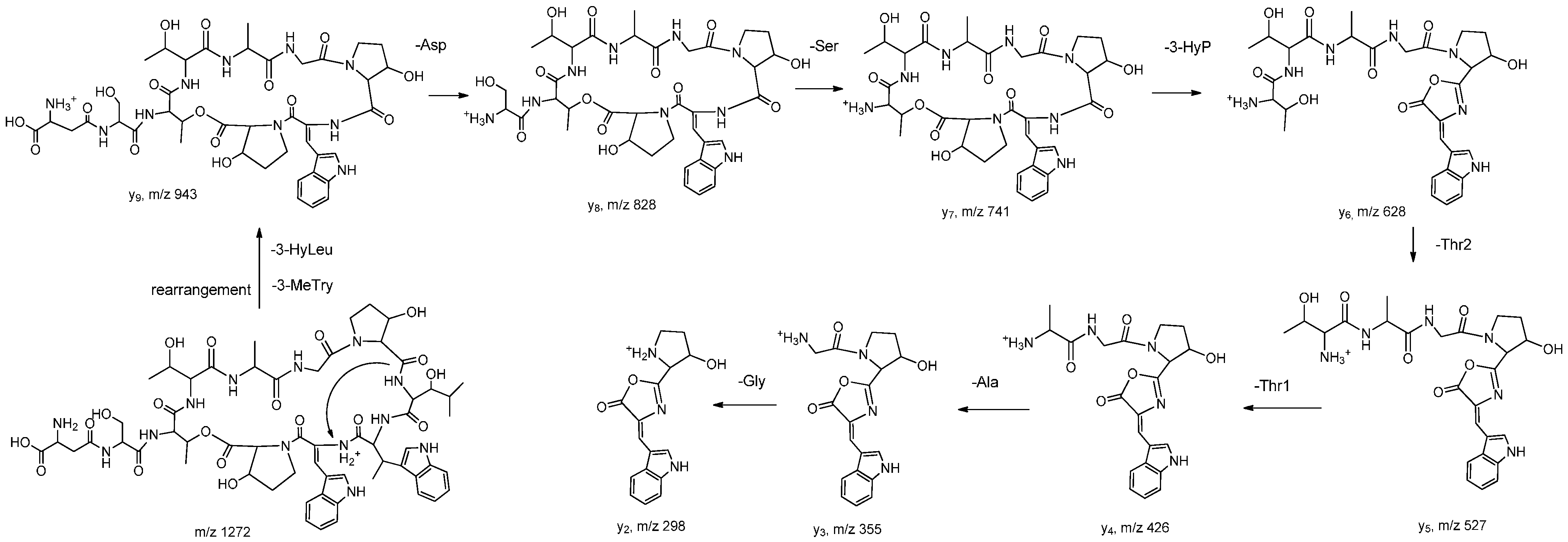{kind=link}
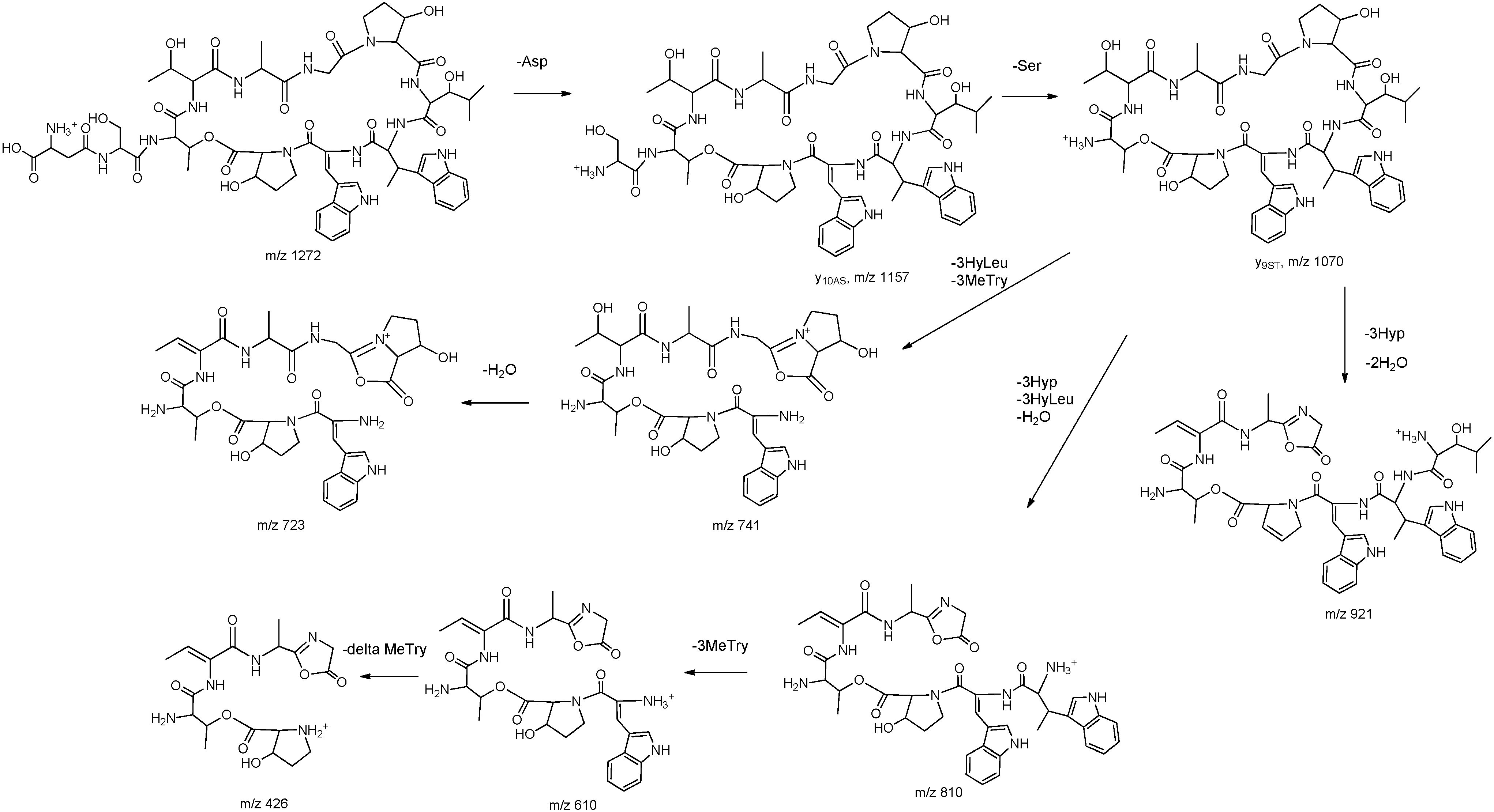{kind=link}
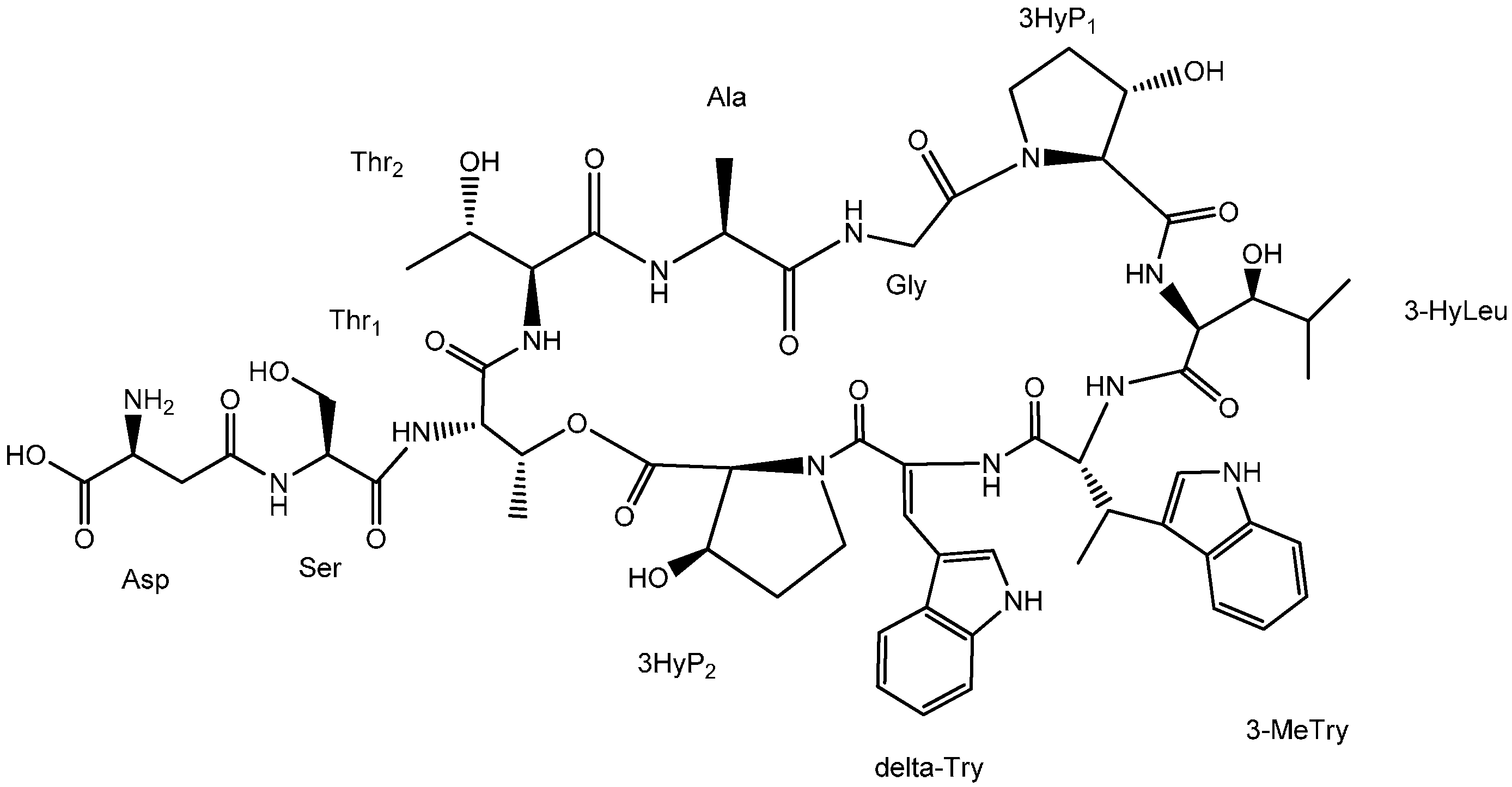{kind=link}
| Ambobactin (1) | Telomycin | |||||
|---|---|---|---|---|---|---|
| δC (ppm) * | δH (ppm, J = Hz) * | HMBC | 1H, 1H-COSY | δC (ppm) * | δH (ppm) * | |
| Aspartic Acid (Asp) | ||||||
| COOH | 170.6 | 170.07 | ||||
| Cα | 51.5 | 3.93 (m) | 170.6 | 2.33, 2.74 | 48.65 | 4.17 |
| Cβ | 38.3 | 2.33 (d, J = 10.5 Hz) | 168.8 | 3.93 | 34.24 | 2.82 |
| 2.74 (t, J = 10.5 Hz) | 168.8 | |||||
| COγ | 168.8 | 168.55 | ||||
| Serine (Ser) | ||||||
| CO | 169.0 | 170.70 | ||||
| Cα | 57.8 | 4.11 (m) | 169.0 | 3.64, 3.74, 8.67 | 54.00 | 4.61 |
| Cβ | 61.5 | 3.64, 3.74 (m) | 4.11, 4.46 | 61.66 | 3.46, 3.68 | |
| NH | 8.67 (s) | 4.11 | 8.43 | |||
| OH | 4.46 (s) | 3.64, 3.74 | ||||
| Threonine1 (Thr1) | ||||||
| CO | 168.9 | 168.28 | ||||
| Cα | 57.9 | 4.46 (m) | 168.9 | 4.99, 7.76 | 58.3 | 4.25 |
| Cβ | 71.2 | 4.99 (m) | - | 4.46, 1.15 | 71.0 | 4.98 |
| Cγ | 15.9 | 1.15 (d, J = 6.0 Hz) | 57.9, 71.2 | 4.99 | 15.77 | 1.23 |
| NH | 7.76 (d, J = 7.0 Hz) | 4.46 | 8.10 | |||
| 3-Hydroxy-l-Proline (3-HyP1) | ||||||
| CO | 171.3 | |||||
| C2 | 67.5 | 4.19 (m) | 73.6, 171.3 | 4.21 | 66.8 | 4.21 |
| C3 | 73.6 | 4.21 (m) | 1.96, 2.16, 4.19, 5.78 | 73.07 | 4.22 | |
| C4 | 33.9 | 1.96, 2.16 (m) | 3.52, 3.82, 4.21 | 33.32 | 2.00, 2.19 | |
| C5 | 44.9 | 3.52, 3.82 (m) | 1.96, 2.16 | 44.32 | 3.49, 3.83 | |
| OH | 5.78 (s) | 4.21 | ||||
| α,β-Dehydrotryptophan (ΔTry) | ||||||
| CO | 163.9 | 163.57 | ||||
| C2 | 122.9 | - | 122.17 | |||
| C3 | 122.9 | 7.46 (s) | 127.7, 163.9 | 7.62, 7.08 | 122.67 | |
| C4 | 109.0 | - | 108.38 | |||
| C5 | 128.2 | 7.95 (s) | 109.0, 127.7, 135.9 | 11.72 | 127.67 | 7.95 |
| C6a | 135.9 | - | 135.34 | |||
| C7 | 112.3 | 7.38 (d, J = 8.2 Hz) | 120.6, 127.7 | 7.14 | 111.73 | 7.38 |
| C8 | 122.5 | 7.14 (t, J = 7.6 Hz) | 118.1, 127.7, 135.9 | 7.38 | 121.97 | 7.13 |
| C9 | 120.6 | 7.08 (t, J = 7.6 Hz) | 112.3 | 7.62 | 120.08 | 7.08 |
| C10 | 118.1 | 7.62 (d, J = 8.0 Hz) | 109.0, 122.5, 127.7, 135.9 | 7.08 | 117.54 | 7.60 |
| C10a | 127.7 | - | 127.67 | |||
| 6-NH | 11.72 (s) | 109.0, 127.7, 135.9 | 11.70 | |||
| NH | 10.08 (s) | 163.9, 122.5, 128.2 | 10.14 | |||
| 3-Methyl-Tryptophan (3-MeTry) | ||||||
| CO | 171.8 | |||||
| C2 | 60.2 | 4.46 (m) | 33.2, 171.8 | 7.56, 8.69 | 60.0 | 4.47 |
| C3 | 33.2 | 3.65 (m) | 112.0 | 1.24, 4.46 | 32.49 | 3.61 |
| C4 | 117.5 | - | - | 116.6 | ||
| C5 | 123.3 | 7.08 (s) | 112.0, 117.5, 126.3, 137.2 | 10.75 | 122.87 | |
| C6a | 137.2 | - | - | 136.56 | ||
| C7 | 112.0 | 7.28 (d, J = 8.2 Hz) | 126.3, 118.7 | 10.75, 7.00 | 111.43 | 7.27 |
| C8 | 121.2 | 7.00 (t, J = 7.6 Hz) | 119.5, 121.2, 126.3, 137.2 | 7.28 | 120.63 | 7.01 |
| C9 | 118.7 | 6.93 (t, J = 7.6 Hz) | 112.0, 126.3 | 7.56 | 118.17 | 6.92 |
| C10 | 119.5 | 7.56 (d, J = 8.2 Hz) | 117.5, 121.2, 126.3, 137.2 | 6.93 | 119.01 | 7.58 |
| C10a | 126.3 | - | - | 125.56 | ||
| C11 | 19.8 | 1.24 (d, J = 7.0 Hz) | 33.2, 60.2, 117.5 | 3.65 | 18.37 | |
| 6-NH | 10.75 (s) | 117.5, 126.3, 137.2 | 7.08, 7.28 | 10.75 | ||
| NH | 8.69 (s) | 7.74 | ||||
| 3-Hydroxy-Leucine (3-HyLeu) | ||||||
| CO | 171.6 | |||||
| Cα | 52.4 | 4.49 (m) | 171.6 | 3.32, 6.62 | 51.60 | 4.54 |
| Cβ | 75.2 | 3.32 (m) | - | 1.94, 4.49, 4.74 | 75.18 | 3.29 |
| Cγ | 29.2 | 1.94 (m) | - | 0.79, 0.84, 3.32 | 28.78 | 1.94 |
| Cδ1 | 20.6 | 0.84 (d, J = 6.6 Hz) | 16.5, 29.2, 75.2 | 1.94 | 19.63 | 0.87 |
| Cδ2 | 16.5 | 0.79 (d, J = 6.6 Hz) | 20.6, 29.2, 75.2 | 1.94 | 17.19 | 0.84 |
| OH | 4.74 (s) | 3.32 | ||||
| NH | 6.62 (s) | 4.49 | ||||
| 3-Hydroxy-l-Proline (3-HyP2) | ||||||
| CO | 169.4 | |||||
| C2 | 62.2 | 4.67 (m) | 169.4 | 4.71 | 63.02 | 4.61 |
| C3 | 69.5 | 4.71 (m) | 1.76, 2.03, 4.67, 4.49 | 69.35 | 4.61 | |
| C4 | 32.1 | 1.76, 2.03 (m) | 3.74, 3.93, 4.71 | 33.43 | 1.90 | |
| C5 | 45.0 | 3.74, 3.93 (m) | 1.76, 2.03 | 45.25 | 3.61 | |
| OH | 4.49 (s) | 4.71 | ||||
| Glycine (Gly) | ||||||
| CO | 169.1 | |||||
| Cα | 41.5 | 3.80, 4.44 (m) | 169.1 | 8.86 | 41.0 | 3.84, 4.42 |
| NH | 8.86 (s) | 3.80, 4.44 | 8.66 | |||
| Alanine (Ala) | ||||||
| CO | 172.4 | 171.4 | ||||
| Cα | 48.5 | 4.45 (m) | 172.4 | 1.05, 7.73 | 47.85 | 4.45 |
| Cβ | 17.5 | 1.05 (d, J = 6.0 Hz) | 48.5, 172.4 | 4.45 | 17.05 | 1.02 |
| NH | 7.73 (d, J = 7.0 Hz) | 4.45 | 8.05 | |||
| Threonine2 (Thr2) | ||||||
| CO | 169.3 | |||||
| Cα | 58.8 | 4.09 (m) | 67.2, 169.3 | 3.82, 7.42 | 57.22 | 4.22 |
| Cβ | 67.2 | 3.82 (m) | 1.07, 4.09 | 66.91 | 3.64 | |
| Cγ | 21.5 | 1.07 (d, J = 6.0 Hz) | 67.2, 58.8 | 3.82 | 20.65 | 1.00 |
| NH | 7.42 (s) | 4.09 | 7.53 | |||




| Derivatives | Time (min) | ||||||
|---|---|---|---|---|---|---|---|
| Asp | Ser | 3-HyLeu | Thr | Ala | 3-HyP | 3-MeTry | |
| L-FDAA | 10.62 | 7.12 | 21.36 | 7.93 | 18.86 | 10.47 | 44.11 |
| D-FDAA | 12.21 | 7.55 | 34.95 | 8.68 | 26.99 | 12.13 | 41.59 |
2.2. Antibacterial Activity
| Compounds | MIC (μg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| BS | BC | SA | EC | PA | ErC | PS | AS | XO | |
| Ambobactin | 3.13 | 25 | 25 | 6.25 | >100 | 6.25 | 6.25 | 6.25 | 6.25 |
| Ampicillin | 1.56 | 6.25 | 6.25 | 3.13 | >100 | 6.25 | 6.25 | 6.25 | 6.25 |
3. Experimental Section
3.1. General Experimental Procedures
3.2. Microorganism and Fermentation
3.3. Extraction and Isolation
3.4. Acid Hydrolysis and Advanced Marfey’s Analysis
3.5. HPLC-UV/MS Analysis
3.6. Tandem MS Analysis
3.7. Antibacterial Activity
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Joo, S.H. Cyclic peptides as therapeutic agents and biochemical tools. Biomol. Ther. 2012, 20, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Bockus, A.T.; McEwen, C.M.; Lokey, R.S. Form and function in cyclic peptide natural products: A pharmacokinetic perspective. Curr. Top. Med. Chem. 2013, 13, 821–836. [Google Scholar] [CrossRef] [PubMed]
- Kronenwerth, M.; Bozhüyük, K.A.J.; Kahnt, A.S.; Steinhilber, D.; Gaudriault, S.; Kaiser, M.; Bode, H.B. Characterisation of taxlllaids A–G: Natural products from Xenorhabdus indica. Chem. Eur. J. 2014, 20, 17478–17487. [Google Scholar] [CrossRef] [PubMed]
- Munar, M.Y.; Doyle, I.C.; Meyer, M.M. Cyclosporine and vancomycin disposition during continuous venovenous hemodiafiltration. Ann. Pharmacother. 1995, 29, 374–377. [Google Scholar] [PubMed]
- Misiek, M.; Fardig, O.B.; Gourevitch, A.; Johnson, D.L.; Hooper, I.R.; Lein, J. Telomycin, a new antibiotic. Antibiot. Annu. 1957–1958, 5, 852–855. [Google Scholar]
- Sheehan, J.C.; Mania, D.; Nakamura, S.; Stock, J.A.; Maeda, K. The structure of telomycin. J. Am. Chem. Soc. 1968, 90, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Keller, L.; Bauer, A.; Brönstrup, M.; Froidbise, A.; Hammann, P.; Herrmann, J.; Mondesert, G.; Kurz, M.; Schiell, M.; et al. Biosynthetic studies of telomycin reveal lipopeptides with enhanced activity. J. Am. Chem. Soc. 2015, 137, 7692–7705. [Google Scholar] [CrossRef] [PubMed]
- Vela Gurovic, M.C.; Müller, S.; Domin, N.; Seccareccia, I.; Nietzsche, S.; Martin, K.; Nett, M. Micromonospora schwarzwaldensis sp. nov., a producer of telomycin, isolated from soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 3812–3817. [Google Scholar]
- Whaley, H.A.; Patterson, E.L.; Dann, M.; Shu, P.; Swift, M.E.; Porter, J.N.; Redin, G. LL-AO341A and B, new antibiotics. I. Isolation and characterization. Antimicrob. Agents Chemother. 1966, 6, 587–590. [Google Scholar] [PubMed]
- Whaley, H.A.; Patterson, E.L.; Kunstmann, M.P.; Bohonos, N. LL-AO341A and B, new antibiotics. II. Chemical properties. Antimicrob. Agents Chemother. 1966, 6, 591–594. [Google Scholar] [PubMed]
- Kumar, N.G.; Urry, D.W. Proton magnetic resonance assignments of the polypeptide antibiotic telomycin. Biochemistry 1973, 12, 3811–3817. [Google Scholar] [CrossRef] [PubMed]
- Oliva, B.; Maiese, W.M.; Greenstein, M.; Borders, D.B.; Chopra, I. Mode of action of the cyclic depsipeptide antibiotic LL-AO341 β1 and partial characterization of a Staphylococcus aureus mutant resistant to the antibiotic. J. Antimicrob. Chemother. 1993, 32, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Ngoka, L.C.; Gross, M.L. A nomenclature system for labeling cyclic peptide fragments. J. Am. Soc. Mass. Spectrom. 1999, 10, 360–363. [Google Scholar] [CrossRef]
- Paizs, B.; Suhai, S. Fragmentation pathways of protonated peptides. Mass Spectrom. Rev. 2005, 24, 508–548. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Ikai, Y.; Oka, H.; Suzuki, M.; Harada, K. A nonempirical method using LC/MS for determination of the absolute configuration of constituent amino acids in a peptide: Combination of Marfey’s method with mass spectrometry and its practical application. Anal. Chem. 1997, 69, 5146–5151. [Google Scholar] [CrossRef]
- Bhushan, R.; Brückner, H. Marfey’s reagent for chiral amino acid analysis: A review. Amino Acids 2004, 27, 231–247. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.Q.; Wei, S.P.; Fan, L.X.; Wu, W.J. Three novel cyclic hexapeptides from Streptomyces alboflavus 313 and their antibacterial activity. Eur. J. Med. Chem. 2012, 50, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Uemoto, H.; Yahiro, Y.; Shigemori, H.; Tsuda, M.; Takao, T.; Shimonishi, Y.; Kobayashi, J. Keramamides K and L, new cyclic peptides containing unusual tryptophan residue from Theonella sponge. Tetrahedron 1998, 54, 6719–6724. [Google Scholar] [CrossRef]
- Sample Availability: Samples of Streptomyces ambofaciens F3 and ambobactin are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, S.; Zhang, W.; Ji, Z. Structure and Antibacterial Activity of Ambobactin, a New Telomycin-Like Cyclic Depsipeptide Antibiotic Produced by Streptomyces ambofaciens F3. Molecules 2015, 20, 16278-16289. https://doi.org/10.3390/molecules200916278
Wei S, Zhang W, Ji Z. Structure and Antibacterial Activity of Ambobactin, a New Telomycin-Like Cyclic Depsipeptide Antibiotic Produced by Streptomyces ambofaciens F3. Molecules. 2015; 20(9):16278-16289. https://doi.org/10.3390/molecules200916278
Chicago/Turabian StyleWei, Shaopeng, Wenhao Zhang, and Zhiqin Ji. 2015. "Structure and Antibacterial Activity of Ambobactin, a New Telomycin-Like Cyclic Depsipeptide Antibiotic Produced by Streptomyces ambofaciens F3" Molecules 20, no. 9: 16278-16289. https://doi.org/10.3390/molecules200916278
APA StyleWei, S., Zhang, W., & Ji, Z. (2015). Structure and Antibacterial Activity of Ambobactin, a New Telomycin-Like Cyclic Depsipeptide Antibiotic Produced by Streptomyces ambofaciens F3. Molecules, 20(9), 16278-16289. https://doi.org/10.3390/molecules200916278