
Effect of Amaranthus on Advanced Glycation End-Products Induced Cytotoxicity and Proinflammatory Cytokine Gene Expression in SH-SY5Y Cells

Abstract
:1. Introduction

2. Results and Discussion
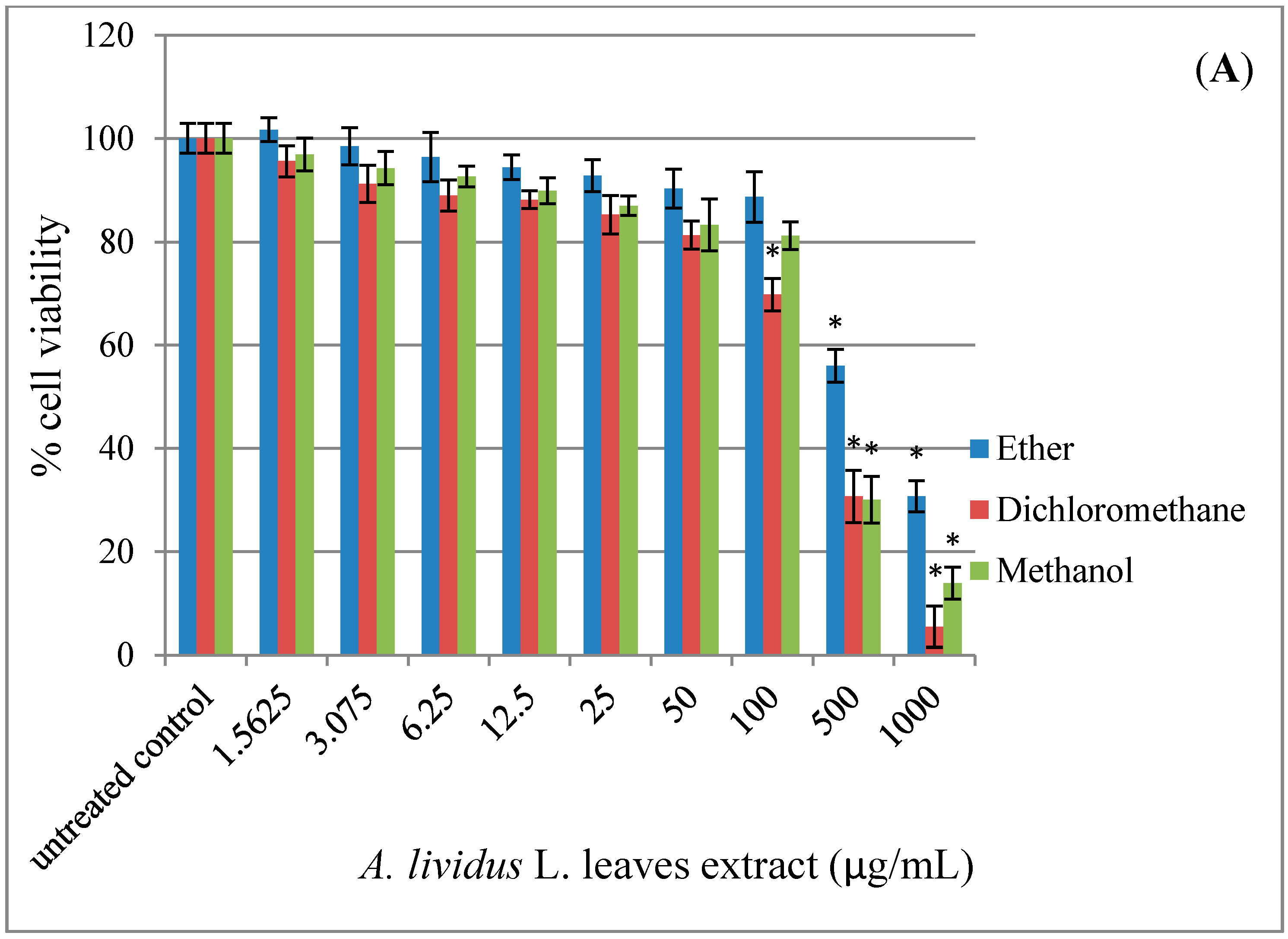
2.1. Effect of A. lividus L. and A. tricolor L. Extracts on Cell Viability in Human Neuroblastoma SH-SY5Y Cells


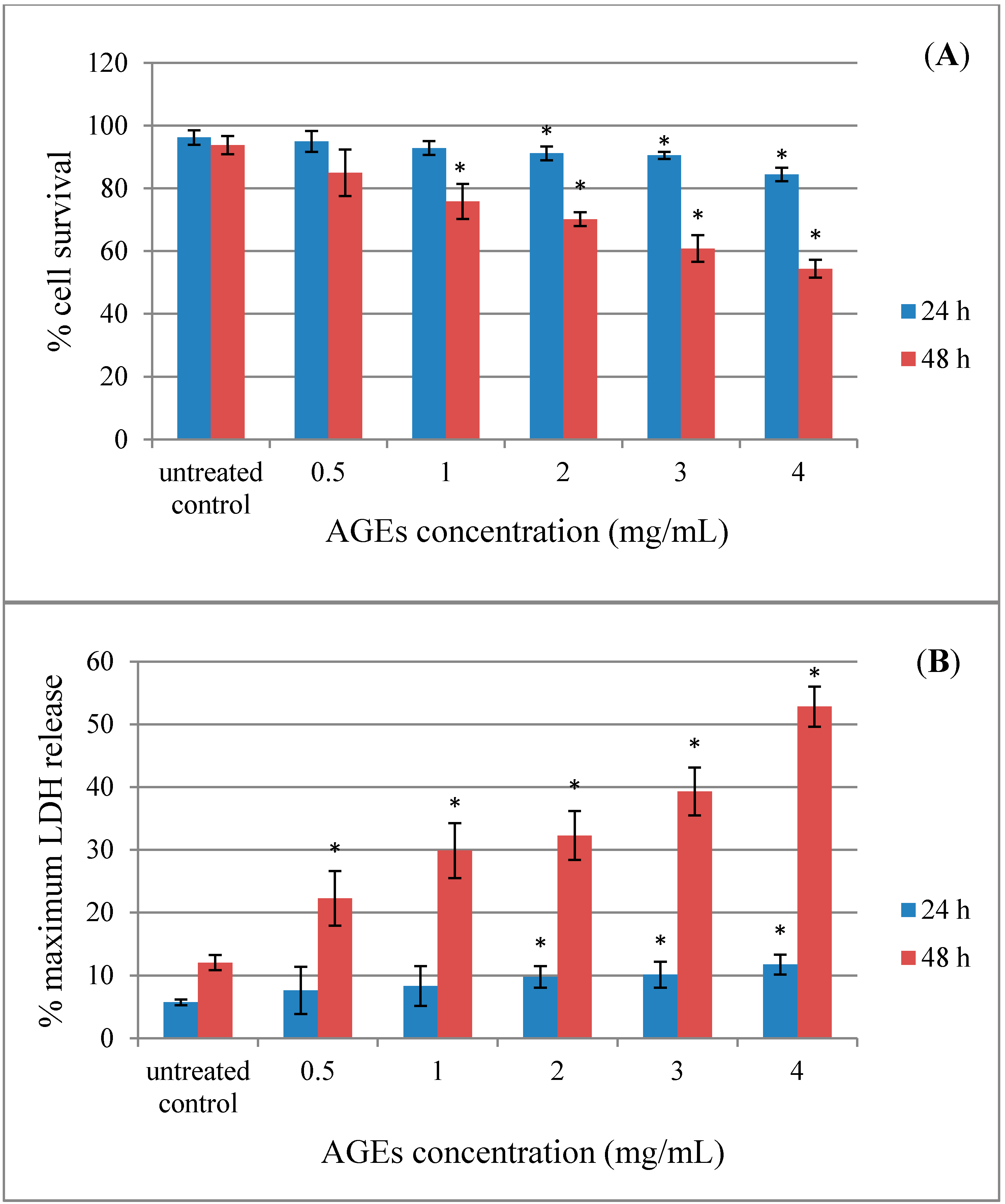
2.2. Effect of AGEs on Cytotoxicity in Human Neuroblastoma SH-SY5Y Cells

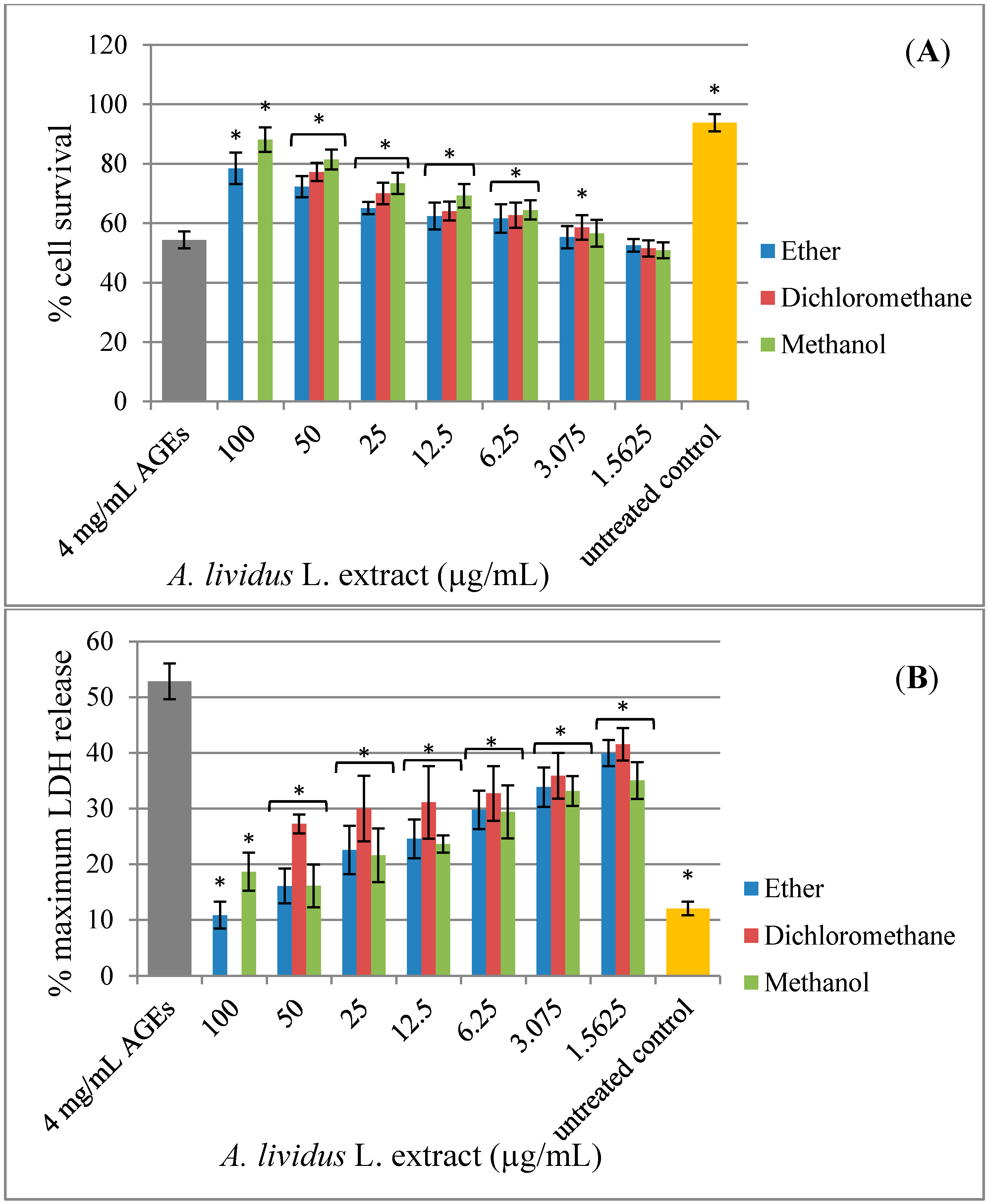
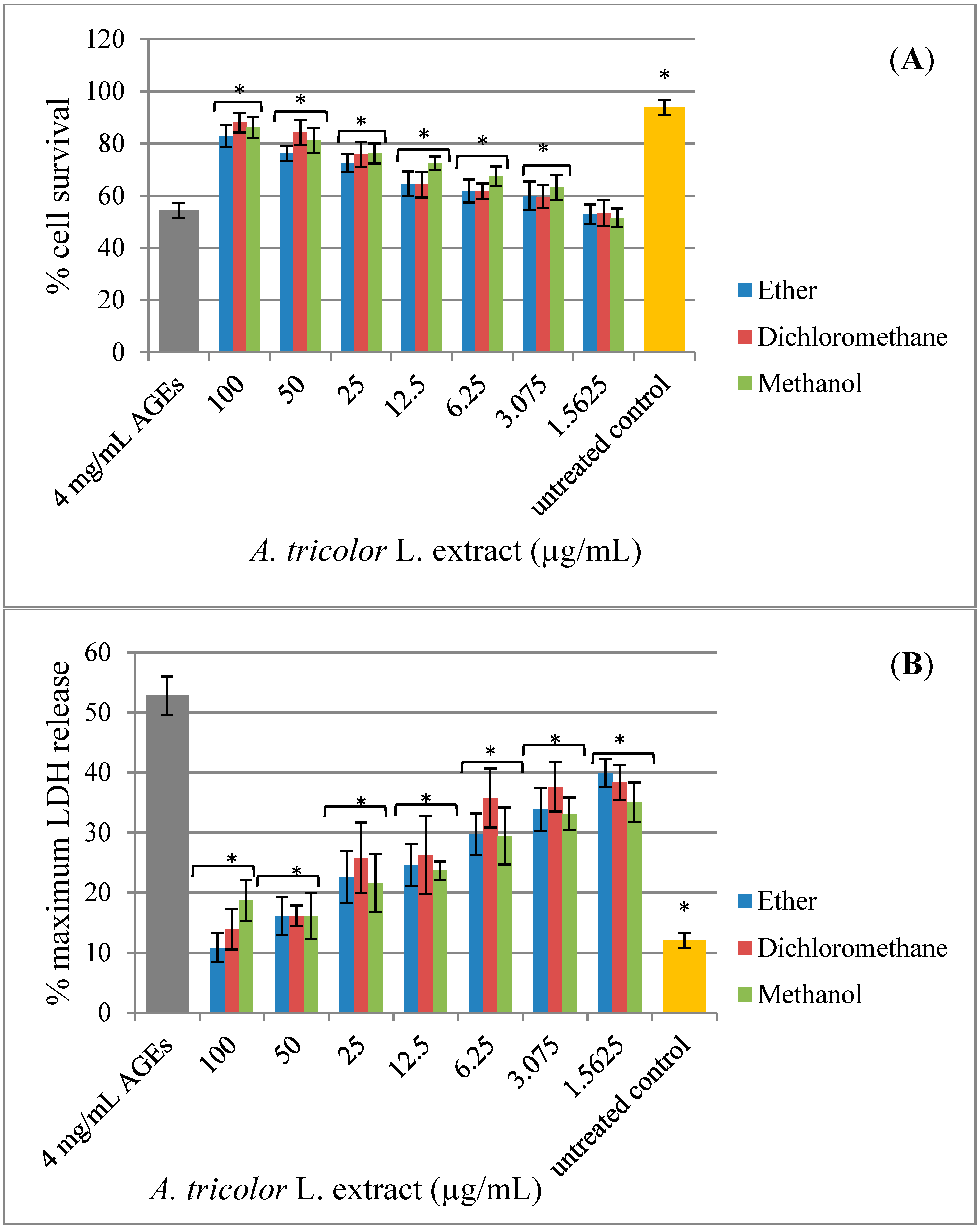
2.3. Effect of A. lividus L. and A. tricolor L. Extracts on AGEs-Induced Cytotoxicity


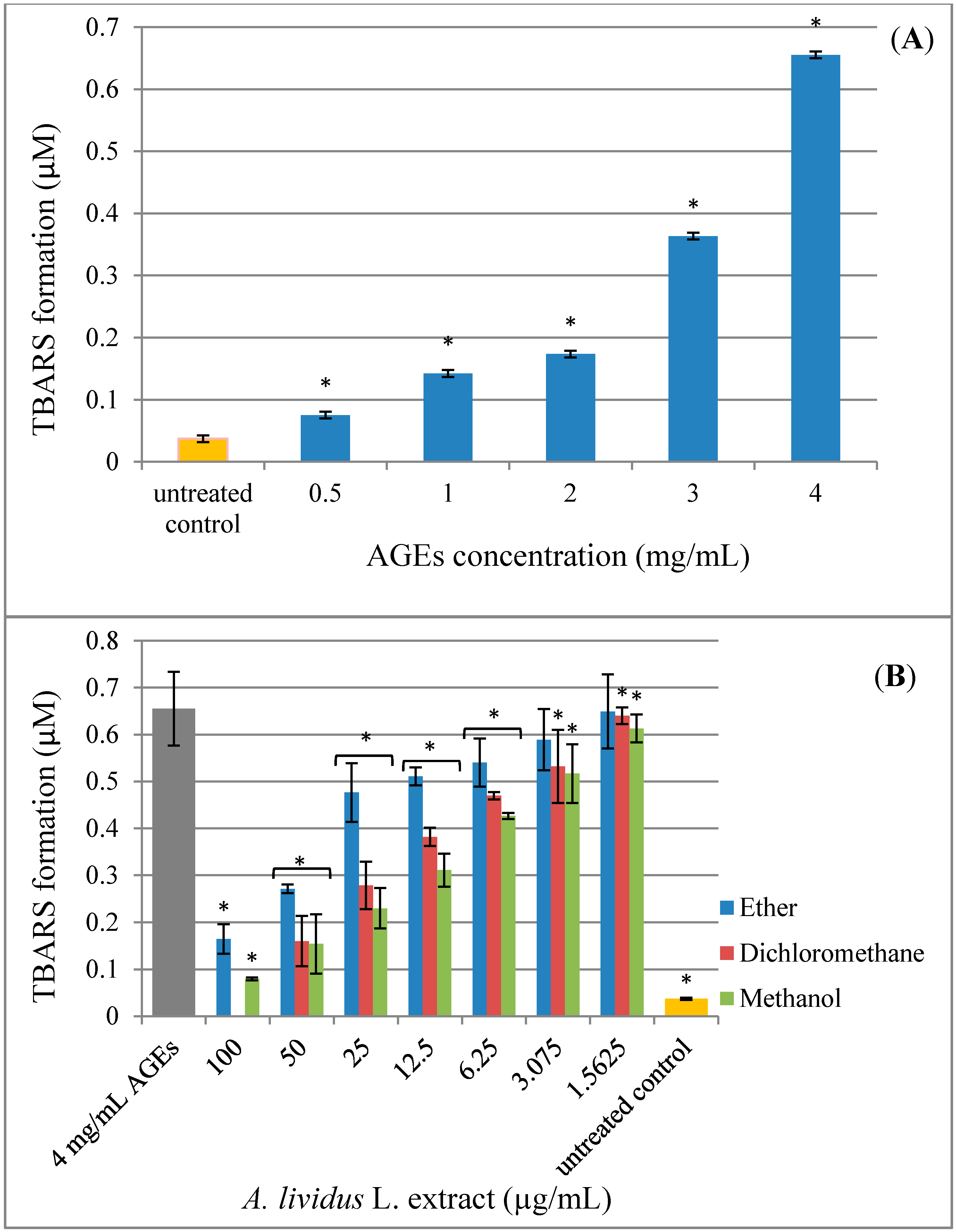
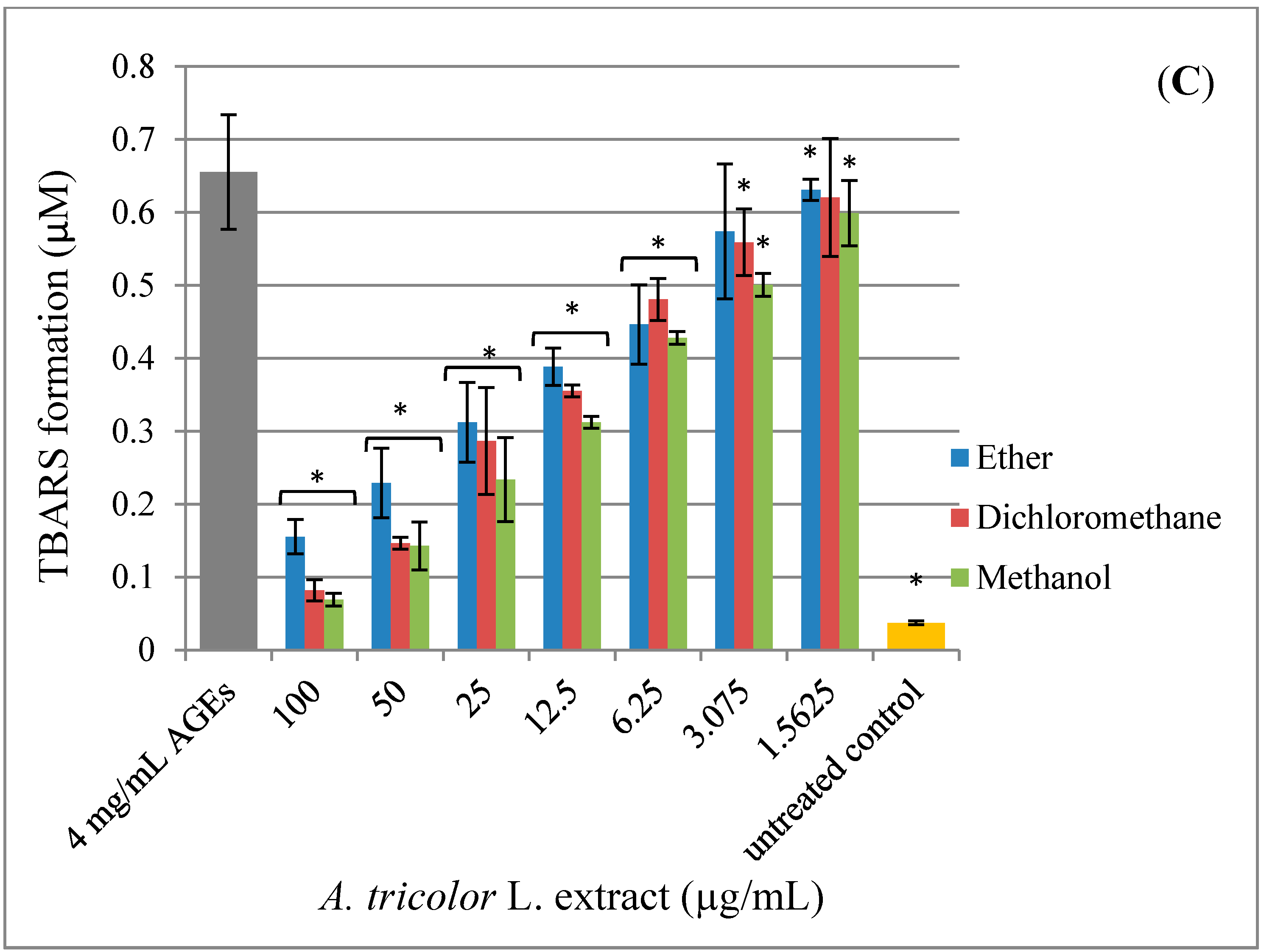
2.4. Effect of A. lividus L. and A. tricolor L. Extracts on AGEs-Induced Generation of Oxidant Stress


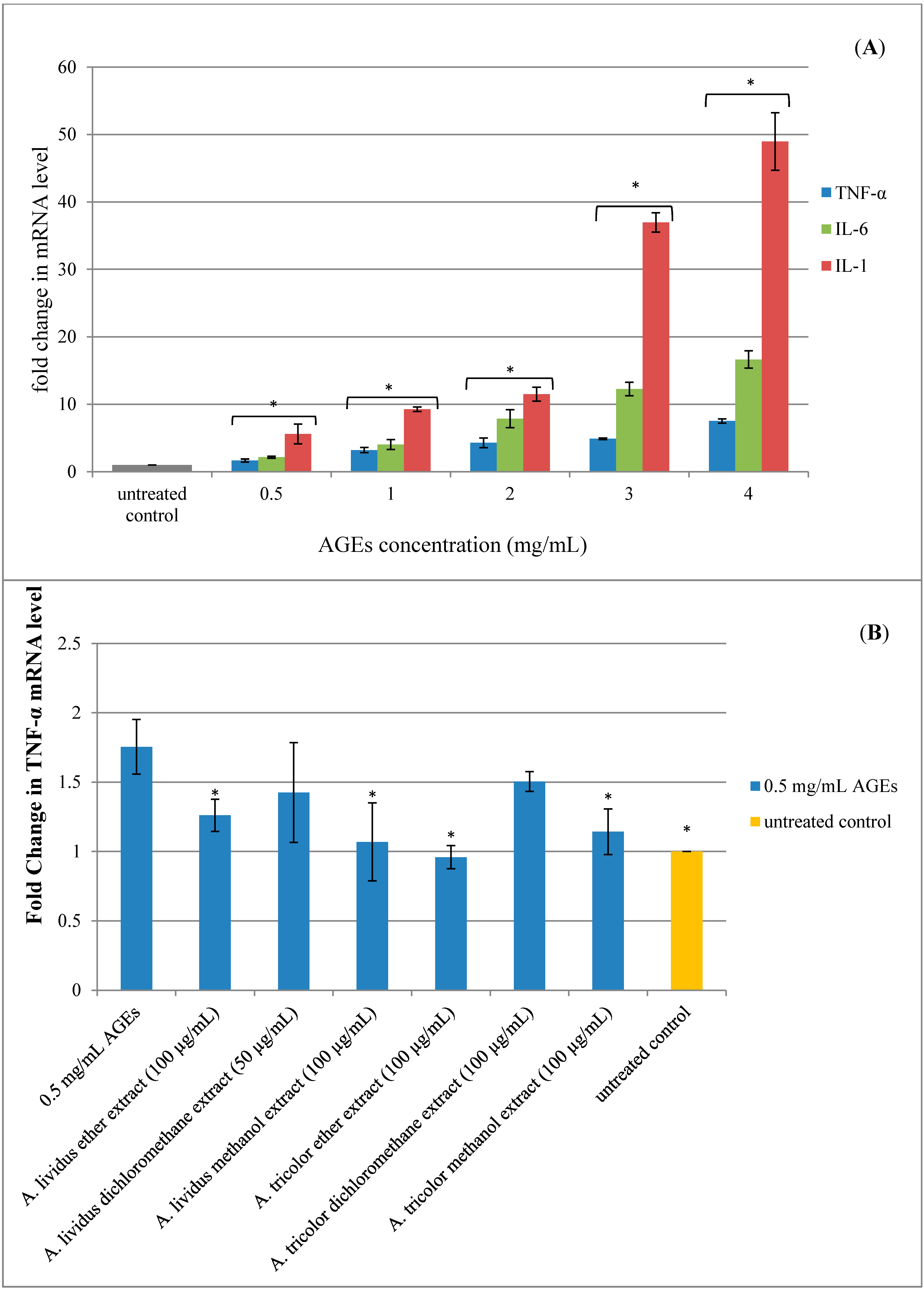
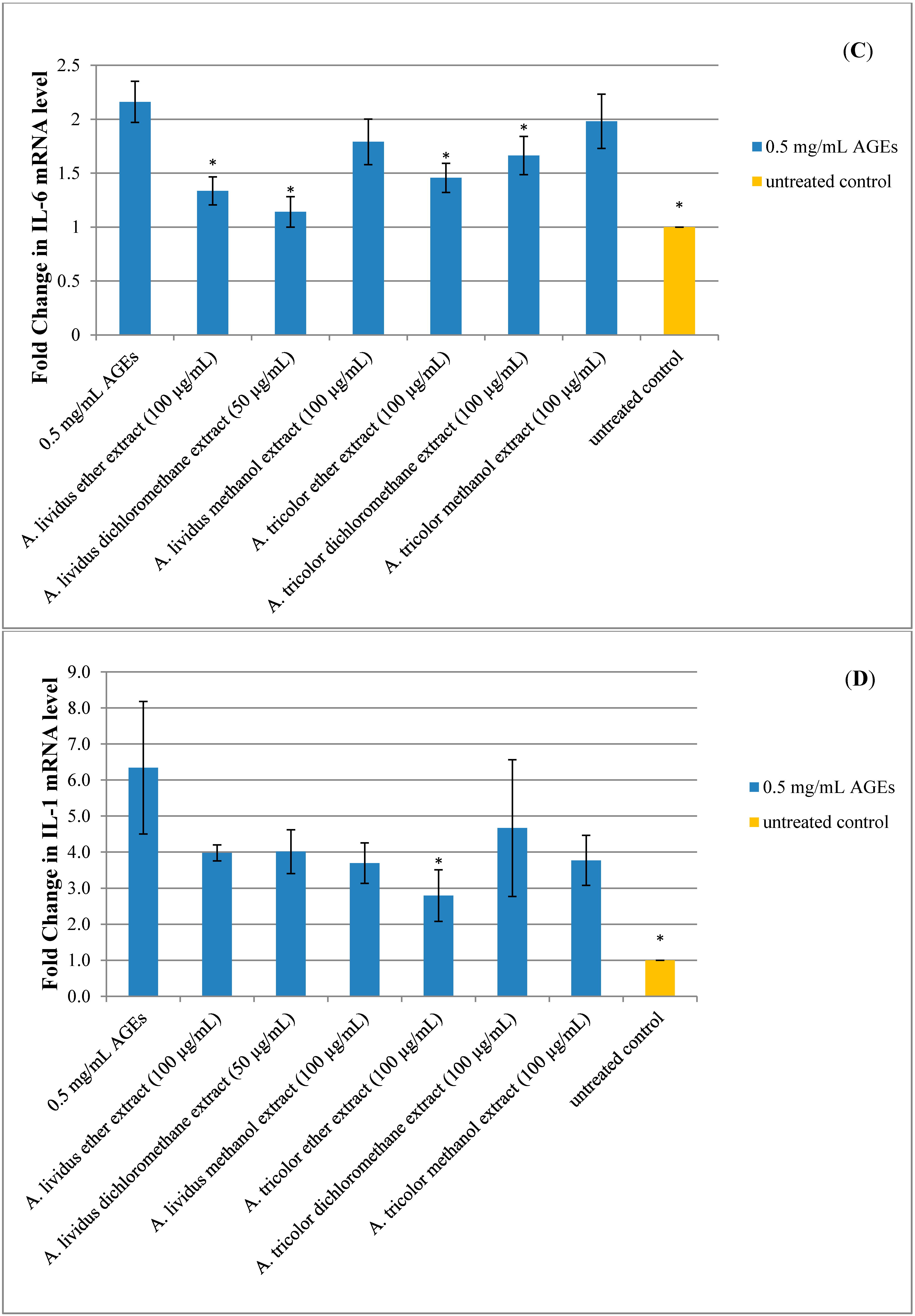
2.5. Effect of A. lividus L. and A. tricolor L. Extracts on AGEs-Induced Expression of Proinflammatory Cytokine Genes in Human Neuroblastoma SH-SY5Y Cells


3. Experimental Section
3.1. Plant Material
3.2. Cell Culture
3.3. Advanced Glycation End-Product Preparation
3.4. 3-(4, 5-Ethylthiazol-2-yl)-2, 5-diphenyltetrazolium Bromide (MTT) Assay
3.5. Trypan Blue Exclusion Assay
3.6. Lactate Dehydrogenase Release Assay
3.7. Malondialdehyde Assay
3.8. Quantitative-Reverse-Transcription Polymerase Chain Reaction Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Annealing Temperature (°C) | Product Size (bp) | |
|---|---|---|---|---|
| TNF-α | Forward primer | 5ʹ TCTCGAACCCCGAGTGACAA 3ʹ | 55 | 181 |
| Reverse primer | 5ʹ TGAAGAGGACCTGGGAGTAG 3ʹ | |||
| IL-1 | Forward primer | 5ʹ ACCAAACCTCTTCGAGGCAC 3ʹ | 56 | 300 |
| Reverse primer | 5ʹ CATGGCCACAACAACTGACG 3ʹ | |||
| IL-6 | Forward primer | 5ʹ GAAGAGAGCCCTCAGGCTGGACTG 3ʹ | 64 | 627 |
| Reverse primer | 5ʹ TGAACTCCTTCTCCACAAGCGC 3ʹ | |||
| GAPDH | Forward primer | 5ʹ GAAAGCCTGCCGGTGACTAA 3ʹ | 60 | 370 |
| Reverse primer | 5ʹ TCGCCCCACTTGATTTTGGA 3ʹ | |||
| Step | Condition | Cycle (s) |
|---|---|---|
| Pre-denaturation | 95 °C, 10 min | 1 |
| Denaturation | 95 °C, 15 s | 35–40 |
| Annealing/extension | 60 °C, 15 s (GAPDH) | |
| 55 °C, 15 s (TNF-α) | ||
| 56 °C, 15 s (IL-1) | ||
| 64 °C, 30 s (IL-6) | ||
| Detection | Scan | |
| Melting | 1 |
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yong, V.W. Inflammation in neurological disorders: A help or a hindrance? Neuroscientist 2010, 16, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Emerit, J.; Edeas, M.; Bricaire, F. Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 2004, 58, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.L.; Yang, C.M. Role of redox signaling in neuroinflammation and neurodegenerative diseases. BioMed Res. Int. 2013, 2013, 484613. [Google Scholar] [CrossRef] [PubMed]
- Haddad, J.J. Oxygen-sensitive pro-inflammatory cytokines, apoptosis signaling and redox-responsive transcription factors in development and pathophysiology. Cytokines Cell. Mol. Ther. 2002, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, N. Advanced glycation endproducts—role in pathology of diabetic complications. Diabetes Res. Clin. Pract. 2005, 67, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Woltjer, R.L.; Maezawa, I.; Ou, J.J.; Montine, K.S.; Montine, T.J. Advanced glycation endproduct precursor alters intracellular amyloid-β/A β PP carboxy-terminal fragment aggregation and cytotoxicity. J. Alzheimer’s Dis. 2003, 5, 467–476. [Google Scholar]
- Mruthinti, S.; Sood, A.; Humphrey, C.L.; Swamy-Mruthinti, S.; Buccafusco, J.J. The induction of surface beta-amyloid binding proteins and enhanced cytotoxicity in cultured PC-12 and IMR-32 cells by advanced glycation end products. Neuroscience 2006, 142, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, P.; Cerami, A. Protein glycation, diabetes, and aging. Recent Prog. Horm. Res. 2001, 56, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hurley, L.L.; Tizabi, Y. Neuroinflammation, neurodegeneration, and depression. Neurotox. Res. 2013, 23, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Howes, M.J.; Perry, E. The role of phytochemicals in the treatment and prevention of dementia. Drugs Aging 2011, 28, 439–468. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.M.; Li, J.T. A systematic review of single Chinese herbs for Alzheimer’s disease treatment. Evid. Based Complement. Alternat. Med. 2011, 2011, 640284. [Google Scholar] [CrossRef] [PubMed]
- López-Mejía, O.A.; López-Malo, A.; Palou, E. Antioxidant capacity of extracts from amaranth (Amaranthus hypochondriacus L.) seeds or leaves. Ind. Crops Prod. 2014, 53, 55–59. [Google Scholar]
- Kraujalis, P.; Venskutonis, P.R.; Kraujalienė, V.; Pukalskas, A. Antioxidant properties and preliminary evaluation of phytochemical composition of different anatomical parts of amaranth. Plant Foods Hum. Nutr. 2013, 68, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Sian, J.; Dexter, D.T.; Lees, A.J.; Daniel, S.; Agid, Y.; Javoy‐Agid, F.; Jenner, P.; Marsden, C.D. Alterations in glutathione levels in Parkinson’s disease and other neurodegenerative disorders affecting basal ganglia. Ann. Neurol. 1994, 36, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Multhaup, G.; Ruppert, T.; Schlicksupp, A.; Hesse, L.; Beher, D.; Masters, C.L.; Beyreuther, K. Reactive oxygen species and Alzheimer’s disease. Biochem. Pharmacol. 1997, 54, 533–539. [Google Scholar] [CrossRef]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Nicholson, L.F. Expression of the receptor for advanced glycation end products in Huntington’s disease caudate nucleus. Brain Res. 2004, 1018, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.S.; Chen, X.; Fu, J.; Chen, M.; Zhu, H.J.; Roher, A.; Slattery, T.; Zhao, L.; Nahashima, M.; Morser, J.; et al. RAGE and amyloid-β peptide neurotoxicity in Alzheimer’s disease. Nature 1996, 382, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, M.; Bucala, R.; Suzuki, T.; Ohkubo, T.; Yamazaki, M.; Koike, T.; Kameda, Y.; Makita, Z. Neurotoxicity of advanced glycation end-products for cultured cortical neurons. J. Neuropathol. Exp. Neurol. 2000, 59, 1094–1105. [Google Scholar] [PubMed]
- Ozsoy, N.; Yilmaz, T.; Kurt, O.; Can, A.; Yanardag, R. In vitro antioxidant activity of Amaranthus lividus L. Food Chem. 2009, 116, 867–872. [Google Scholar] [CrossRef]
- Anitha, R.; Ponbavani, S. Antioxidant and anticoagulant activity in Amaranthus gangeticus L. aqueous leaf extract. WJPPS 2013, 2682–2688. [Google Scholar]
- Zhang, J.; Slevin, M.; Duraisamy, Y.; Gaffney, J.; Smith, C.A.; Ahmed, N. Comparison of protective effects of aspirin, d-penicillamine and vitamin E against high glucose-mediated toxicity in cultured endothelial cells. Biochim. Biophys. Acta 2006, 1762, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Kajimoto, Y.; Miyagawa, J.I.; Matsuoka, T.A.; Fujitani, Y.; Umayahara, Y.; Hanafusa, T.; Matsuzawa, Y.; Yamasaki, Y.; Hori, M. Beneficial effects of antioxidants in diabetes: Possible protection of pancreatic β-cells against glucose toxicity. Diabetes 1999, 48, 2398–2406. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, K.; Naito, Y.; Hasegawa, G.; Nakamura, N.; Takahashi, J.; Yoshikawa, T. Astaxanthin protects β-cells against glucose toxicity in diabetic db/db mice. Redox Rep. 2002, 7, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.D.; Schmidt, A.M.; Anderson, G.M.; Zhang, J.; Brett, J.; Zou, Y.S.; Pinsky, D.; Stern, D. Enhanced cellular oxidant stress by the interaction of advanced glycation end products with their receptors/binding proteins. J. Biol. Chem. 1994, 269, 9889–9897. [Google Scholar] [PubMed]
- Benzie, I.F. Lipid peroxidation: A review of causes, consequences, measurement and dietary influences. Int. J. Food Sci. Nutr. 1996, 47, 233–261. [Google Scholar] [CrossRef] [PubMed]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Ortwerth, B.J.; James, H.; Simpson, G.; Linetsky, M. The generation of superoxide anions in glycation reactions with sugars, osones, and 3-deoxyosones. Biochem. Biophys. Res. Commun. 1998, 245, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Carubelli, R.; Schneider, J.E.; Pye, Q.N.; Floyd, R.A. Cytotoxic effects of autoxidative glycation. Free Radic. Biol. Med. 1995, 18, 265–269. [Google Scholar] [CrossRef]
- Limon-Pacheco, J.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. 2009, 674, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Friedman, W.J. Cytokines regulate expression of the type 1 interleukin-1 receptor in rat hippocampal neurons and glia. Exp. Neurol. 2001, 168, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Tchelingerian, J.L.; Le Saux, F.; Jacque, C. Identification and topography of neuronal cell populations expressing TNFα and IL-1α in response to hippocampal lesion. J. Neurosci. Res. 1996, 43, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Barger, S.W.; Liu, L.; Mrak, R.E.; Griffin, W.S. S100β induction of the proinflammatory cytokine interleukin-6 in neurons. J. Neurochem. 2000, 74, 143–150. [Google Scholar] [PubMed]
- Amor, S.; Puentes, F.; Baker, D.; van der Valk, P. Inflammation in neurodegenerative diseases. Immunology 2010, 129, 154–169. [Google Scholar] [CrossRef] [PubMed]
- Dukic-Stefanovic, S.; Gasic-Milenkovic, J.; Deuthe-Conrad, W.; Münch, G. Signal transduction pathways in mouse microglia N-11 cells activated by advanced glycation endproducts (AGEs). J. Neurochem. 2003, 87, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Gasic‐Milenkovic, J.; Dukic‐Stefanovic, S.; Deuther‐Conrad, W.; Gärtner, U.; Münch, G. β-amyloid peptide potentiates inflammatory responses induced by lipopolysaccharide, interferon -γ and “advanced glycation endproducts” in a murine microglia cell line. Eur. J. Neurosci. 2003, 17, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.; Schinzel, R.; Palm, D.; Riederer, P.; Münch, G. High molecular weight hyaluronic acid inhibits advanced glycation endproduct-induced NF-κB activation and cytokine expression. FEBS Lett. 1999, 453, 283–287. [Google Scholar] [CrossRef]
- Ardestani, A.; Yazdanparast, R. Inhibitory effects of ethyl acetate extract of Teucrium polium on in vitro protein glycoxidation. Food Chem. Toxicol. 2007, 45, 2402–2411. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (mtt): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in mtt reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Nachlas, M.M.; Margulies, S.I. The determination of lactic dehydrogenase with a tetrazolium salt. Anal. Biochem. 1960, 1, 317–326. [Google Scholar] [CrossRef]
- Sample Availability: Samples are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amornrit, W.; Santiyanont, R. Effect of Amaranthus on Advanced Glycation End-Products Induced Cytotoxicity and Proinflammatory Cytokine Gene Expression in SH-SY5Y Cells. Molecules 2015, 20, 17288-17308. https://doi.org/10.3390/molecules200917288
Amornrit W, Santiyanont R. Effect of Amaranthus on Advanced Glycation End-Products Induced Cytotoxicity and Proinflammatory Cytokine Gene Expression in SH-SY5Y Cells. Molecules. 2015; 20(9):17288-17308. https://doi.org/10.3390/molecules200917288
Chicago/Turabian StyleAmornrit, Warisa, and Rachana Santiyanont. 2015. "Effect of Amaranthus on Advanced Glycation End-Products Induced Cytotoxicity and Proinflammatory Cytokine Gene Expression in SH-SY5Y Cells" Molecules 20, no. 9: 17288-17308. https://doi.org/10.3390/molecules200917288
APA StyleAmornrit, W., & Santiyanont, R. (2015). Effect of Amaranthus on Advanced Glycation End-Products Induced Cytotoxicity and Proinflammatory Cytokine Gene Expression in SH-SY5Y Cells. Molecules, 20(9), 17288-17308. https://doi.org/10.3390/molecules200917288