
Proteome and Peptidome of Vipera berus berus Venom


 , ,
, ,
Abstract
:1. Introduction
2. Results
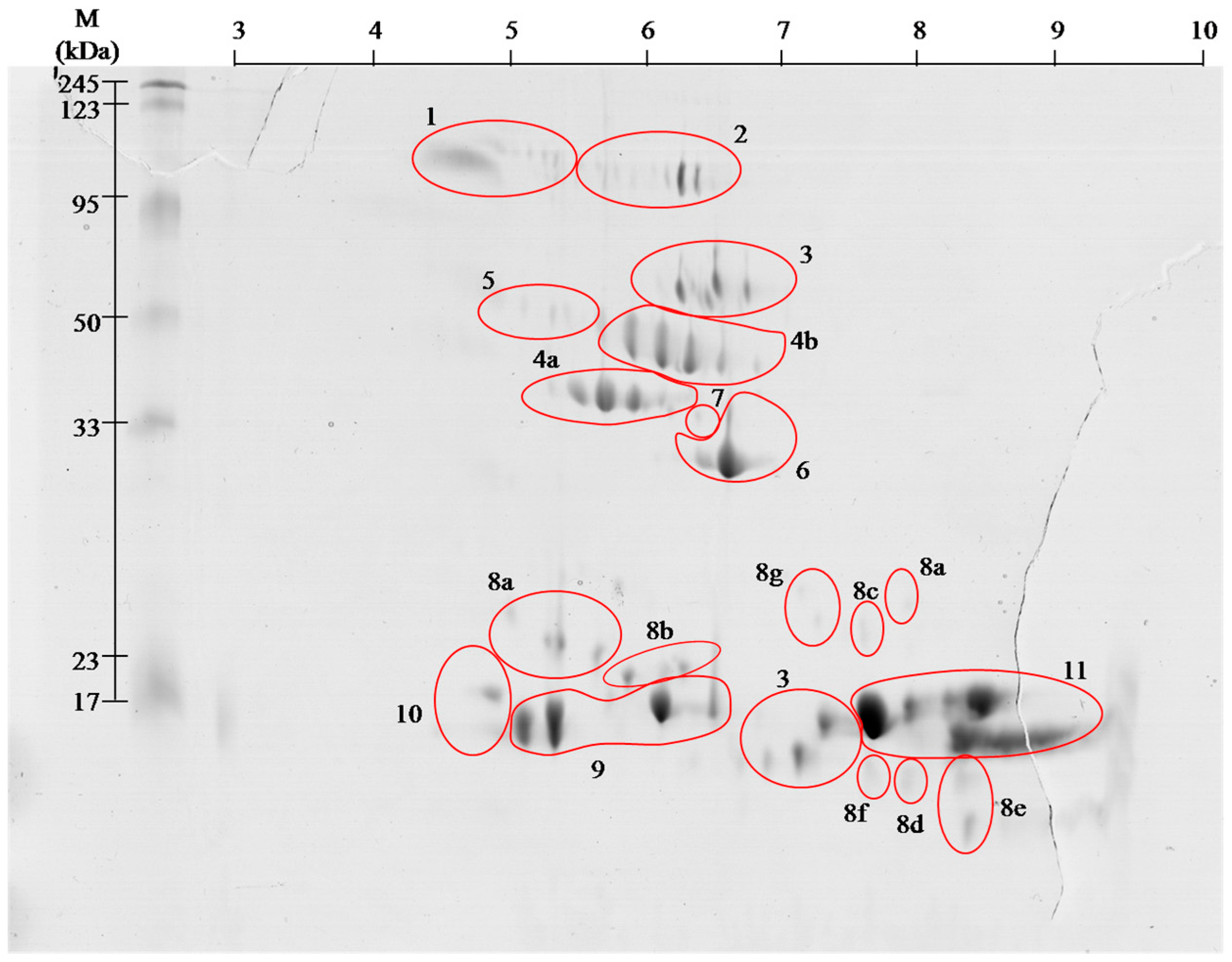
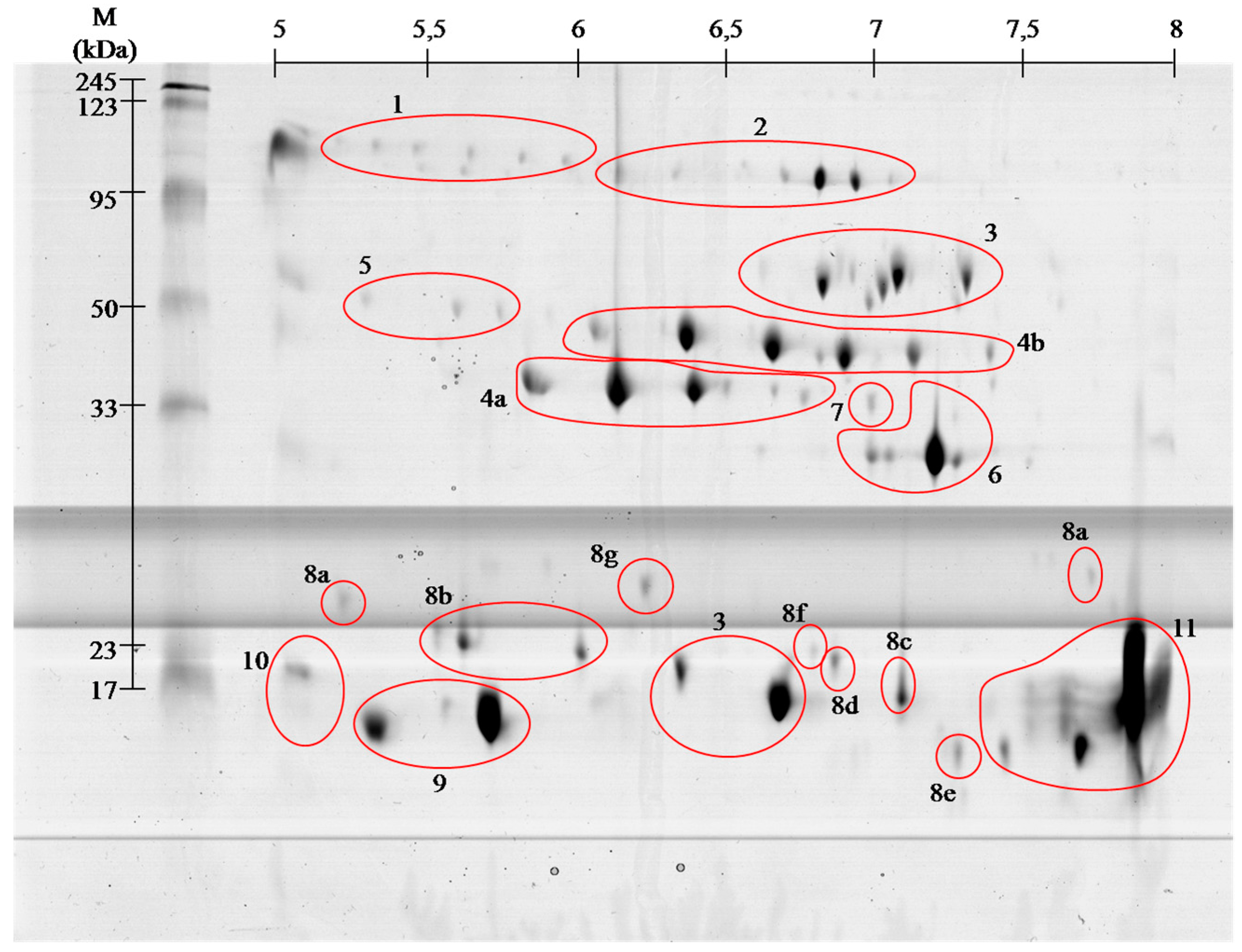
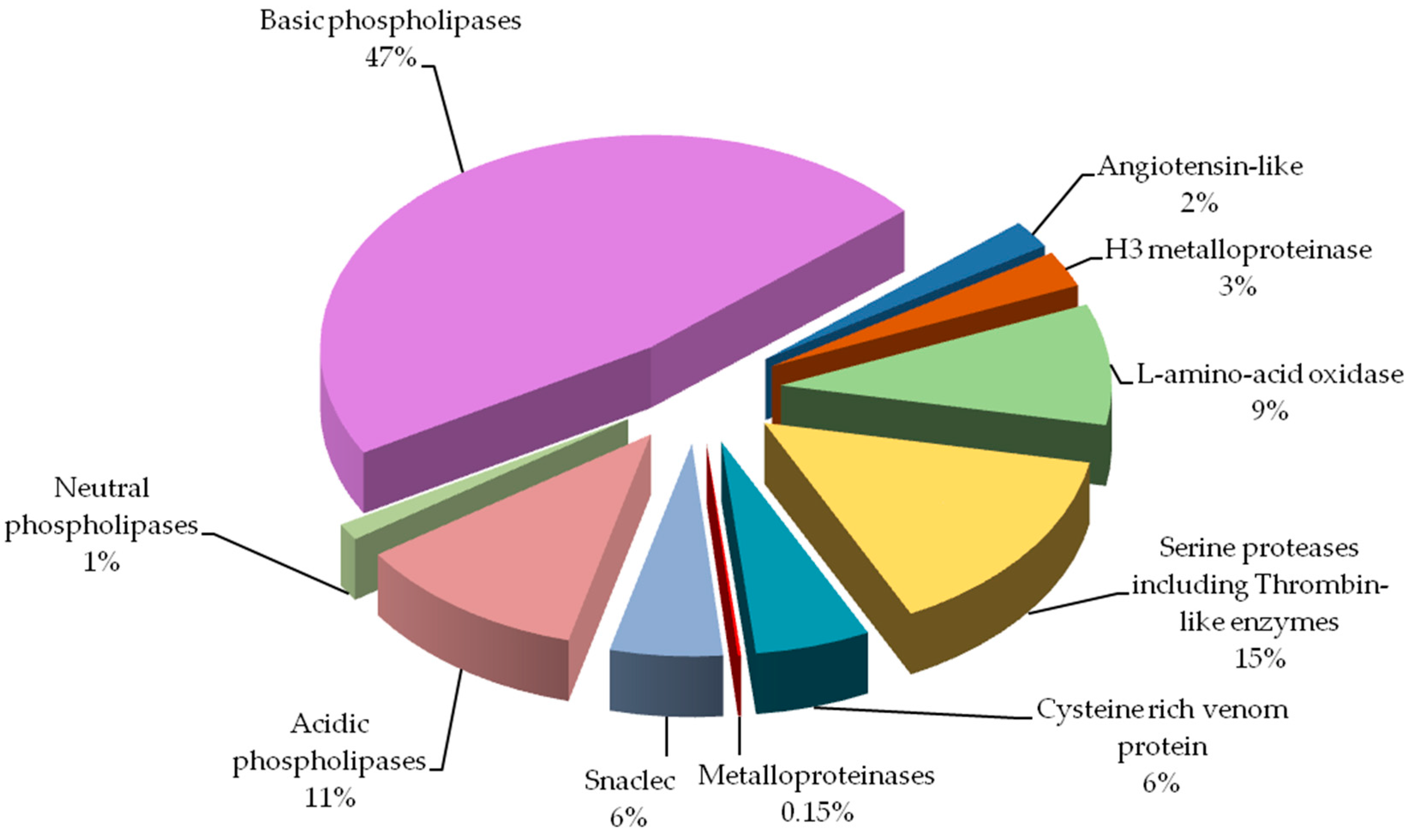
2.1. Proteome
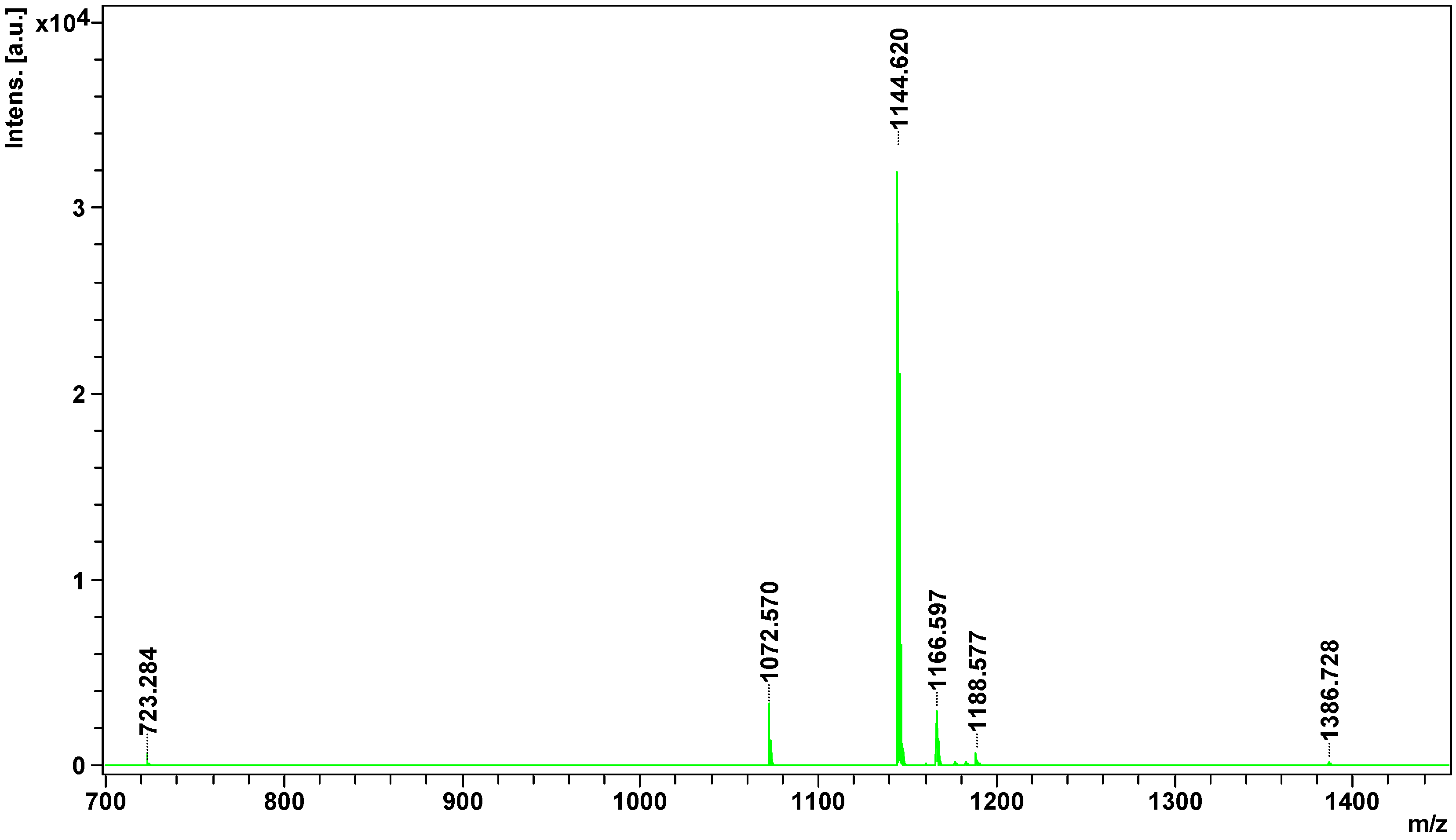
2.2. Peptidome
3. Discussion
4. Materials and Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koh, D.C.I.; Armugam, A.; Jeyaseelan, K. Snake venom components and their applications in biomedicine. Cell. Mol. Life Sci. 2006, 63, 3031–3041. [Google Scholar] [CrossRef] [PubMed]
- Mallow, D.; Ludwig, D.; Nilson, G. True Vipers: Natural History and Toxinology of Old World Vipers; Krieger Publishing Company: Malabar, FL, USA, 2003; p. 410. [Google Scholar]
- Czajka, U.; Wiatrzyk, A.; Lutyńska, A. Mechanizm działania jadu żmii zygzakowatej i zasady zastosowania antytoksyny w leczeniu. Przegl. Epidemiol. 2013, 67, 729–733. [Google Scholar]
- Palviainen, M.; Raekallio, M.; Vainionpää, M.; Kosonen, S.; Vainio, O. Proteomic profiling of dog urine after European adder (Vipera berus berus) envenomation by two-dimensional difference gel electrophoresis. Toxicon 2012, 60, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J.; Kini, R.M. Introduction. In Toxins and Hemostasis; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 1–9. [Google Scholar]
- Perchuc, A.M.; Wilmer, M. Diagnostic use of snake venom components in the coagulation laboratory. In Toxins and Hemostasis; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 747–766. [Google Scholar]
- Rojnuckarin, P. Snakebite-Induced Coagulopathy and Bleeding Disorders. In Toxins and Hemostasis; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 699–710. [Google Scholar]
- Latinović, Z.; Leonardi, A.; Šribar, J.; Sajevic, T.; Žužek, M.C.; Frangež, R.; Halassy, B.; Trampuš-Bakija, A.; Pungerčar, J.; Križaj, I. Venomics of Vipera berus berus to explain differences in pathology elicited by Vipera ammodytes ammodytes envenomation: Therapeutic implications. J. Proteom. 2012, 146, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Vejayan, J.; Shin Yee, L.; Ponnudurai, G.; Ambu, S.; Ibrahim, I. Protein profile analysis of Malaysian snake venoms by two-dimensional gel electrophoresis. J. Venom Anim. Toxins Incl. Trop. Dis. 2010, 16, 623–630. [Google Scholar] [CrossRef]
- Serrano, S.M.T.; Shannon, J.D.; Wang, D.; Camargo, A.C.M.; Fox, J.W. A multifaceted analysis of viperid snake venoms by two-dimensional gel electrophoresis: An approach to understanding venom proteomics. Proteomics 2005, 5, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Vejayan, J.; Tang, M.S.; Halijah, I. The role of conventional two-dimensional electrophoresis (2DE) and its newer applications in the study of snake venoms. In Proteomic Applications in Biology; Heazlewood, J.L., Petzold, C.J., Eds.; IN TECH: Rijeka, Croatia, 2012; pp. 226–252. [Google Scholar]
- Du, X.-Y.; Clemetson, K.J. Reptile C-type lectins. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 359–376. [Google Scholar]
- Nevalainen, T.J.; Graham, G.G.; Scott, K.F. Antibacterial actions of secreted phospholipases A2. Biochim. Biophys. Acta 2008, 1781, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Doley, R.; Zhou, X.; Kini, R.M. Snake venom phospholipase A2 enzymes. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 173–206. [Google Scholar]
- Phillips, D.J.; Swenson, S.D.; Markland, F.S., Jr. Thrombin-like Snake Venom Serine Proteinases. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 139–154. [Google Scholar]
- Sweetnam, P.M.; Thomas, H.F.; Yarnell, J.W.; Beswick, A.D.; Baker, I.A.; Elwood, P.C. Fibrinogen, viscosity and the 10-year incidence of ischaemic heart disease: The Caerphilly and Speedwell studies. Eur. Heart J. 1996, 17, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.W.; Serrano, S.M.T. Snake Venom Metalloproteinases. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 95–114. [Google Scholar]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T. Snake venom metalloproteinases: Biological roles and participation in the pathophysiology of envenomation. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 115–138. [Google Scholar]
- Hsu, C.-C.; Huang, T.-F. Biological activities of snake venom metalloproteinases on platelets, neutrophils, endothelial cells, and extracellular matrices. In Toxins and Hemostasis; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 723–732. [Google Scholar]
- Borgheresi, R.A.M.B.; Dalle Lucca, J.; Carmona, E.; Picarelli, Z.P. Isolation and identification of angiotensin-like peptides from the plasma of the snake Bothrops jararaca. Comp. Biochem. Physiol. 1996, 113, 467–473. [Google Scholar] [CrossRef]
- Du, X.Y.; Clemetson, K.J. Snake venom l-amino acid oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef]
- Sun, M.-Z.; Guo, C.; Tian, X.; Chen, D.; Greenaway, F.T.; Liu, S. Biochemical, functional and structural characterization of Akbu-LAAO: A novel snake venom l-amino acid oxidase from Agkistrodon blomhoffii ussurensis. Biochimie 2010, 92, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Escolano, J.; Fernandez, J.; Sanz, L.; Angulo, Y.; Gutierrez, J.M.; Calvete, J.J. Snake venomics and antivenomics of the arboreal neotropical pitvipers Bothriechis lateralis and Bothriechis schlegelii. J. Proteome Res. 2008, 7, 2445–2457. [Google Scholar] [CrossRef] [PubMed]
- Stabeli, R.G.; Marcussi, S.; Carlos, G.B.; Pietro, R.C.; Selistre-de-Araujo, H.S.; Giglio, J.R.; Oliveira, E.B.; Soares, A.M. Platelet aggregation and antibacterial effects of an l-amino acid oxidase purified from Bothrops alternatus snake venom. Bioorg. Med. Chem. 2004, 12, 2881–2886. [Google Scholar] [CrossRef] [PubMed]
- Samel, M.; Vija, H.; Ronnholm, G.; Siigur, J.; Kalkkinen, N.; Siigur, E. Isolation and characterization of an apoptotic and platelet aggregation inhibiting l-amino acid oxidase from Vipera berus berus (common viper) venom. Biochim. Biophys. Acta 2006, 4, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Stiles, B.G.; Sexton, F.W.; Weinstein, S.A. Antibacterial effects of different snake venoms: Purification and characterization of antibacterial proteins from Pseudechis australis (Australian king brown or mulga snake) venom. Toxicon 1991, 29, 1129–1141. [Google Scholar] [CrossRef]
- Ramazanova, A.S.; Starkov, V.G.; Osipov, A.V.; Ziganshin, R.H.; Filkin, S.Y.; Tsetlin, V.I.; Utkin, Y.N. Cysteine-rich venom proteins from the snakes of Viperinae subfamily-molecular cloning and phylogenetic relationship. Toxicon 2009, 53, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Calderon, L.; Lomonte, B.; Gutierrez, J.M.; Tarkowski, A.; Hanson, L.A. Biological and biochemical activities of Vipera berus (European viper) venom. Toxicon 1993, 31, 743–753. [Google Scholar] [CrossRef]
- Reading, C.J. Incidence, pathology, and treatment of adder (Vipera berus L.) bites in man. J. Accid. Emerg. Med. 1996, 13, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Terra, R.M.S.; Pinto, A.F.M.; Guimaraes, J.A.; Fox, J.W. Proteomic profiling of snake venom metalloproteinases (SVMPs): Insights into venom induced pathology. Toxicon 2009, 54, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Siigur, E.; Siigur, J.; Nômmeots, M.; Ilomets, T. Fractionation and enzymatic activities of common viper (Vipera berus berus) venom. Toxicon 1979, 17, 623–630. [Google Scholar] [CrossRef]
- Munawar, A.; Zahid, A.; Negm, A.; Akrem, A.; Spencer, P.; Betzel, C. Isolation and characterization of Bradykinin potentiating peptides from Agkistrodon bilineatus venom. Proteome Sci. 2016, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Munawar, A.; Trusch, M.; Georgieva, D.; Spencer, P.; Frochaux, V.; Harder, S.; Arni, R.K.; Duhalov, D.; Genov, N.; Schlüter, H.; et al. Venom peptide analysis of Vipera ammodytes meridionalis (Viperinae) and Bothrops jararacussu (Crotalinae) demonstrates subfamily-specificity of the peptidome in the family Viperidae. Mol. Biosyst. 2011, 7, 3298–3307. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, A.M.; De Lima, M.E. Small peptides, big world: Biotechnological potential in neglected bioactive peptides from arthropod venoms. J. Pept. Sci. 2005, 11, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Hochstrasser, D.F.; Harrringto, M.G.; Hochstrasser, A.C.; Miller, M.J.; Meril, C.R. Methods for increasing the resolution of two-dimensional protein electrophoresis. Anal. Biochem. 1988, 73, 424–435. [Google Scholar] [CrossRef]
- Neuhoff, V.; Arold, N.; Taube, D.; Ehrhardt, W. Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis 1988, 9, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Vizcaíno, J.A.; Csordas, A.; del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and related tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples are not available from the authors.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. # | Identified Protein & | Accession * | Organism ¥ | Mass (kDa) £ | S ± | Peptide Sequence ≠ |
|---|---|---|---|---|---|---|
| 1 | Angiotensin- like peptide 2 | ANGT2_BOTJA | Bothrops jararaca | 1.04 | 44 | DRVYIHPF (1046.5816) |
| 2 | H3 metalloproteinase precursor 1 | VM1H3_DEIAC | Vipera ammotydes ammotydes | 70.7 | 48 | NPCQIYYTPR (1311.7225) |
| 3 | l-amino acid oxidase | OXLA_BOTPC | Bothrops pictus | 56.7 | 102 | SAGQLYEESLR (1252.6349) |
| OXLA_BOTCO | Bothrops cotiara | 1.8 | 49 | NPLEECFR (1252.6575) | ||
| OXLA_MACLB | Macrovipera lebetina | 12.5 | 58 | PMF SC 40.2% | ||
| OXLA_BOTJR | Bothrops jararacussu | 56.7 | 59 | FWEDDGIHGGK (1260.6088) | ||
| OXLA_VIPBB | Vipera berus berus | 10.3 | 68 | ADDICNPLEECFR (1493.7661) | ||
| OXLA_BOTJA | Bothrops jararacussu | 56.5 | 50 | HIDDIFAYEK (1137.5991) | ||
| OXLA_BOTPC | Bothrops pictus | 56.7 | 101 | SAGQLYEESLR (1252.7064) | ||
| OXLA_BOTIN | Bothrops insularis | 5.3 | 37 | ADDKNPLEECFR (1493.7653) | ||
| 4a | Serine protease VLSp-3 | VSP3_MACLB | Macrovipera lebetina | 29 | 25 | TSTHIAPLSLPSSPPSVGSVCR (2250.4505) |
| 4b | Snake venom serine protease nikobin | VSP_VIPNI | Vipera nikolskii | 28.8 | 64 | PMF SC 25.7% |
| 53 | CQGVHPELPAK (1235.6438) | |||||
| 55 | VVCAGIWQGGK (1174.6110) | |||||
| 96 | VILPDVPHCANIEIIK (1831.0636) | |||||
| 36 | EYTMWDK (972.4713) | |||||
| 5 | Beta- fibrinogenase brevinase | VSPB_GLOBL | Gloydius blomhoffi | 26.3 | 56 | VIGGDECNINEHR (1512.7806) |
| VSPBF_MACLB | Macrovipera lebetina | 28.2 | 26 | FFCLSSK (888.4284) | ||
| Thrombin-like enzyme crotalase | VSPCR_CROAD | Crotalus adamanteus | 30.1 | 36 | WDKDIMLIR (1189.6561) | |
| Venom serine proteinase-like protein 2 | VSP2_MACLB | Macrovipera lebetina | 29.5 | 70 | IMGWGTITTTK (1208.6420) | |
| 134 | TLCAGILQGGIDSCK (1592.7767) | |||||
| 6 | Cysteine rich venom protein | CRVP_VIPBE | Vipera berus | 27.4 | 73 | MEWYPEAAANAER (1537.6894) |
| 53 | PMF SC 24.7% | |||||
| 74 | PMF SC 36.4% | |||||
| 51 | KPEIQNEIIDLHNSLR (1919.0970) | |||||
| 7 | Snake venom metalloproteinase VMP1 | VM1V1_AGKPL | Agkistrodon piscivorus leucostoma | 47.1 | 58 | NPQCILNKPLR (1352.7017) |
| Zinc metalloproteinase/disintegrin | VM2L2_MACLB | Macrovipera lebetina | 47.1 | 58 | NPQCILNQPL (1352.7017) | |
| 8a | Snaclec rhinocetin submit beta | SLRB_BITRH | Bitis rhinoceros | 18.7 | 59 | TTDNQWLR (1033.6040) |
| 8b | Snaclec A14 | SLAE_MACLB | Macrovipera lebetina | 18 | 39 | TSADYVWIGLWNQR (1708.9104) |
| 8c | Snaclec B6 | SLB6_MACLB | Macrovipera lebetina | 15.3 | 43 | ANLVWIGLR (1041.6803) |
| 8d | Snaclec echicetin subunit alpha | SLA_ECHCA | Echis carinatus | 16.1 | 37 | TWDEAEKFCNK (1427.6446) |
| 8e | Snaclec 1 | SL1_ECHCS | Echis carinatus sochureki | 17.1 | 35 | GSHLVSLHNIAEADFVVK (1936.0465) |
| 8f | Snaclec rhodocetin subunit alpha | SLEA_CALRH | Calloselasma rhodostoma | 15.9 | 47 | TWEEAER (920.46147) |
| 8f | Snaclec A13 | SLAD_MACLB | Macrovipera lebetina | 15.6 | 26 | DQDCLPGWSFYEGHCYK (2161.9310) |
| 8g | Snaclec jerdonibitin submit alpha | SLA_PROJR | Protobothrops jerdoni | 18.1 | 45 | TWEDAER (906.4700) |
| 9 | Acidic phospholipase ammodytin I1 | PA2A1_VIPAA | Vipera ammodytes ammodytes | 16.2 | 56 | PMF SC 30.4% |
| Acidic phospholipase A2 PLA-1 | PA2A1_ERIMA | Eristicophis macmahoni | 14.3 | 58 | PMF SC 36.4% | |
| Acidic phospholipase A2 PL1 | PA2A1_VIPRE | Viprea renardi | 16.2 | 72 | CCFVHDCCYGR (1533.4506) | |
| 10 | Neutral phospholipase A2 ammodytoxin I2 | PA2N_DABRR | Vipera ammodytes ammodytes | 16.1 | 56 | PMF SC 30% |
| 11 | Basic phospholipase A2 ammodytoxin C | PA2BA_VIPAA | Vipera ammodytes ammodytes | 15.4 | 28 | AAAICFR (808.4193) |
| Basic phospholipase A2 Pla2Vb | PA2B_VIPBB | Vipera berus berus | 15.7 | 48 | HICECDR (989.4389) | |
| Basic phospholipase A2 vurtoxin | PA2B_VIPRE | Vipera renardi | 16.4 | 31 | YYPDFLCK (1105.5024) | |
| Basic phospholipase A2 | PA2B3_DABRR | Daboia russelii | 14.4 | 142 | CCFVHDCCYGNLPDCNPK (2315.8464) |
| Parent Ion m/z | Identified Peptide & | Accession * | Organism ¥ | Peptide Sequence ≠ | Mass (Da) £ | S ± |
|---|---|---|---|---|---|---|
| 1072.5704 | Bradykinin potentiating peptide D | gi 229282 | Gloydius blomhoffii | EGLPPRPIPP | 1072 | 14 |
| 1144.6198 | Bradykinin-potentiating peptide S1,2,4 | BPP4_BOTIN | Bothrops insularis | QGGPPRPQIPP + Deamidated (NQ) | 1143 | 31 |
| 1166.5968 | Bradykinin-potentiating peptide 11a | BP11A_CROVV | Crotalus viridis viridis | QGPSPRHPIPP + Gln→pyro-Glu (N-term Q); Deamidated (NQ) | 1182 | 5 |
| 1182.5725 | Bradykinin-potentiating peptide 10d | BPPAD_BOTCO | Bothrops cotiara | QNWPHPPMPP + Gln→pyro-Glu (N-term Q) | 1200 | 9 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bocian, A.; Urbanik, M.; Hus, K.; Łyskowski, A.; Petrilla, V.; Andrejčáková, Z.; Petrillová, M.; Legath, J. Proteome and Peptidome of Vipera berus berus Venom. Molecules 2016, 21, 1398. https://doi.org/10.3390/molecules21101398
Bocian A, Urbanik M, Hus K, Łyskowski A, Petrilla V, Andrejčáková Z, Petrillová M, Legath J. Proteome and Peptidome of Vipera berus berus Venom. Molecules. 2016; 21(10):1398. https://doi.org/10.3390/molecules21101398
Chicago/Turabian StyleBocian, Aleksandra, Małgorzata Urbanik, Konrad Hus, Andrzej Łyskowski, Vladimír Petrilla, Zuzana Andrejčáková, Monika Petrillová, and Jaroslav Legath. 2016. "Proteome and Peptidome of Vipera berus berus Venom" Molecules 21, no. 10: 1398. https://doi.org/10.3390/molecules21101398
APA StyleBocian, A., Urbanik, M., Hus, K., Łyskowski, A., Petrilla, V., Andrejčáková, Z., Petrillová, M., & Legath, J. (2016). Proteome and Peptidome of Vipera berus berus Venom. Molecules, 21(10), 1398. https://doi.org/10.3390/molecules21101398