
Effects of Flavonoids in Lysimachia clethroides Duby on the Activities of Cytochrome P450 CYP2E1 and CYP3A4 in Rat Liver Microsomes


Abstract
:1. Introduction
2. Results
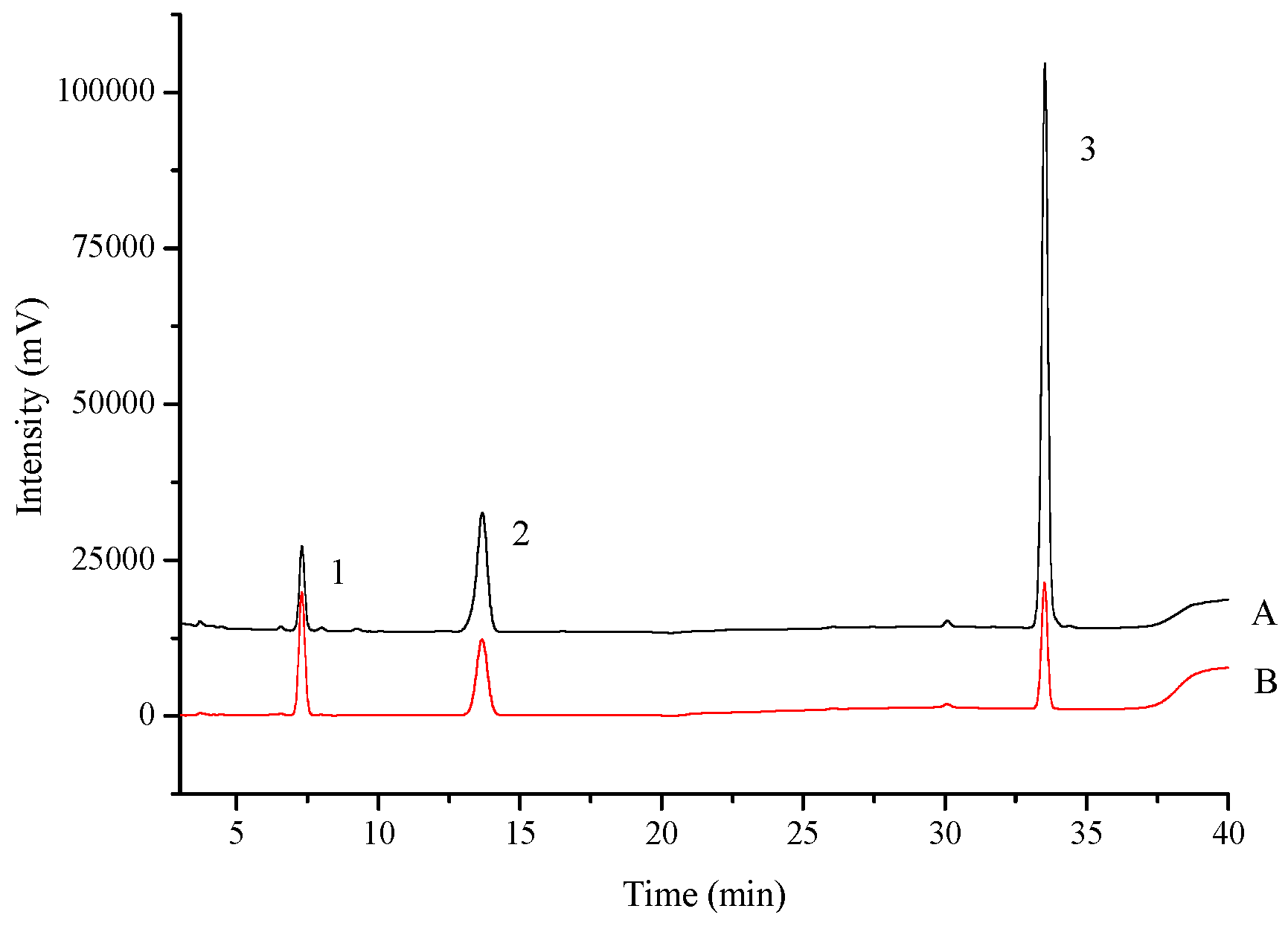
2.1. Method Validation
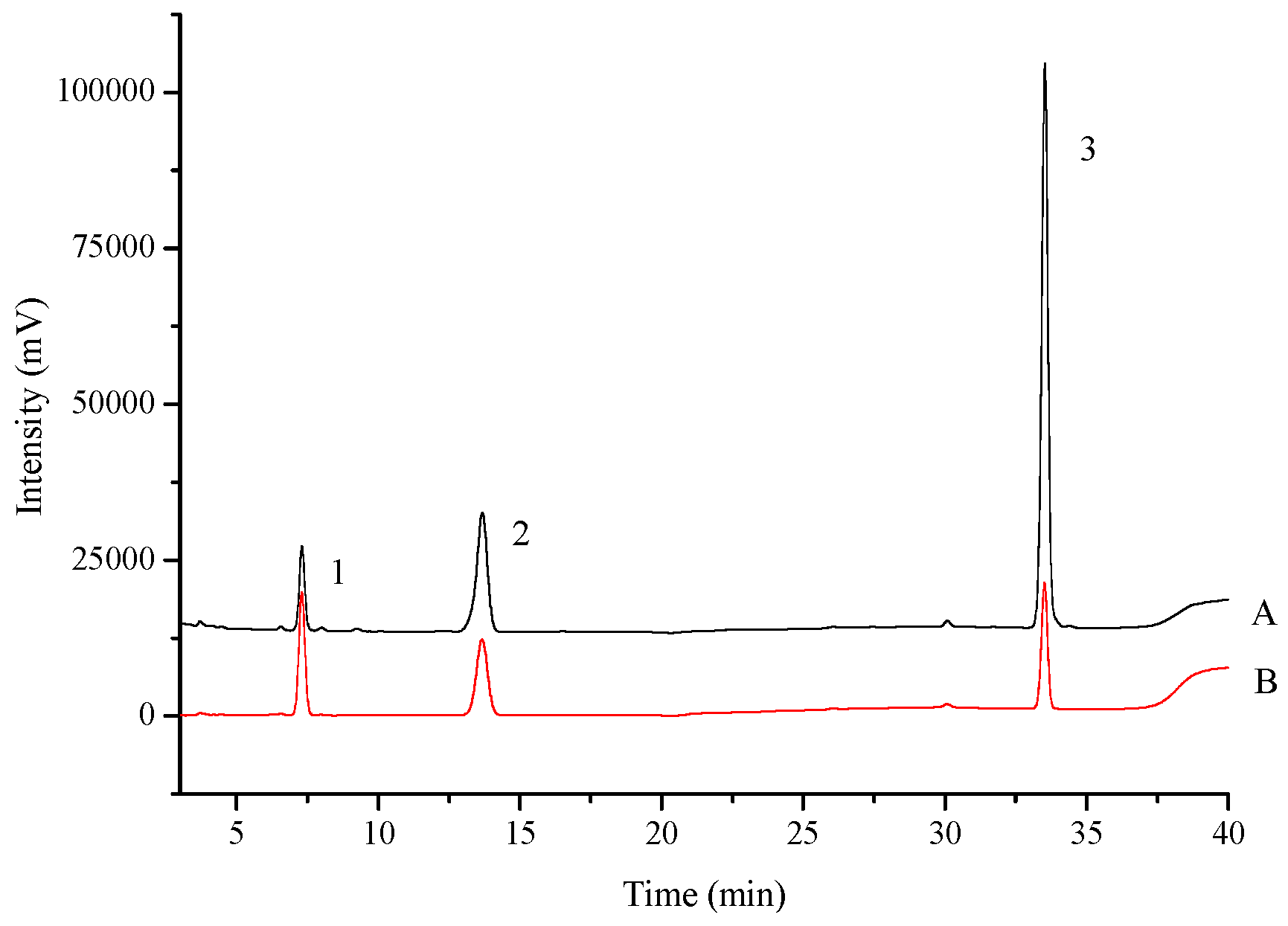
2.1.1. Standard Curves and the Linearity
2.1.2. Precision and Stability
2.1.3. Recovery
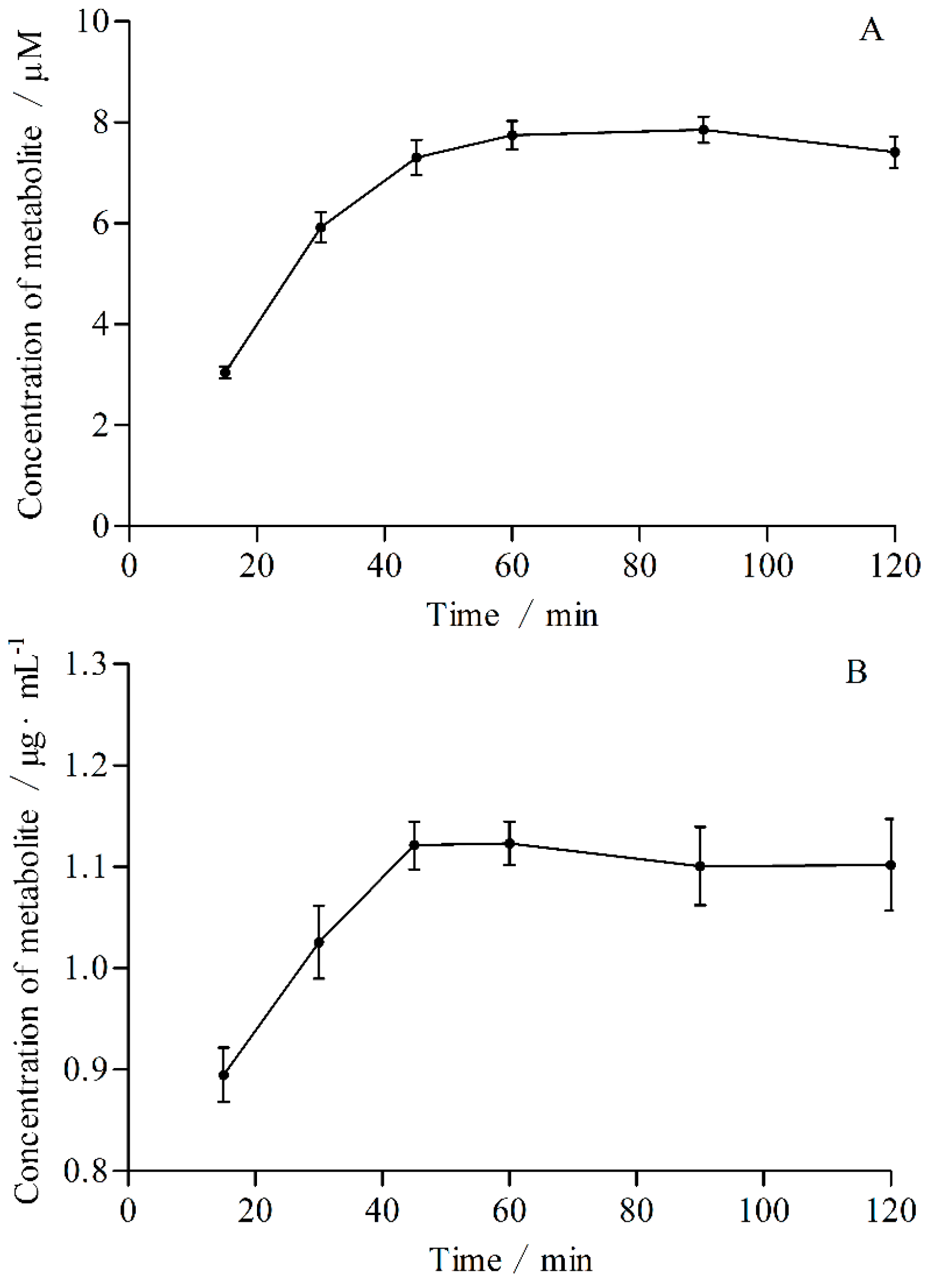
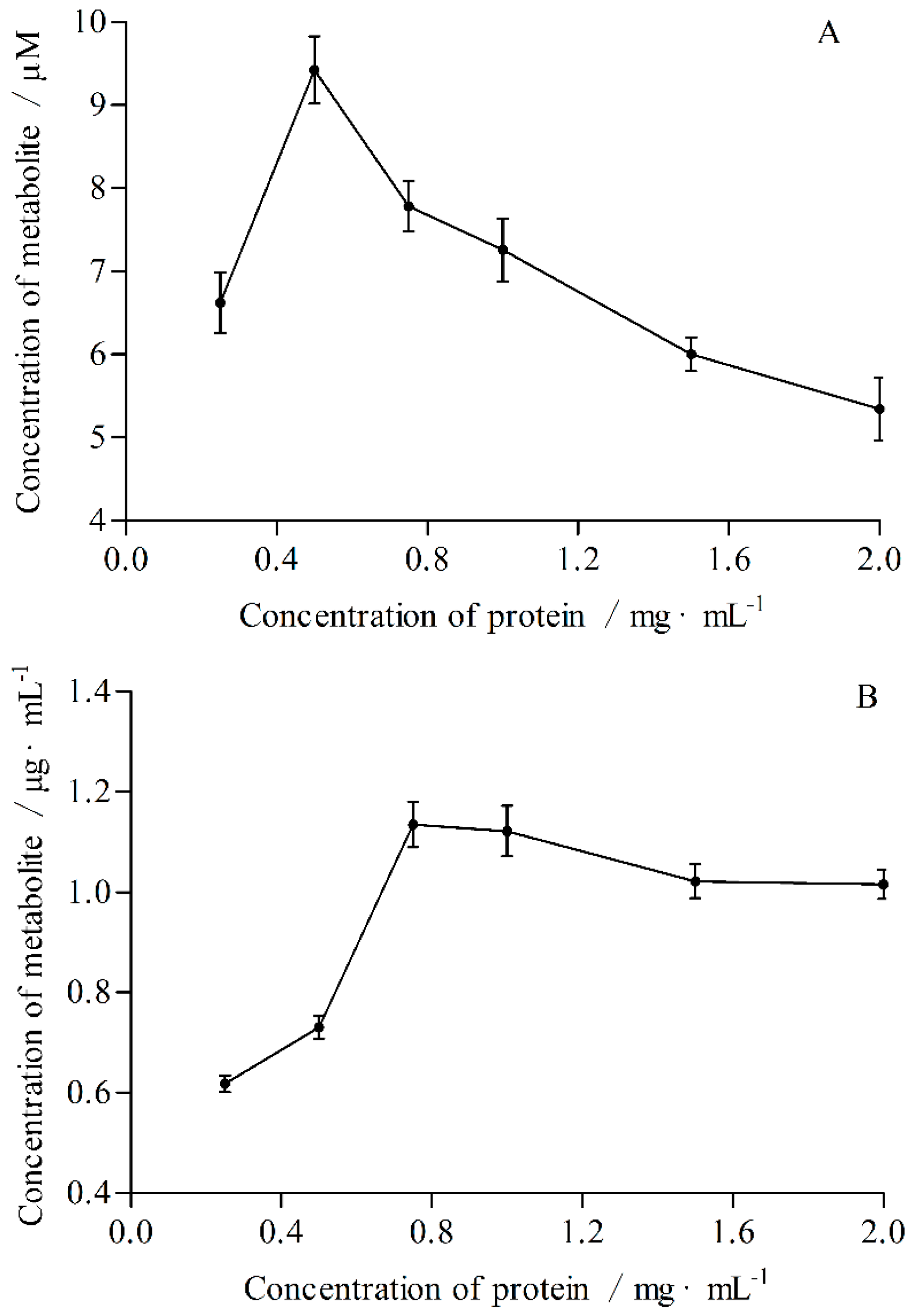
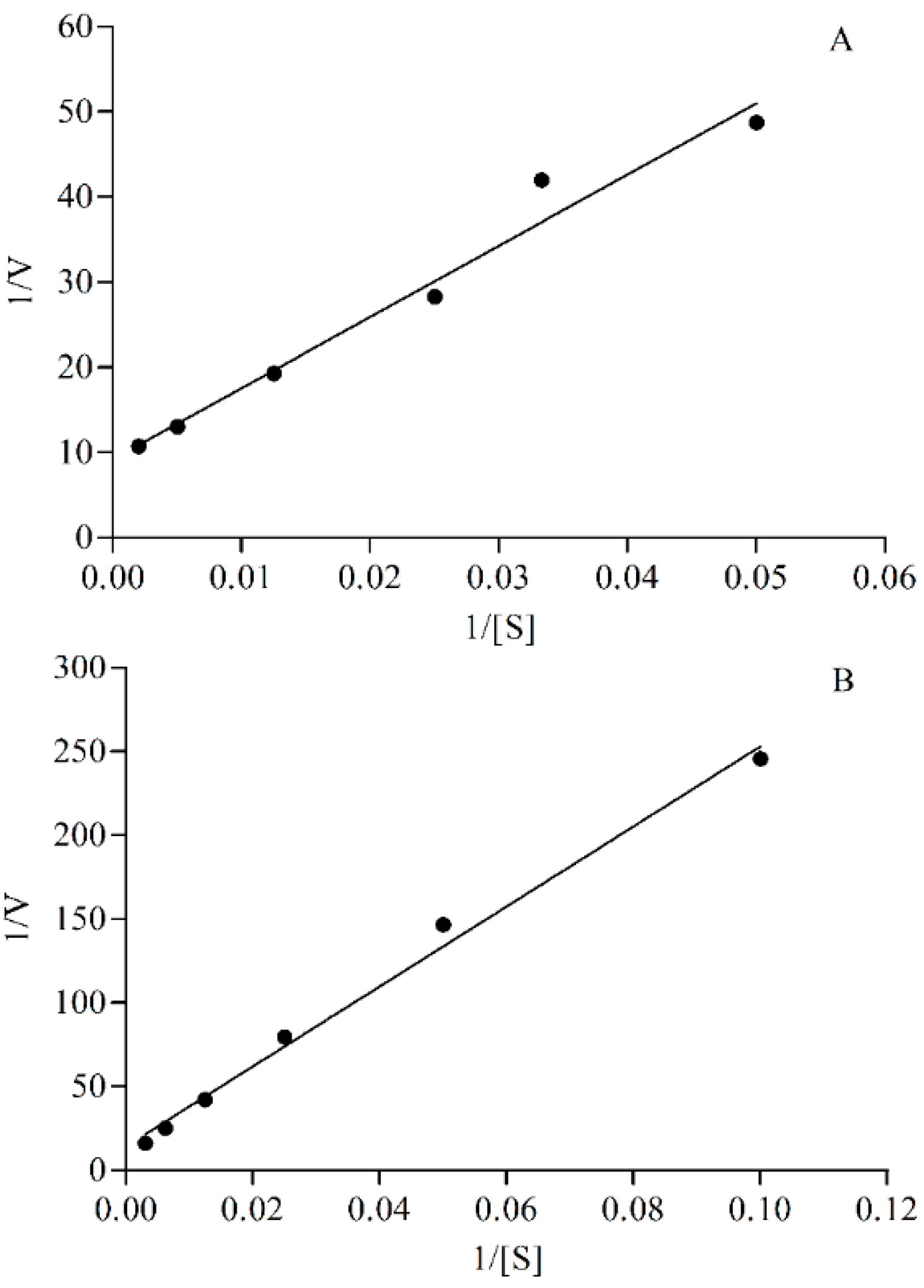
2.2. The Optimal Conditions of Incubation System in Vitro
2.3. Effects of Flavonoids in L. clethroides on CYP450 Activity
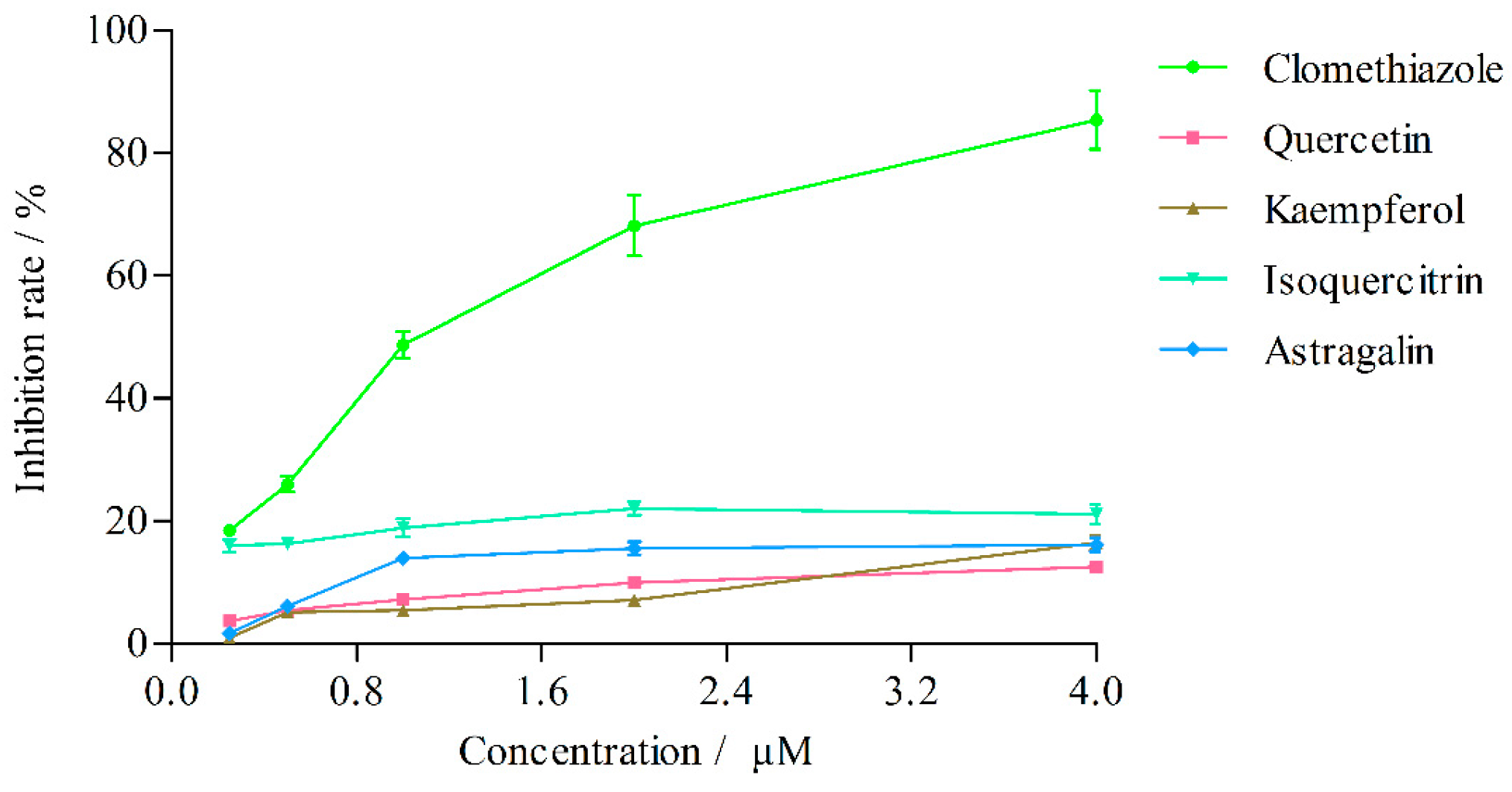
2.3.1. Effects on CYP2E1 Activity
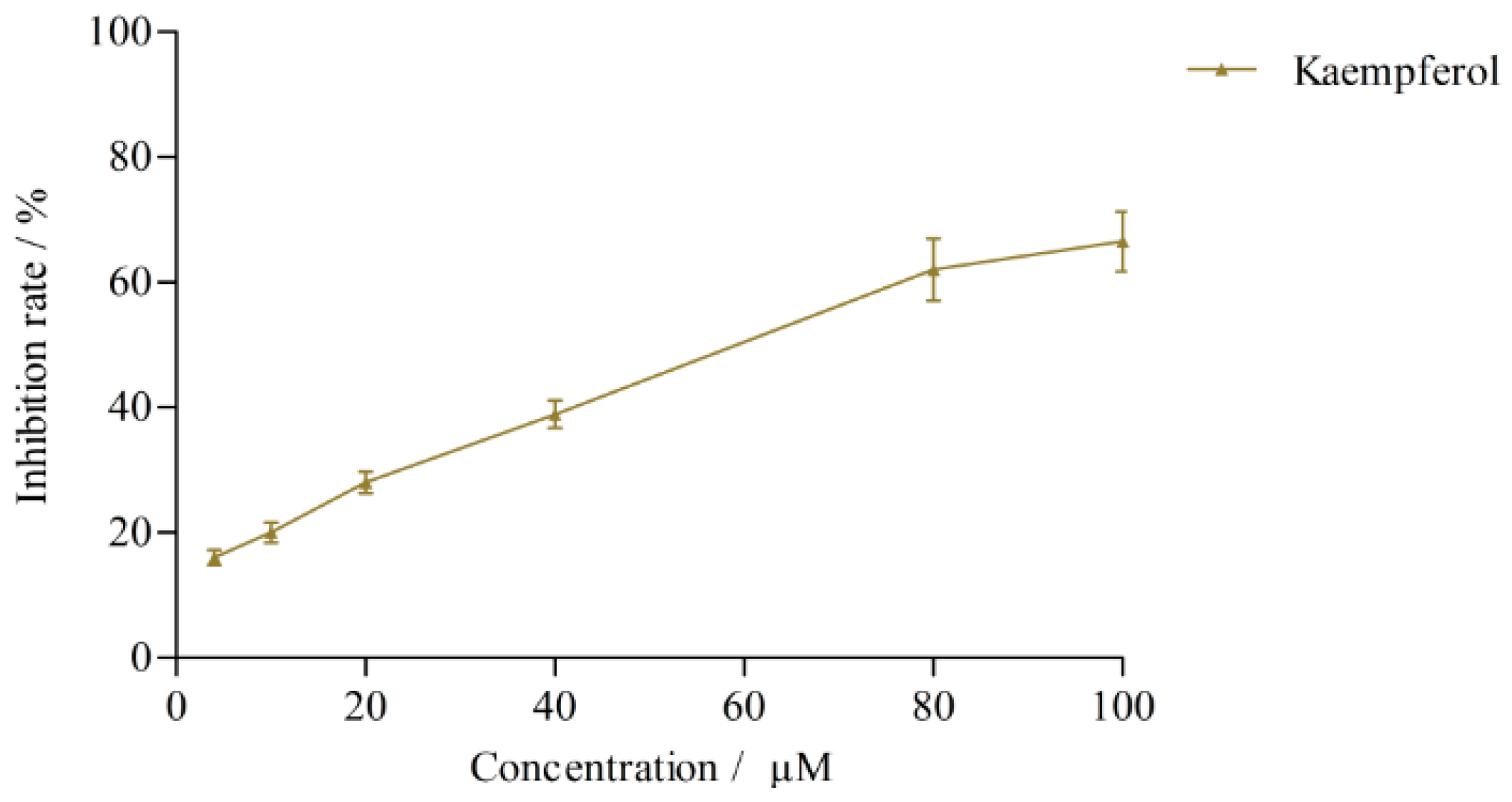
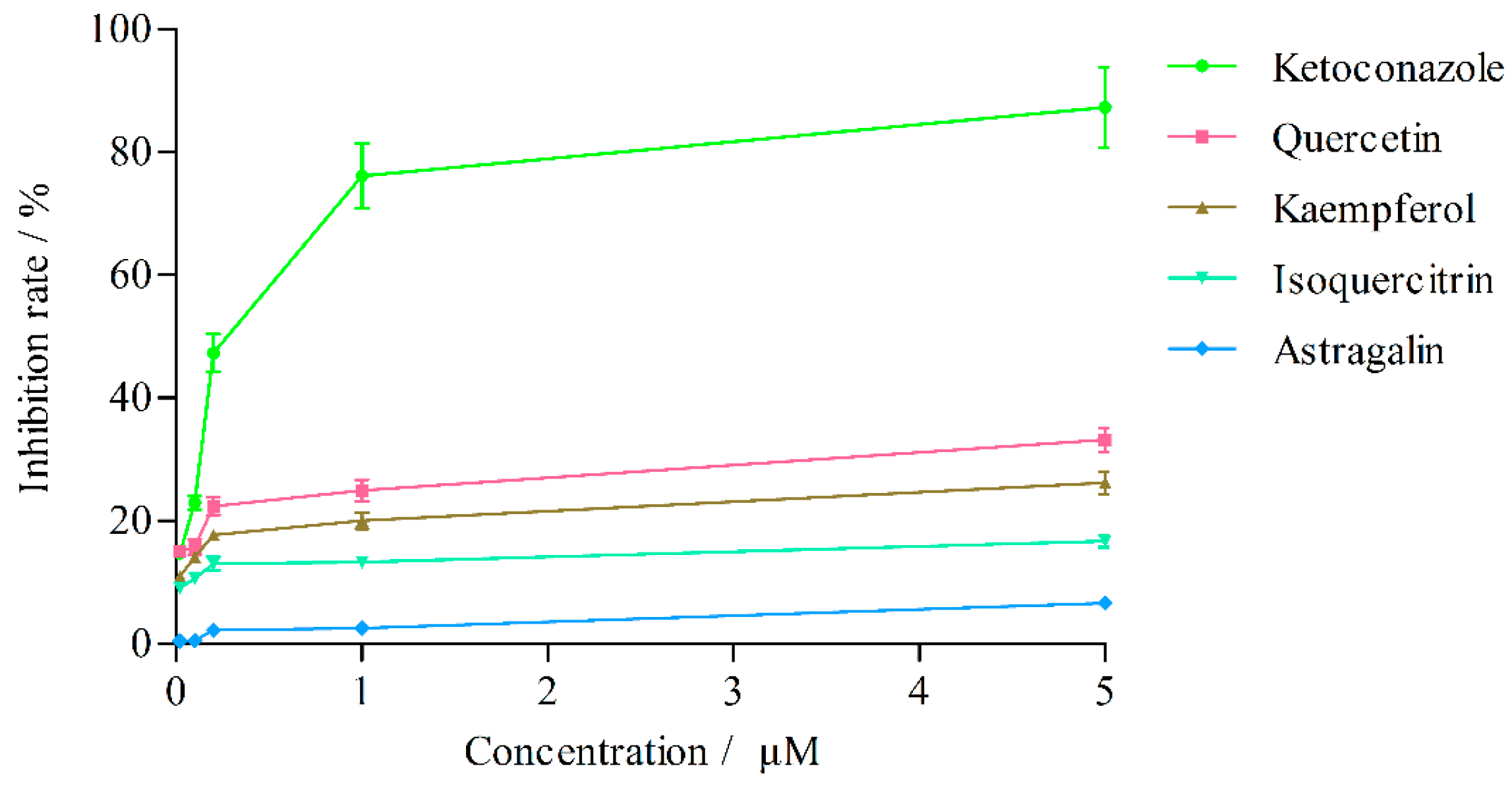
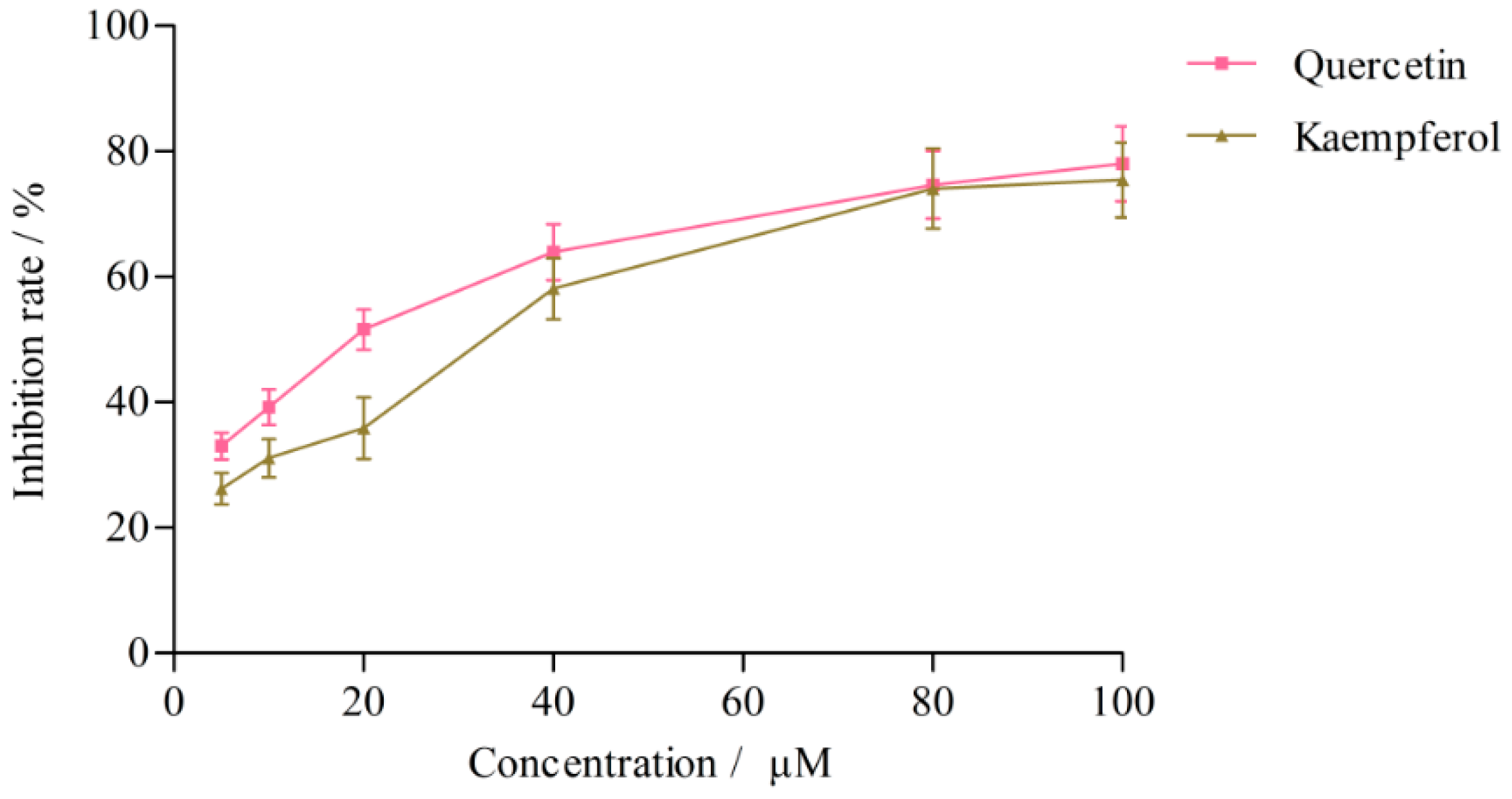
2.3.2. Effects on CYP3A4 Activity
3. Discussion
4. Material and Methods
4.1. Chemicals
4.2. Instruments
4.3. Animals
4.4. Preparation of Rat Liver Microsomes
4.5. Cytochrome P450 Probe Substrate Assays
4.5.1. 4-Nitrophenol 2-Hydroxylation Assay for CYP2E1
4.5.2. Testosterone 6β-Hydroxylation Assay for CYP3A4
4.6. Preparation of Standard Solutions
4.7. Optimization of Incubation Conditions in Vitro
4.7.1. Incubation Time
4.7.2. The Concentration of Protein
4.7.3. The Concentration of Probe Substrate
4.8. Effects of Flavonoids in L. clethroides on CYP450 Activity
4.8.1. CYP2E1 Assay
4.8.2. CYP3A4 Assay
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hu, X.M.; Zhang, W.K.; Zhu, Q.S. The Chinese MateriaMedica; Shanghai Science and Technology Press: Shanghai, China, 1999; Volume 16, pp. 96–97. [Google Scholar]
- Yue, S.M.; Chen, B.Q.; Yuan, P.F.; Cui, W.H.; Kang, W.Y. Chemical Constituents of Lysimachia clethroides Duby. Chin. Pharm. J. 2011, 46, 341–343. [Google Scholar]
- Ding, L.F.; Guo, Y.D.; Wu, X.D.; Ma, Y.H. Chemical constituents of flavonoids in Lysimachia clethroide. Chin. Tradit. Pat. Med. 2010, 32, 827–830. [Google Scholar]
- Xu, Q.M.; Tang, L.H.; Li, X.; Hao, L.L.; Li, X.R.; Yang, S.L. Isolation and Identification of 3-O-P-Coumaroyloxyl Pentacyclic Triterpenoids from Lysimachia clethroides Duby. Chin. Pharm. J. 2010, 45, 825–828. [Google Scholar]
- Zou, H.Y.; Tu, P.F. Studies on the chemical constituents of Lysimachia clethroides Duby. Chin. Tradit. Herb. Drugs 2009, 40, 704–708. [Google Scholar]
- Tang, L.H.; Xu, X.Y.; You, B.G.; Zhang, W.; Wang, Y.Q. Anti-tumor effect and its mechanism of total flavones of Lysimachia clethroides Duby. Shanghai J. Tradit. Chin. Med. 2007, 41, 74–76. [Google Scholar]
- Xu, Q.M.; Liu, Y.L.; Feng, Y.L.; Tang, L.H.; Yang, S.L. A new E-ring γ-lactone pentacyclic triterpene from Lysimachia clethroides and its cytotoxic activities. Chem. Nat. Compd. 2012, 48, 597–600. [Google Scholar] [CrossRef]
- Li, C.F.; Song, Y.L.; Liu, Y.X.; Kang, W.Y. Antioxidant activity of extracts from Lysimachia clethroides. Fine Chem. 2008, 25, 1191–1193. [Google Scholar]
- Wei, J.F. Study on Chemical Constitutens, Hepatoprotective and Antihyperglycemic Effect of Lysimachia paridiformis var. stenophylla and Lysimachia clethroides. Ph.D. Thesis, Henan University, Kaifeng, China, 2012. [Google Scholar]
- Liang, D. Study on the Chemical Composition and Biological Activity of Lysimachia clethroides and Chemical Conversion of Synthetic. Ph.D. Thesis, Peking Union Medical College, Beijing, China, 2012. [Google Scholar]
- Cao, L.P.; Jia, R.; Du, J.L.; Ding, W.D.; Yin, G.J. Protective Effect of Glycyrrhiza glabra Extract against tert-butyl hydroperoxide (t-BHP)-induced Hepatotoxicity in Primary Cultured Hepatocytes of Jian Carp (Cyprinuscarpio var. jian). J. Agric. Biotechnol. 2012, 20, 1192–1200. [Google Scholar]
- Pan, Y.; Zhang, L.L. Experimental study of the Malusdoumeri (Bois) Chev flavonoids extracion on chronic hepatic injury induced by CCl4 in rats. LSZ Med. Mater. Med. Res. 2008, 19, 318–319. [Google Scholar]
- Zhang, Q.J.; Yu, H.L.; Li, C.K.; Kang, W.Y. Hepatoprotective effects of Desmodium multiflorum DC on CCl4-induced acute liver injury in mice. Chin. Tradit. Pat. Med. 2011, 33, 1993–1995. [Google Scholar]
- Wei, J.F.; Li, Y.Y.; Yin, Z.H.; Gong, F.; Shang, F.D. Antioxidant activities in vitro and hepatoprotective effects of Lysimachia clethroides Duby on CCl4-induced acute liver injury in mice. Afr. J. Pharm. Pharmacol. 2012, 6, 743–750. [Google Scholar]
- Gonzalez, F.J. Role of cytochromes P450 in chemical toxicity and oxidative stress: Studies with CYP2E1. Mutat. Res. 2005, 569, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Pan, S.H. Influence of the compound Ginkgo Biloba on CYP2E1 and CYP3A4 enzyme activity in hepatic injuried rats. Mil. Med. J. Southeast China. 2011, 13, 100–103. [Google Scholar]
- Huang, H.Y.; Xu, B.; Duan, C.S. Antioxidative activity and components of Lysinachia christinae Hance extracrt. China Oils Fats 2006, 31, 48–51. [Google Scholar]
- Bjornsson, T.D.; Callaghan, J.T.; Einolf, H.J.; Fischer, V.; Gan, L.; Grimm, S.; Kao, J.; King, S.P.; Miwa, G.; Ni, L.; et al. The conduct of in vitro and in vivo drug-drug interaction studies: A pharmaceutical research and manufacturers of America (PhRMA) perspective. Drug Metab. Dispos. 2003, 31, 815–832. [Google Scholar] [CrossRef] [PubMed]
- Thummel, K.E.; Wilkinson, G.R. In vitro and in vivo drug interactions involving human CYP3A. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 389–430. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.F.; Li, T.J.; Zhao, J.Y.; Lin, J.M.; Zhou, J.H.; Hu, J. Effect of Total Akaloids of Rubus alceaefolius poiron on Gene Expressions of CYP2E1 and CYP3A1 in Rats with Acute Liver Injury. Chin. J. Integr. Tradit. Chin. West. Med. 2009, 29, 711–715. [Google Scholar]
- Tian, W.Y.; Wang, Q.X.; Wang, W.J.; Yang, Z. Effect of Gegen Powder on CYP450 Content of Liver Microsomal and Activity of CYP2E1 in Mice Acute Alcohol-induced Liver Injury. Chin. J. Exp. Tradit. Med. Form 2012, 18, 183–186. [Google Scholar]
- Hua, B.C.; Huang, Z.F.; Liu, J.; Cheng, X.L.; Chen, X.F.; Wang, Y.H.; Zhuo, S. Effects of Radix Glycyrrhizae compatibility of Rhizoma dioscorea bulbifera on the activity and expression of CYP450 enzymes in rat liver. Pharmacol. Clin. Chin. Mater. Med. 2014, 30, 79–81. [Google Scholar]
- Gao, Z.W.; Shi, X.J.; Yu, C.; Li, S.J.; Zhong, M.K. Simultaneous determination of the inhibitory potency of compounds on the activity of five cytochrome P-450 enzymes using a cocktail probe substrates method. Yao Xue Xue Bao 2007, 42, 589–594. [Google Scholar] [PubMed]
- Shen, Y.J. Determination of the CYP3A4 and CYP2E1 Enzyme Activity in Rat Liver Microsomes by High Performance Liquid Chromatography and Its Application. Master’s Thesis, Dalian Medical University, Dalian, China, 2013. [Google Scholar]
- Shen, G.L.; Liang, A.H.; Zhao, Y.; Cao, C.Y.; Liu, T.; Li, C.Y.; Odd, G.N. Interaction between four herb compounds and a western drug by CYP3A4 enzyme metabolism in vitro. China J. Chin. Mater. Med. 2009, 34, 1705–1711. [Google Scholar]
- Qin, M.N.; Liu, R.; Liu, G.F.; Dong, F. Effects of Breviscapines Injections on CYP Activities in Rat Liver Microsomes in vitro. China Pharm. 2012, 15, 147–150. [Google Scholar]
- Li, J.K.; He, F.; Bi, H.C.; Zuo, Z.; Liu, B.D.; Luo, H.B.; Huang, M. Inhibition of human cytochrome P-450 CYPP1A2 by flavonoids a quantitative structure activity relationship study. Yao Xue Xue Bao 2008, 43, 1198–1204. [Google Scholar] [PubMed]
- Ho, P.C.; Saville, D.J. Inhibition of human CYP3A4 activity by grapefruit flavonoids, furanocoumarins and related compounds. J. Pharm. Pharm. Sci. 2001, 4, 217–227. [Google Scholar] [PubMed]
- Pan, S.H.; Liu, P.P.; Liu, Y.F.; Gao, Q. Protective effect of compound Ginkgo biloba on alcohol-induced liver injury and its mechanism. Chin. J. Pharmacol. Toxicol. 2012, 26, 41–46. [Google Scholar]
- Lu, Y.K.; Wu, D.F.; Wang, X.D.; Ward, S.C.; Cederbaum, A.I. Chronic alcohol-induced liver injury and oxidant stress are decreased in cytochrome P4502E1 knockout mice and restored in humanized cytochrome P4502E1 knock-in mice. Free Radic. Biol. Med. 2010, 49, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Hamagawa, E.; Tsutsui, C.; Ono, Y.; Ogiri, Y.; Kojo, S. Evaluation Evaluation of oxidative stress during apoptosis and necrosis caused by carbon tetrachloride in rat liver. Biochim. Biophys. Acta 2001, 1535, 186–191. [Google Scholar]
- Leung, T.M.; Nieto, N. CYP2E1 and oxidant stress in alcoholic and non-alcoholic fatty liver disease. J. Hepatol. 2013, 58, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J. Effect and Inhibitory Type of Magnolol and Honokiol on the Activities of CYP450 Isozymes of Rats in Vitro. Master’s Thesis, Hubei University, Wuhan, China, 2012. [Google Scholar]
- Shi, J.; Chen, A.J.; Zhang, F.; Wang, B.J. Effects of Panaxnotoginoside on CYP450 Subtype Enzymes. Chin. J. Pharmacoepidemiol. 2008, 17, 281–284. [Google Scholar]
- Ye, L.H.; Yan, M.Z.; Kong, L.T.; He, M.; Chang, Q. In vitro Inhibition of Quercetin and Its Glycosides on P450 Enzyme Activities. Chin. Pharm. J. 2014, 49, 1051–1055. [Google Scholar]
- Sample Availability: Not available.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | 4-Nitrocatechol | 6β-Hydroxytestosterone | |
|---|---|---|---|
| Regressive equation | Y = 0.0184X − 0.0042 | Y = 0.0880X − 0.0062 | |
| r | 0.9999 | 0.9998 | |
| Linear range | 0.8–51.2 μM | 0.1–25.6 μg·mL−1 | |
| Precision (RSD, %) | High concentration | 0.25 | 0.22 |
| Moderate concentration | 0.35 | 0.46 | |
| Low concentration | 3.93 | 0.96 | |
| Stability (RSD, %) | High concentration | 0.28 | 0.075 |
| Moderate concentration | 0.80 | 0.17 | |
| Low concentration | 2.63 | 1.39 | |
| Metabolite | Original | Added | Found | Recovery (%) | RSD (%) |
|---|---|---|---|---|---|
| 4-Nitrocatechol (μM) | 10.9 | 5 | 15.4 ± 0.11 | 89.4 ± 2.27 | 5.02 |
| 10.9 | 10 | 19.7 ± 0.09 | 87.1 ± 0.85 | ||
| 10.9 | 15 | 25.5 ± 0.17 | 96.8 ± 1.12 | ||
| 6β-Hydroxytestosterone (μg·mL−1) | 0.296 | 0.15 | 0.43 ± 0.01 | 91.5 ± 4.77 | 4.40 |
| 0.296 | 0.30 | 0.55 ± 0.01 | 85.4 ± 1.01 | ||
| 0.296 | 0.45 | 0.69 ± 0.01 | 86.9 ± 2.46 |
| Concentration (μM) | Inhibition Rate (%) | ||||
|---|---|---|---|---|---|
| Clomethiazole | Quercetin | Kaempferol | Isoquercitrin | Astragalin | |
| 4 | 85.41 ± 4.78 | 12.54 ± 0.81 | 16.49 ± 1.23 | 21.12 ± 1.56 | 16.16 ± 1.13 |
| 2 | 68.15 ± 4.97 | 9.99 ± 0.71 | 7.12 ± 0.50 | 22.06 ± 1.12 | 15.58 ± 1.02 |
| 1 | 48.70 ± 2.19 | 7.21 ± 0.43 | 5.50 ± 0.29 | 18.91 ± 1.43 | 13.97 ± 0.81 |
| 0.5 | 25.99 ± 1.27 | 5.46 ± 0.33 | 5.09 ± 0.37 | 16.27 ± 0.83 | 6.14 ± 0.45 |
| 0.25 | 18.49 ± 0.59 | 3.68 ± 0.30 | 0.97 ± 0.80 | 16.02 ± 1.05 | 1.73 ± 0.13 |
| Concentration (μM) | Inhibition Rate (%) | ||||
|---|---|---|---|---|---|
| Ketoconazole | Quercetin | Kaempferol | Isoquercitrin | Astragalin | |
| 5 | 87.28 ± 6.55 | 33.18 ± 1.96 | 26.20 ± 1.81 | 16.68 ± 1.00 | 6.65 ± 0.41 |
| 1 | 76.11 ± 5.25 | 24.97 ± 1.77 | 20.00 ± 1.28 | 13.31 ± 0.63 | 2.57 ± 0.19 |
| 0.2 | 47.33 ± 3.08 | 22.41 ± 1.46 | 17.70 ± 0.80 | 13.04 ± 1.11 | 2.22 ± 0.14 |
| 0.1 | 22.95 ± 1.08 | 15.79 ± 1.23 | 14.12 ± 0.55 | 10.62 ± 0.62 | 0.51 ± 0.04 |
| 0.02 | 14.72 ± 0.56 | 15.04 ± 0.83 | 10.98 ± 0.87 | 9.03 ± 0.35 | 0.42 ± 0.04 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-J.; Xia, Z.-Y.; Wang, J.-M.; Song, X.-T.; Wei, J.-F.; Kang, W.-Y. Effects of Flavonoids in Lysimachia clethroides Duby on the Activities of Cytochrome P450 CYP2E1 and CYP3A4 in Rat Liver Microsomes. Molecules 2016, 21, 738. https://doi.org/10.3390/molecules21060738
Zhang Z-J, Xia Z-Y, Wang J-M, Song X-T, Wei J-F, Kang W-Y. Effects of Flavonoids in Lysimachia clethroides Duby on the Activities of Cytochrome P450 CYP2E1 and CYP3A4 in Rat Liver Microsomes. Molecules. 2016; 21(6):738. https://doi.org/10.3390/molecules21060738
Chicago/Turabian StyleZhang, Zhi-Juan, Zhao-Yang Xia, Jin-Mei Wang, Xue-Ting Song, Jin-Feng Wei, and Wen-Yi Kang. 2016. "Effects of Flavonoids in Lysimachia clethroides Duby on the Activities of Cytochrome P450 CYP2E1 and CYP3A4 in Rat Liver Microsomes" Molecules 21, no. 6: 738. https://doi.org/10.3390/molecules21060738
APA StyleZhang, Z.-J., Xia, Z.-Y., Wang, J.-M., Song, X.-T., Wei, J.-F., & Kang, W.-Y. (2016). Effects of Flavonoids in Lysimachia clethroides Duby on the Activities of Cytochrome P450 CYP2E1 and CYP3A4 in Rat Liver Microsomes. Molecules, 21(6), 738. https://doi.org/10.3390/molecules21060738