
Justicidin B: A Promising Bioactive Lignan †

Abstract
:1. Introduction
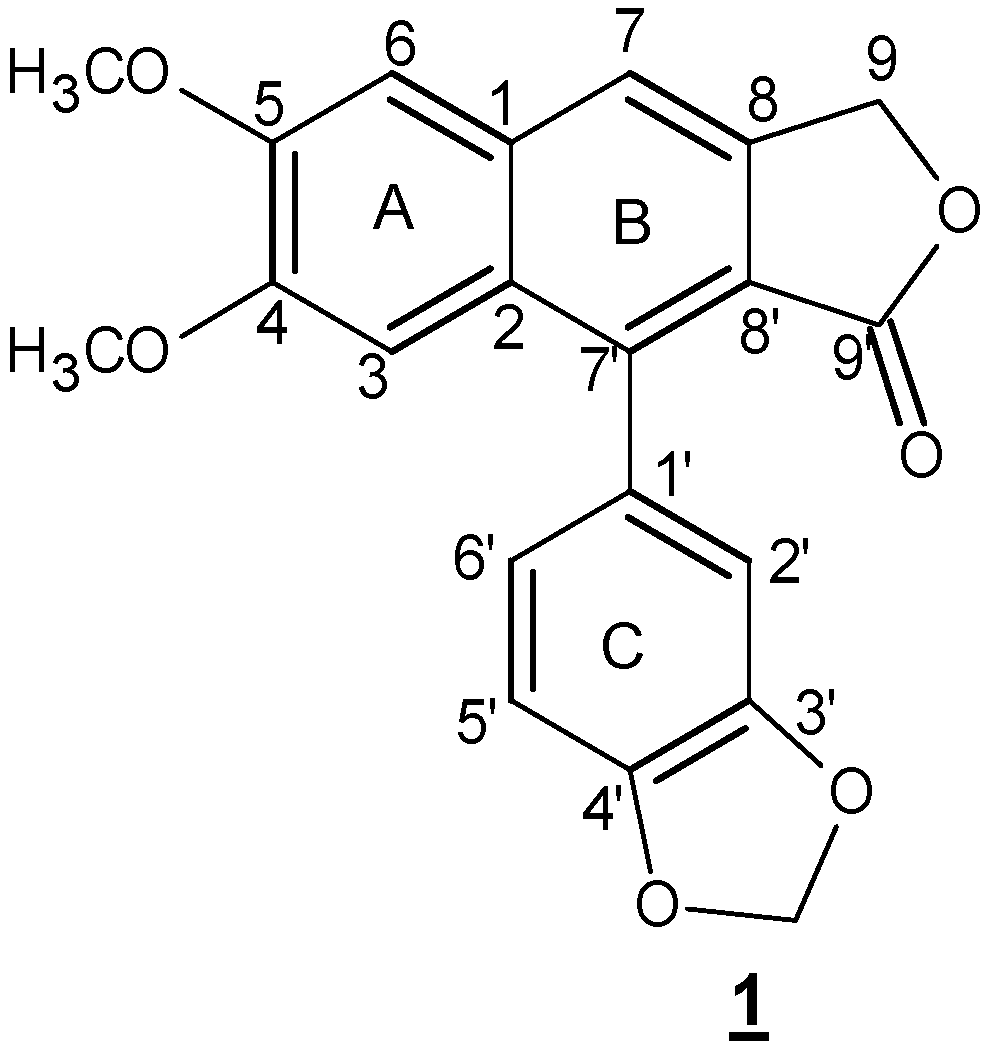
2. Chemistry
Solubility
3. Distribution
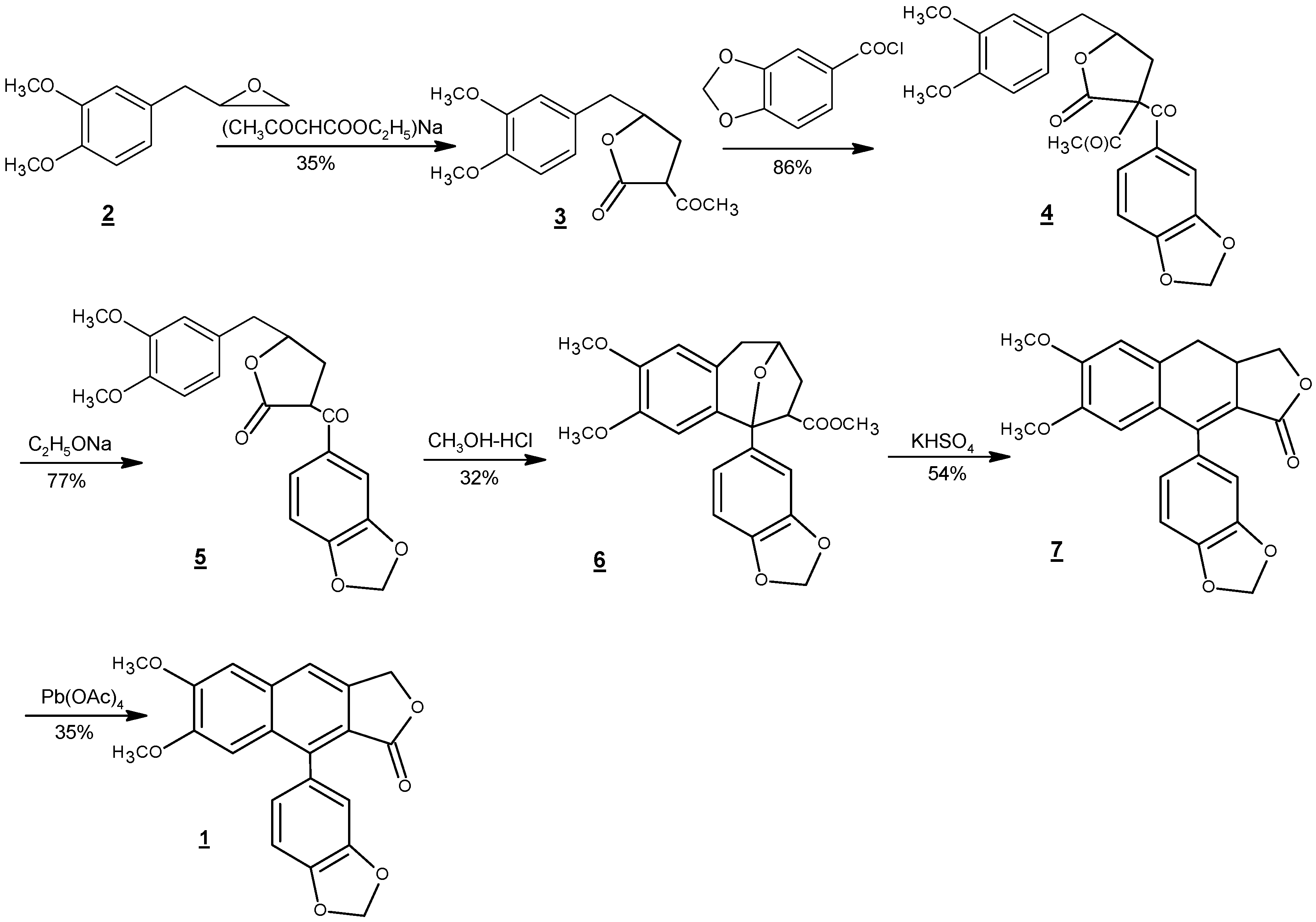
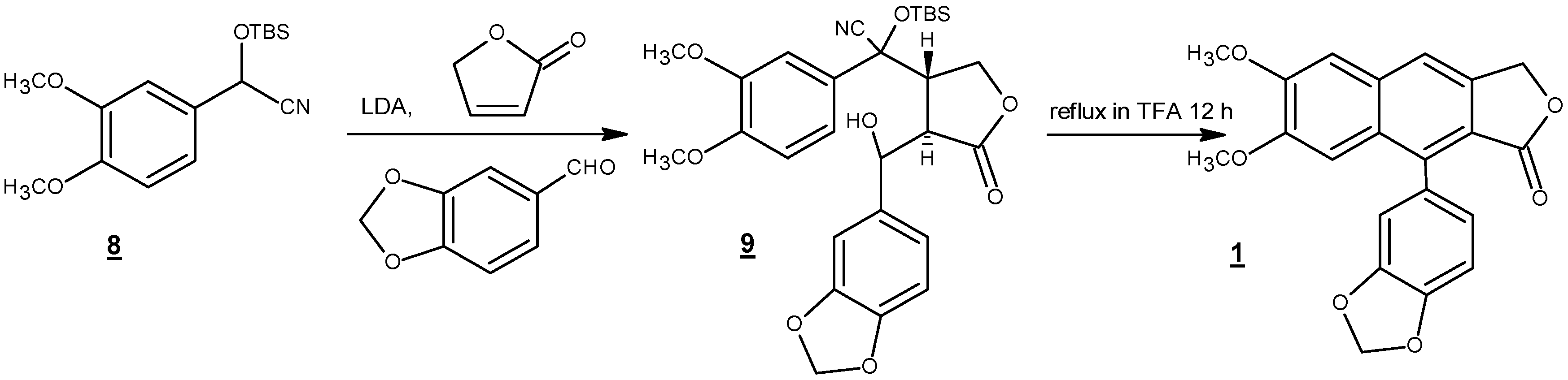
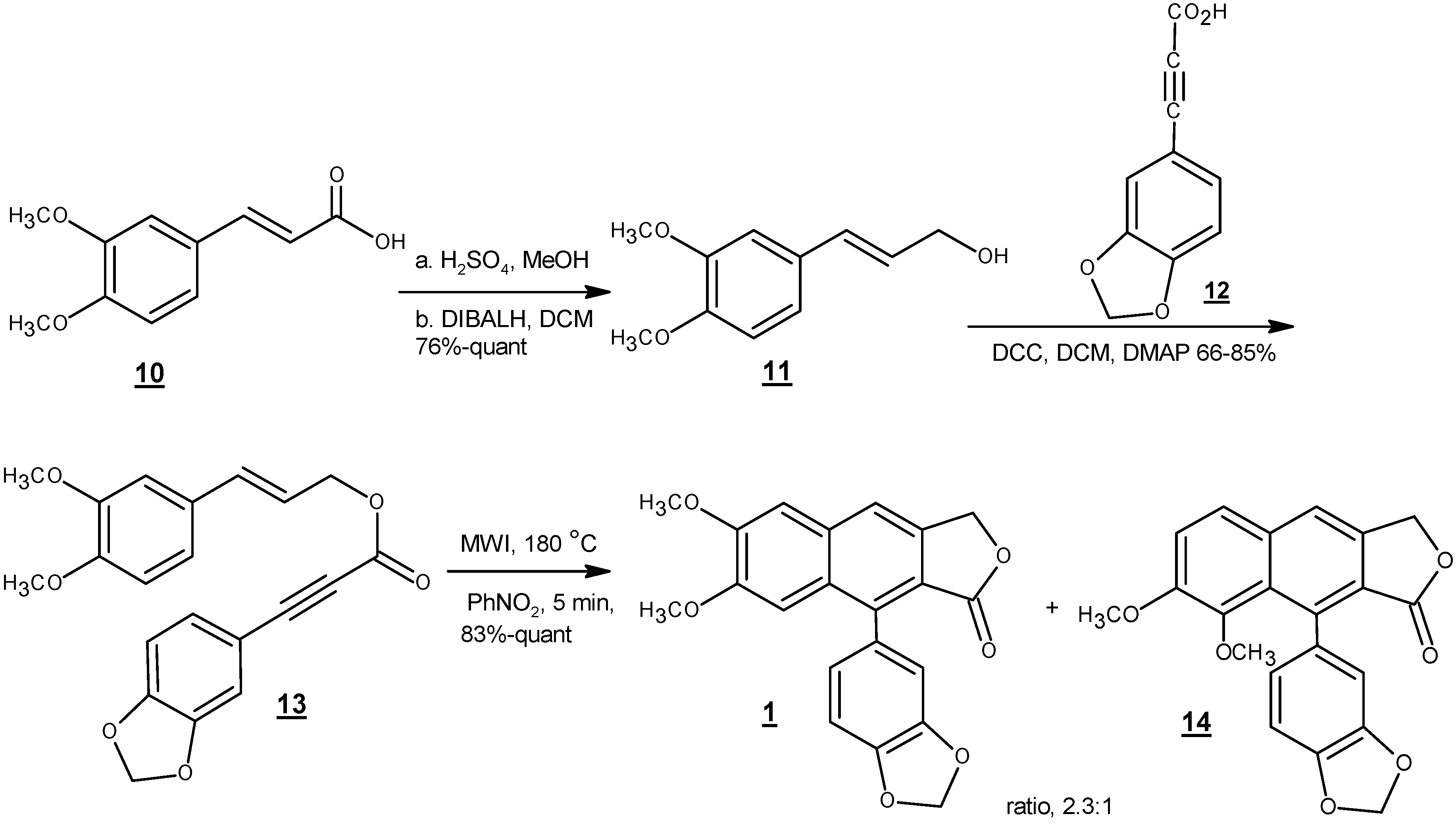
4. Total Synthesis of Justicidin B
5. Production of Justicidin B Using Biotechnology
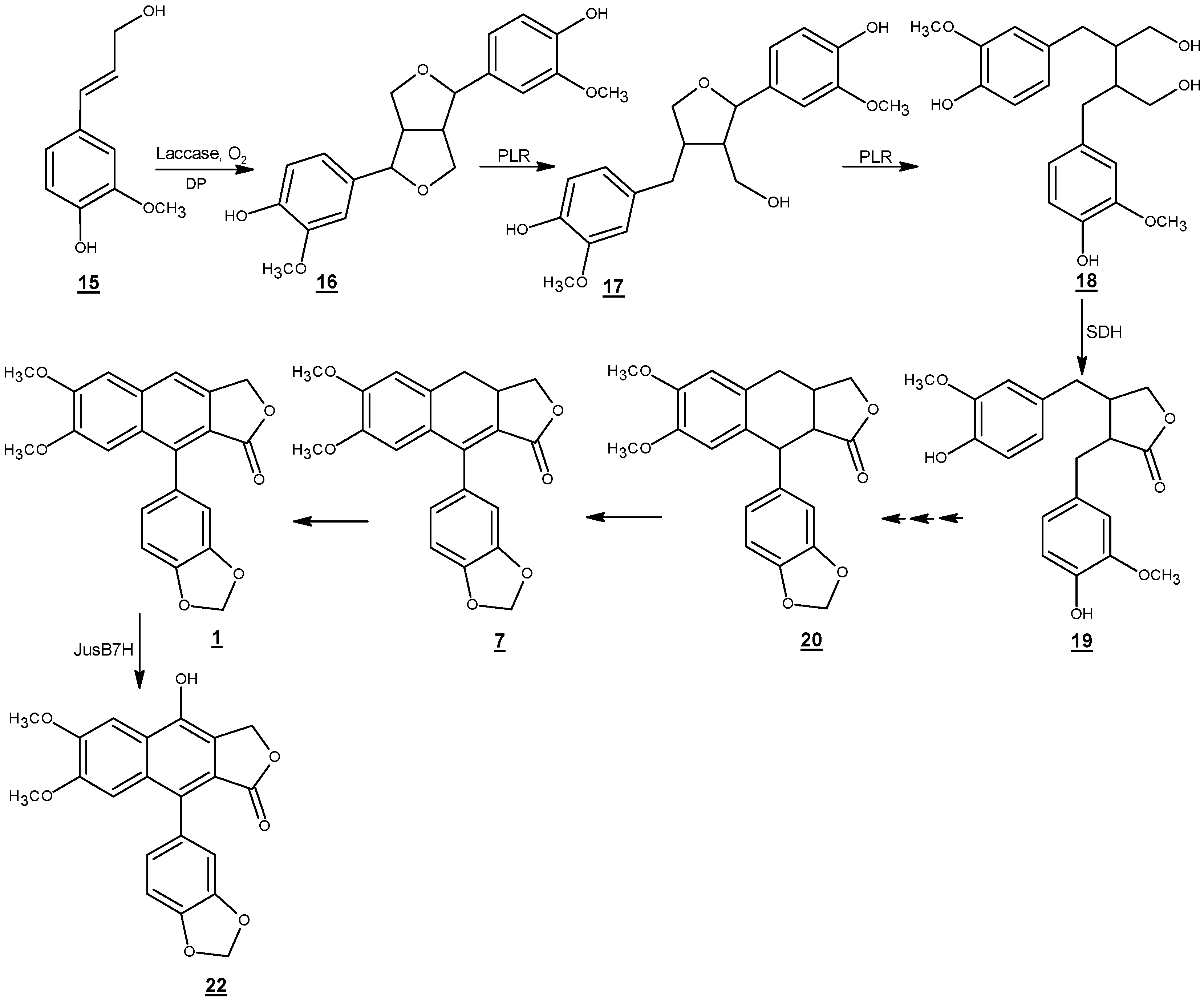
6. Biosynthesis
7. Analysis
7.1. HPLC and HPLC-MS Analysis of Justicidin B
7.2. High Speed Counter Current Chromatography (HSCCC) Analysis
7.3. High Performance Thin Layer Chromatography (HPTLC) Analysis
8. Biological Activities
8.1. Antifungal Activity
8.2. Antiviral Activity
8.3. Antibacterial Activity
8.4. Antiparasitic Activity
8.5. Piscicidal Activity
8.6. Antiplatelet Activity
8.7. Anti-Inflammatory Activity
8.8. Cytotoxic Activity
8.9. Evaluation of in Vivo Animal Toxicity
8.10. Miscellaneous Activities
9. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Davin, L.B.; Lewis, N.G. An historical perspective on lignan biosynthesis: Monolignol, allylphenol and hydroxycinnamic acid coupling and downstream metabolism. Phytochem. Rev. 2003, 2, 257–288. [Google Scholar] [CrossRef]
- Umezawa, T. Diversity in lignan biosynthesis. Phytochem. Rev. 2003, 2, 371–390. [Google Scholar] [CrossRef]
- Lee, K.H.; Xiao, Z. Lignans in treatment of cancer and other diseases. Phytochem. Rev. 2003, 2, 341–362. [Google Scholar] [CrossRef]
- Canel, C.; Moraes, R.M.; Dayan, F.E.; Ferreira, D. Podophyllotoxin. Phytochemistry 2000, 54, 115–120. [Google Scholar] [CrossRef]
- DellaGreca, M.; Zuppolini, S.; Zarrelli, A. Isolation of lignans as seed germination and plant growth inhibitors from mediterranean plants and chemical synthesis of some analogues. Phytochem. Rev. 2013, 12, 717–731. [Google Scholar] [CrossRef]
- Charlton, J.L.; Oleschuk, C.J.; Chee, G.L. Hindered rotation in arylnaphthalene lignans. J. Org. Chem. 1996, 61, 3452–3457. [Google Scholar] [CrossRef]
- Okigawa, M.; Maeda, T.; Kawano, N. The isolation and structure of three new lignans from Justicia procumbens linn. var. Leucantha honda. Tetrahedron 1970, 26, 4301–4305. [Google Scholar] [CrossRef]
- Mohagheghzadeh, A.; Schmidt, T.J.; Alfermann, A.W. Arylnaphthalene lignans from in vitro cultures of Linum austriacum. J. Nat. Prod. 2002, 65, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.; Ruas, M.M.; Donate, P.M. Complete assignments of 1H and 13C NMR spectral data for arylnaphthalene lignana lactones. Magn. Reson. Chem. 2007, 45, 902–904. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J.; Tobler, R.T.; Brun, R.; Sticher, O.; Heilmann, J. Antifungal, antiprotozoal, cytotoxic and piscicidal properties of justicidin B and a new aryinaphthalide lignan from Phyllanthus piscatorum. Planta Med. 2003, 69, 420–424. [Google Scholar] [PubMed]
- Wang, L.; Pan, J.; Yang, M.; Wu, J.; Yang, J. Chromatographic fingerprint analysis and simultaneous determination of eight lignans in Justicia procumbens and its compound preparation by HPLC-DAD. J. Sep. Sci. 2011, 34, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Munakata, K.; Marumo, S.; Ohta, K.; Chen, Y.L. Justicidin A and B, the fish-killing components of Justicia hayatai var. Decumbens. Tetrahedron Lett. 1965, 6, 4167–4170. [Google Scholar] [CrossRef]
- Govindachari, T.R.; Sathe, S.S.; Viswanathan, N.; Pai, B.R.; Srinivasan, M. Revised structures of diphyllin and justicidin A. Tetrahedron Lett. 1967, 36, 3517–3519. [Google Scholar]
- Corrêa, G.M.; de Minas, C. ; Federal University of Minas Gerais. Chemical constituents and biological activities of species of Justicia: A review. Rev. Bras. Farmacogn. 2012, 22, 220–238. [Google Scholar] [CrossRef]
- Asano, J.; Chiba, K.; Tada, M.; Yoshii, T. Antiviral activity of lignans and their glycosides from Justicia procumbens. Phytochemistry 1996, 42, 713–717. [Google Scholar] [CrossRef]
- Joseph, H.; Gleye, J.; Moulis, C.; Mensah, L.J.; Roussakis, C.; Gratas, C. Justicidin B, a cytotoxic principle from Justicia pectoralis. J. Nat. Prod. 1988, 51, 599–600. [Google Scholar] [CrossRef] [PubMed]
- Day, S.H.; Chiu, N.Y.; Won, S.J.; Lin, C.N. Cytotoxic lignans of Justicia ciliata. J. Nat. Prod. 1999, 62, 1056–1058. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, J.; Gopalaiah, K.; Rajasekhar, D.; Subbaraju, G.V. Juspurpurin, an unusual secolignan glycoside from Justicia purpurea. J. Nat. Prod. 2003, 66, 1113–1115. [Google Scholar] [CrossRef] [PubMed]
- Ulubelen, A.; Öztürk, M. Alkaloids, coumarins and lignans from Haplophyllum species. Rec. Nat. Prod. 2008, 2, 54–69. [Google Scholar]
- Batirov, E.K.; Matkarimov, A.D.; Malikov, V.M. Justicidin B and diphyllin from Haplophyllum obtusifolium. Khimija Prir. Soyedineniy 1981, 3, 386–387. [Google Scholar]
- Abdullaev, N.D.; Yagudaev, M.R.; Batirov, E.K.; Malikov, V.M. 13C NMR spectra of arylnaphthalene lignans. Chem. Nat. Compd. 1987, 23, 63–74. [Google Scholar] [CrossRef]
- Batsuren, D.; Batirov, E.K.; Malikov, V.M.; Zemlyanskii, V.N.; Yagudaev, M.R. Arylnaphthalene lignans of Haplophyllum dauricum. The structure of daurinol. Chem. Nat. Compd. 1982, 17, 223–225. [Google Scholar] [CrossRef]
- Ulubelen, A. Alkaloids from Haplophyllum buxbaumii. Phytochemistry 1985, 24, 372–374. [Google Scholar] [CrossRef]
- Nukul, G.S.; Abu Zarga, M.H.; Sabri, S.S.; Al-Eisawi, D.M. Chemical constituents of the flora of Jordan, part III. Mono-o-acetyl diphyllin apioside, a new arylnaphthalene lignan from Haplophyllum buxbaumii. J. Nat. Prod. 1987, 50, 748–750. [Google Scholar] [CrossRef]
- Sheriha, G.M.; Abou Amer, K.M. Lignans of Haplophyllum tuberculatum. Phytochemistry 1984, 23, 151–153. [Google Scholar] [CrossRef]
- Al-Yahya, M.A.; El-Domiaty, M.M.; Al-Meshal, I.A.; Al-Said, M.S.; El-Feraly, F.S. (+)-Dihydroperfamine: An alkaloid from Haplophyllum tuberculatum. Int. J. Pharmacogn. 1991, 29, 268–272. [Google Scholar] [CrossRef]
- Gozler, B.; Arar, G.; Gozler, T.; Hesse, M. Isodaurinol, an arylnaphthalene lignan from Haplophyllum cappadocicum. Phytochemistry 1992, 31, 2473–2475. [Google Scholar]
- Puricelli, L.; Innocenti, G.; Piacente, S.; Caniato, R.; Filippini, R.; Capelletti, E.M. Production of lignans by Haplophyllum patavinum in vivo and in vitro. Heterocycles 2002, 56, 607–612. [Google Scholar]
- Qi, W.; Hua, L.; Gao, K. Chemical constituents of the plants from the genus Phyllanthus. Chem. Biodivers. 2014, 11, 364–395. [Google Scholar] [CrossRef] [PubMed]
- Pettit, G.R.; Cragg, G.M.; Suffness, M.I.; Gust, D.; Boettner, F.E.; Williams, M.; Saenz-Renauld, J.A.; Brown, P.; Schmidt, J.M.; Ellis, P.D. Antineoplastic agents. 104. Isolation and structure of the Phyllanthus acuminatus vahl (Euphorbiaceae) glycosides. J. Org. Chem. 1984, 49, 4258–4266. [Google Scholar] [CrossRef]
- Pettit, G.R.; Schaufelberger, D.E. Isolation and structure of the cytostatic lignan glycoside phyllanthostatin A. J. Nat. Prod. 1988, 51, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, T.L.; Ghia, F.; Torssell, K.B.G. Lignans and lactones from Phyllanthus anisolobus. Phytochemistry 1993, 33, 189–191. [Google Scholar] [CrossRef]
- Lin, M.-T.; Lee, S.-S.; Chen Liu, K.C.S. Phyllamyricins A-C, three novel lignans from Phyllanthus myrtifolius. J. Nat. Prod. 1995, 58, 244–249. [Google Scholar] [CrossRef]
- Wang, C.Y.; Lam, S.H.; Tseng, L.H.; Lee, S.S. Rapid screening of lignans from Phyllanthus myrtifolius and stilbenoids from Syagrus romanzoffiana by HPLC-SPE-NMR. Phytochem. Anal. 2011, 22, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.K.; Fang, S.H.; Tzeng, Y.M. Anti-inflammatory activities of constituents isolated from Phyllanthus polyphyllus. J. Ethnopharmacol. 2006, 103, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Hemmati, S.; Klaes, M.; Konuklugil, B.; Mohagheghzadeh, A.; Ionkova, I.; Fuss, E.; Alfermann, A.W. Lignans in flowering aerial parts of Linum species—Chemodiversity in the light of systematics and phylogeny. Phytochemistry 2010, 71, 1714–1728. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Vossing, S.; Klaes, M.; Grimme, S. An aryldihydronaphthalene lignan with a novel type of ring system and further new lignans cells from Linum perenne L. Planta Med. 2007, 73, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, N.; Ebel, R.; Edrada, R.; Fuss, E.; Alfermann, A.W.; Ionkova, I.; Petrova, A.; Repplinger, M.; Schmidt, T.J. Metabolic profiling of lignan variability in Linum species of section Syllinum native to Bulgaria. Planta Med. 2008, 74, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Klaes, M.; Sendker, J. Lignans in seeds of Linum species. Phytochemistry 2012, 82, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Hui, Y.H.; Chang, C.J.; McLaughlin, J.L.; Powell, R.G. Justicidin B, a bioactive trace lignan from the seeds of Sesbania drummondii. J. Nat. Prod. 1986, 49, 1175–1176. [Google Scholar] [CrossRef] [PubMed]
- El-Gendy, M.M.A.; Hawas, U.W.; Jaspars, M. Novel bioactive metabolites from a marine derived bacterium Nocardia sp. ALAA 2000. J. Antibiot. 2008, 61, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Munakata, K.; Marumo, S.; Ohta, K.; Chen, Y.L. The synthesis of justicidin B and related compounds. Tetrahedron Lett. 1967, 8, 3821–3825. [Google Scholar] [CrossRef]
- Block, E.; Stevenso, R. Lignan lactones. Synthesis of (±)-collinusin and justicidin B. J. Org. Chem. 1971, 36, 3453–3455. [Google Scholar] [CrossRef]
- Momose, T.; Kanai, K.; Hayashi, K. Synthetic studies on lignans and related compounds. VIII. Synthesis of justicidin B and diphyllin and of taiwanin C and E from 2,3-dibenzylidenebutyrolactones via β-apolignans. Chem. Pharm. Bull. 1978, 26, 3195–3198. [Google Scholar] [CrossRef]
- Ghosal, S.; Banerjee, S. Synthesis of retrochinensin; a new naturally occurring 4-aryl-2,3-naphthalide lignan. J. Chem. Soc. Chem. Commun. 1979, 165–166. [Google Scholar] [CrossRef]
- Rodrigo, R. Progress in the chemistry of isobenzofurans: Applications to the synthesis of natural products and polyaromatic hydrocarbons. Tetrahedron 1988, 44, 2093–2135. [Google Scholar] [CrossRef]
- Ogiku, T.; Seki, M.; Takahashi, M.; Ohmizu, H.; Iwasaki, T. A new two-step synthesis of 1-arylnaphthalene lignans from cyanohydrins. Tetrahedron Lett. 1990, 31, 5487–5490. [Google Scholar] [CrossRef]
- Kamal, A.; Daneshtalab, M.; Micetich, R.G. A rapid entry into podophyllotoxin congeners: Synthesis of justicidin B. Tetrahedron Lett. 1994, 35, 3879–3882. [Google Scholar] [CrossRef]
- Kobayashi, K.; Tokimatsu, J.; Maeda, K.; Morikawa, O.; Konishi, H. New, short synthesis of arylnaphthofuranone lignans based on reactions of o-aroylbenzyllithiums with furan-2(5H)-one. J. Chem. Soc. Perkin Trans. 1 1995, 3013–3016. [Google Scholar] [CrossRef]
- Cochran, J.E.; Padwa, A. A new approach to the 1-arylnaphthalene lignans utilizing a tandem Pummerer-Diels-Alder reaction sequence. J. Org. Chem. 1995, 60, 3938–3939. [Google Scholar] [CrossRef]
- Harrowven, D.C.; Bradley, M.; Castro, J.L.; Flanagan, S.R. Total syntheses of justicidin B and retrojusticidin B using a tandem Horner-Emmons-Claisen condensation sequence. Tetrahedron Lett. 2001, 42, 6973–6975. [Google Scholar] [CrossRef]
- Flanagan, S.R.; Harrowven, D.C.; Bradley, M. A new benzannulation reaction and its application in the multiple parallel synthesis of arylnaphthalene lignans. Tetrahedron 2002, 58, 5989–6001. [Google Scholar] [CrossRef]
- Nishii, Y.; Yoshida, T.; Asano, H.; Wakasugi, K.; Morita, J.; Aso, Y.; Yoshida, E.; Motoyoshiya, J.; Aoyama, H.; Tanabe, Y. Regiocontrolled benzannulation of diaryl (gem-dichlorocyclopropyl) methanols for the synthesis of unsymmetrically substituted α-arylnaphthalenes: Application to total synthesis of natural lignan lactones. J. Org. Chem. 2005, 70, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Foley, P.; Eghbali, N.; Anastas, P.T. Silver-catalyzed one-pot synthesis of arylnaphthalene lactone natural products. J. Nat. Prod. 2010, 73, 811–813. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.; Maji, M.; Basak, A. A Garratt-Braverman route to aryl naphthalene lignans. Tetrahedron Lett. 2011, 52, 1183–1186. [Google Scholar] [CrossRef]
- Patel, R.M.; Argade, N.P. Palladium-promoted [2+2+2] cocyclization of arynes and unsymmetrical conjugated dienes: Synthesis of justicidin B and retrojusticidin B. Org. Lett. 2013, 15, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Kudoh, T.; Shishido, A.; Ikeda, K.; Saito, S.; Ishikawa, T. Concise synthesis of arylnaphthalene lignans by regioselective intramolecular anionic Diels-Alder reactions of 1,7-diaryl-1,6-diynes. Synlett 2013, 24, 1509–1512. [Google Scholar] [CrossRef]
- Kocsis, L.S.; Brummond, K.M. Intramolecular dehydro-Diels-Alder reaction affords selective entry to arylnaphthalene or aryldihydronaphthalene lignans. Org. Lett. 2014, 16, 4158–4161. [Google Scholar] [CrossRef] [PubMed]
- Cow, C.; Leung, C.; Charlton, J.L. Antiviral activity of arylnaphthalene and aryldihydronaphthalene lignans. Can. J. Chem. Rev. Can. Chim. 2000, 78, 553–561. [Google Scholar] [CrossRef]
- Wink, M.; Alfermann, A.W.; Franke, R.; Wetterauer, B.; Distl, M.; Windhövel, J.; Krohn, O.; Fuss, E.; Garden, H.; Mohagheghzadeh, A.; et al. Sustainable bioproduction of phytochemicals by plant in vitro cultures: Anticancer agents. Plant Genet. Resour. C 2005, 3, 90–100. [Google Scholar] [CrossRef]
- Ionkova, I. Podophyllotoxin and related lignans: Biotechnological production by in vitro plant cell cultures. In Medicinal Plant Biotechnology, 1st ed.; Arora, R., Ed.; CABI: Wallingford, CT, USA, 2010; pp. 138–155. [Google Scholar]
- Fuss, E. Lignans in plant cell and organ cultures: An overview. Phytochem. Rev. 2003, 2, 307–320. [Google Scholar] [CrossRef]
- Alfermann, A.; Petersen, M.; Fuss, E. Production of natural products by plant cell biotechnology: Results, problems and perspectives. In Plant Tissue Culture 100 Years since Gottlieb Haberlandt, 1st ed.; Laimer, M., Rucker, W., Eds.; Springer-Verlag Wien: Vienna, Austria, 2003; pp. 153–166. [Google Scholar]
- Innocenti, G.; Puricelli, L.; Piacente, S.; Caniato, R.; Filippini, R.; Cappelletti, E.M. Patavine, a new arylnaphthalene lignan glycoside from shoot cultures of Haplophyllum patavinum. Chem. Pharm. Bull. 2002, 50, 844–846. [Google Scholar] [CrossRef] [PubMed]
- Konuklugil, B.; Ionkova, I.; Vasilev, N.; Schmidt, T.J.; Windhövel, J.; Fuss, E.; Alfermann, A.W. Lignans form Linum species of sections Syllinum and Linum. Nat. Prod. Res. 2007, 21, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nikolov, S.; Momekov, G.; Kitanov, G.; Ionkova, I.; Krasteva, I.; Toshkova, R.; Konstantinov, S.; Nedialkov, P.; Karaivanova, M. Exploitation of the bulgarian flora's biodiversity as a source of immunomodulatory and/or antineoplastic agents: Current challenges and perspectives. Biotechnol. Biotechnol. Equip. 2007, 21, 471–477. [Google Scholar] [CrossRef]
- Vasilev, N.P.; Ionkova, I. Cytotoxic activity of extracts from Linum cell cultures. Fitoterapia 2005, 76, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Ionkova, I.; Sasheva, P.; Ionkov, T.; Momekov, G. Linum narbonense: A new valuable tool for biotechnological production of a potent anticancer lignan justicidine B. Pharmacogn. Mag. 2013, 9, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, N.; Elfahmi; Bos, R.; Kayser, O.; Momekov, G.; Konstantinov, S.; Ionkova, I. Production of justicidin B, a cytotoxic arylnaphthalene lignan from genetically transformed root cultures of Linum leonii. J. Nat. Prod. 2006, 69, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Ionkova, I.; Sasheva, P.; Momekov, G. Justicidin B—A potent cytotoxic arylnaphtalene lignan from in vitro cultures of Linum leonii. Planta Med. 2011, 77, 1384. [Google Scholar] [CrossRef]
- Vasilev, N.; Ionkova, I. Lignan production by cell cultures of Linum setaceum and Linum campanulatum. Pharm. Biol. 2005, 43, 509–511. [Google Scholar] [CrossRef]
- Mohagheghzadeh, A.; Dehshahri, S.; Hemmati, S. Accumulation of lignans by in vitro cultures of three Linum species. Z. Naturforsch. C 2009, 64, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.; Schneider, B.; Schmidt, T.; Federolf, K.; Alfermann, A.W.; Fuss, E. Biosynthesis of justicidin B and diphyllin in cell cultures of Linum perenne L. Himmelszelt. In Proceedings of the 7th Joint Meeting of GA, AFERP, ASP, ESP & SIF, Athens, Greece, 3–8 Auguest 2008; Skaltsounis, L., Magiatis, P., Eds.; Volume 74, p. 1168.
- Hemmati, S.; Schmidt, T.J.; Fuss, E. (+)-Pinoresinol/(−)-lariciresinol reductase from Linum perenne Himmelszelt involved in the biosynthesis of justicidin B. FEBS Lett. 2007, 581, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Jullian-Pawlicki, N.; Lequart-Pillon, M.; Huynh-Cong, L.; Lesur, D.; Cailleu, D.; Mesnard, F.; Laberche, J.C.; Gontier, E.; Boitel-Conti, M. Arylnaphthalene and aryltetralin-type lignans in hairy root cultures of Linum perenne, and the stereochemistry of 6-methoxypodophyllotoxin and one diastereoisomer by HPLC-MS and NMR spectroscopy. Phytochem. Anal. 2015, 26, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Satake, H.; Koyama, T.; Bahabadi, S.E.; Matsumoto, E.; Ono, E.; Murata, J. Essences in metabolic engineering of lignan biosynthesis. Metabolites 2015, 5, 270–290. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.; Schneider, B.; Schmidt, T.J.; Federolf, K.; Alfermann, A.W.; Fuss, E. Justicidin B 7-hydroxylase, a cytochrome p450 monooxygenase from cell cultures of Linum perenne Himmelszelt involved in the biosynthesis of diphyllin. Phytochemistry 2007, 68, 2736–2743. [Google Scholar] [CrossRef] [PubMed]
- Davin, L.B.; Bedgar, D.L.; Katayama, T.; Lewis, N.G. On the stereoselective synthesis of (+)-pinoresinol in Forsythia suspensa from its achiral precursor, coniferyl alcohol. Phytochemistry 1992, 31, 3869–3874. [Google Scholar] [CrossRef]
- Halls, S.C.; Davin, L.B.; Kramer, D.M.; Lewis, N.G. Kinetic study of coniferyl alcohol radical binding to the (+)-pinoresinol forming dirigent protein. Biochemistry 2004, 43, 2587–2595. [Google Scholar] [CrossRef] [PubMed]
- Pickel, B.; Constantin, M.A.; Pfannstiel, J.; Conrad, J.; Beifuss, U.; Schaller, A. N enantiocomplementary dirigent protein for the enantioselective lacease-catalyzed oxidative coupling of phenols. Angew. Chem. Int. Ed. 2010, 49, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Davin, L.B.; Wang, H.B.; Crowell, A.L.; Bedgar, D.L.; Martin, D.M.; Sarkanen, S.; Lewis, N.G. Stereoselective bimolecular phenoxy radical coupling by an auxiliary (dirigent) protein without an active center. Science 1997, 275, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Kazenwadel, C.; Klebensberger, J.; Richter, S.; Pfannstiel, J.; Gerken, U.; Pickel, B.; Schaller, A.; Hauer, B. Optimized expression of the dirigent protein AtDIR6 in Pichia pastoris and impact of glycosylation on protein structure and function. Appl. Microbiol. Biotechnol. 2013, 97, 7215–7227. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Moinuddin, S.G.A.; Atwell, K.M.; Costa, M.A.; Davin, L.B.; Lewis, N.G. Opposite stereoselectivities of dirigent proteins in arabidopsis and schizandra species. J. Biol. Chem. 2012, 287, 33957–33972. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.; von Heimendahl, C.B.; Klaes, M.; Alfermann, A.W.; Schmidt, T.J.; Fuss, E. Pinoresinol-lariciresinol reductases with opposite enantiospecificity determine the enantiomeric composition of lignans in the different organs of Linum usitatissimum L. Planta Med. 2010, 76, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Gang, D.R.; Davin, L.B.; Lewis, N.G. Recombinant pinoresinol-lariciresinol reductases from western red cedar (Thuja plicata) catalyze opposite enantiospecific conversions. J. Biol. Chem. 1999, 274, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.Q.; Costa, M.A.; Pelissier, H.C.; Davin, L.B.; Lewis, N.G. Secoisolariciresinol dehydrogenase purification, cloning, and functional expression. Implications for human health protection. J. Biol. Chem. 2001, 276, 12614–12623. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.L. HPLC in natural product analysis: The detection issue. Planta Med. 2009, 75, 719–734. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.T.; Wang, X.G.; Xu, R.; Meng, F.H.; Yu, N.J.; Zhao, Y.M. Qualitative and quantitative analysis of lignan constituents in Caulis trachelospermi by HPLC-QTOF-MS and HPLC-UV. Molecules 2015, 20, 8107–8124. [Google Scholar] [CrossRef] [PubMed]
- Eklund, P.C.; Backman, M.J.; Kronberg, L.Å.; Smeds, A.I.; Sjöholm, R.E. Identification of lignans by liquid chromatography-electrospray ionization ion-trap mass spectrometry. J. Mass Spectrom. 2008, 43, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Hemmati, S.; Fuss, E.; Alfermann, A.W. A combined HPLC-UV and HPLC-MS method for the identification of lignans and its application to the lignans of Linum usitatissimum L. and L. bienne Mill. Phytochem. Anal. 2006, 17, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Luo, Q.; Ding, L.; Fang, F.; Yuan, Y.; Chen, J.; Zhang, J.; Jin, H.; He, S. Preparative isolation and purification of lignans from Justicia procumbens using high-speed counter-current chromatography in stepwise elution mode. Molecules 2015, 20, 7048–7058. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, N.; Nedialkov, P.; Ionkova, I.; Ninov, S. HPTLC densitomeric determination of justicidin B in Linum in vitro cultures. Pharmazie 2004, 59, 528–529. [Google Scholar] [PubMed]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.; Lee, J.; Liu, Q.H.; Woo, E.R.; Lee, D.G. Antifungal effect of (+)-pinoresinol isolated from Sambucus williamsii. Molecules 2010, 15, 3507–3516. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.; Cho, J.; Hwang, I.S.; Jin, H.G.; Woo, E.R.; Lee, D.G. Antifungal activity of lariciresinol derived from Sambucus williamsii and their membrane-active mechanisms in Candida albicans. Biochem. Biophys. Res. Commun. 2011, 410, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Välimaa, A.L.; Honkalampi-Hämäläinen, U.; Pietarinen, S.; Willför, S.; Holmbom, B.; von Wright, A. Antimicrobial and cytotoxic knotwood extracts and related pure compounds and their effects on food-associated microorganisms. Int. J. Food Microbiol. 2007, 115, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Panagouleas, C.; Skaltsa, H.; Lazari, D.; Skaltsounis, A.L.; Sokovic, M. Antifungal activity of secondary metabolites of Centaurea raphanina ssp. mixta, growing wild in greece. Pharm. Biol. 2003, 41, 266–270. [Google Scholar]
- Windayani, N.; Rukayadi, Y.; Hakim, E.H.; Ruslan, K.; Syah, Y.M. Antifungal activity of lignans isolated from Phyllanthus myrtifolius Moon. against Fusarium oxysporum. Phytochemistry 2014, 12, 33–39. [Google Scholar]
- Chang, C.W.; Lin, M.T.; Lee, S.S.; Liu, K.C.; Hsu, F.L.; Lin, J.Y. Differential inhibition of reverse transcriptase and cellular DNA polymerase-alpha activities by lignans isolated from chinese herbs, Phyllanthus myrtifolius moon, and tannins from Lonicera japonica thunb and Castanopsis hystrix. Antivir. Res. 1995, 27, 367–374. [Google Scholar] [PubMed]
- MacRae, W.D.; Hudson, J.B.; Towers, G.H.N. The antiviral action of lignans. Planta Med. 1989, 55, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Charlton, J.L. Antiviral activity of lignans. J. Nat. Prod. 1998, 61, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Kajunguri, D.; Hargrove, J.W.; Ouifki, R.; Mugisha, J.; Coleman, P.G.; Welburn, S.C. Modelling the use of insecticide-treated cattle to control tsetse and Trypanosoma brucei rhodesiense in a multi-host population. Bull. Math. Biol. 2014, 76, 673–696. [Google Scholar] [PubMed]
- Baker, N.; de Koning, H.P.; Mäser, P.; Horn, D. Drug resistance in african trypanosomiasis: The melarsoprol and pentamidine story. Trends Parasitol. 2013, 29, 110–118. [Google Scholar] [PubMed]
- Kennedy, P.G. Clinical features, diagnosis, and treatment of human african trypanosomiasis (sleeping sickness). Lancet Neurol. 2013, 12, 186–194. [Google Scholar] [CrossRef]
- Urbina, J.A. Recent clinical trials for the etiological treatment of chronic chagas disease: Advances, challenges and perspectives. J. Eukaryot. Microbiol. 2015, 62, 149–156. [Google Scholar] [CrossRef] [PubMed]
- De Souza, V.A.; da Silva, R.; Pereira, A.C.; de A. Royo, V.; Saraiva, J.; Montanheiro, M.; de Souza, G.H.; da Silva Filho, A.A.; Grando, M.D.; Donate, P.M.; et al. Trypanocidal activity of (−)-cubebin derivatives against free amastigote forms of Trypanosoma cruzi. Bioorg. Med. Chem. Lett. 2005, 15, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.J.; Khalid, S.A.; Romanha, A.J.; Alves, T.M.A.; Biavatti, M.W.; Brun, R.; Da Costa, F.B.; de Castro, S.L.; Ferreira, V.F.; de Lacerda, M.V.G.; et al. The potential of secondary metabolites from plants as drugs or leads against protozoan neglected diseases-part II. Curr. Med. Chem. 2012, 19, 2176–2228. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Chen, Y.L.; Marumo, S.; Munakata, K. Studies on piscicidal components of Justicia hayatai var decumbens. Part I. Isolation and piscicidal activities of justicidin A and B. Agric. Biol. Chem. 1969, 33, 610–614. [Google Scholar]
- Inamori, Y.; Kubo, M.; Tsujibo, H.; Ogawa, M.; Baba, K.; Kozawa, M.; Fujita, E. The biological activities of podophyllotoxin compounds. Chem. Pharm. Bull. 1986, 34, 3928–3932. [Google Scholar] [CrossRef] [PubMed]
- Lievens, D.; von Hundelshausen, P. Platelets in atherosclerosis. Thromb. Haemost. 2011, 106, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Schmaier, A.H. Platelet aggregation testing in platelet-rich plasma: Description of procedures with the aim to develop standards in the field. Am. J. Clin. Pathol. 2005, 123, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Siess, W. Molecular mechanisms of platelet activation. Physiol. Rev. 1989, 69, 58–178. [Google Scholar] [PubMed]
- Chen, C.C.; Hsin, W.C.; Ko, F.N.; Huang, Y.L.; Ou, J.C.; Teng, C.M. Antiplatelet arylnaphthalide lignans from Justicia procumbens. J. Nat. Prod. 1996, 59, 1149–1150. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.M.; Wu, S.C.; Chung, W.J.; Lin, H.C.; Chen, K.T.; Chen, Y.C.; Hsu, M.F.; Yang, J.M.; Wang, J.P.; Lin, C.N. Antiplatelet effect and selective binding to cyclooxygenase (COX) by molecular docking analysis of flavonoids and lignans. Int. J. Mol. Sci. 2007, 8, 830–841. [Google Scholar] [CrossRef]
- Hou, R.C.W.; Chen, H.L.; Tzen, J.T.; Jeng, K.C.G. Effect of sesame antioxidants on LPS-induced NO production by BV2 microglial cells. Neuroreport 2003, 14, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Seo, K.H.; Jeong, R.H.; Lee, S.M.; Kim, G.S.; Noh, H.J.; Kim, S.Y.; Kim, G.W.; Kim, J.Y.; Baek, N.I. Anti-inflammatory lignans from the fruits of Acanthopanax sessiliflorus. Molecules 2012, 18, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.K.; Geethangili, M.; Fang, S.H.; Tzeng, Y.M. Antioxidant and cytotoxic activities of naturally occurring phenolic and related compounds: A comparative study. Food Chem. Toxicol. 2007, 45, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- McGahon, A.; Bissonnette, R.; Schmitt, M.; Cotter, K.M.; Green, D.R.; Cotter, T.G. BCR-ABL maintains resistance of chronic myelogenous leukemia cells to apoptotic cell death. Blood 1994, 83, 1179–1187. [Google Scholar] [PubMed]
- Luo, J.; Hu, Y.; Kong, W.; Yang, M. Evaluation and structure-activity relationship analysis of a new series of arylnaphthalene lignans as potential anti-tumor agents. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Momekov, G.; Konstantinov, S.; Dineva, I.; Ionkova, I. Effect of justicidin B—A potent cytotoxic and pro-apoptotic arylnaphtalene lignan on human breast cancer-derived cell lines. Neoplasma 2011, 58, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Momekov, G.; Yossifov, D.; Guenova, M.; Michova, A.; Stoyanov, N.; Konstantinov, S.; Ionkov, T.; Sacheva, P.; Ionkova, I. Apoptotic mechanisms of the biotechnologically produced arylnaphtalene lignan justicidin B in the acute myeloid leukemia-derived cell line HL-60. Pharmacol. Rep. 2014, 66, 1073–1076. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Yin, H.L.; Liu, S.J.; Chen, L.; Tian, Y.; Li, B.; Wang, Q.; Dong, J.X. Cytotoxic activity of lignans from Justicia procumbens. Fitoterapia 2014, 94, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Ilieva, Y.; Zhelezova, I.; Atanasova, T.; Zaharieva, M.M.; Sasheva, P.; Ionkova, I.; Konstantinov, S. Cytotoxic effect of the biotechnologically-derived justicidin B on human lymphoma cells. Biotechnol. Lett. 2014, 36, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Siggers, T.; Chang, A.B.; Teixeira, A.; Wong, D.; Williams, K.J.; Ahmed, B.; Ragoussis, J.; Udalova, I.A.; Smale, S.T.; Bulyk, M.L. Principles of dimer-specific gene regulation revealed by a comprehensive characterization of NF-κB family DNA binding. Nat. Immunol. 2012, 13, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Baba, A.; Kawamura, N.; Makino, H.; Ohta, Y.; Taketomi, S.; Sohda, T. Studies on disease-modifying antirheumatic drugs: Synthesis of novel quinoline and quinazoline derivatives and their anti-inflamatory effect. J. Med. Chem. 1996, 39, 5176–5182. [Google Scholar] [CrossRef] [PubMed]
- Ueng, Y.F.; Chen, C.C.; Chen, C.F. Inhibition of benzo(a)pyrene hydroxylation by lignans isolated from Justicia procumbens. J. Food Drug Anal. 2000, 8, 309–314. [Google Scholar]
- Nebert, D.W.; Shi, Z.; Galvez-Peralta, M.; Uno, S.; Dragin, N. Oral benzo[a]pyrene: Understanding pharmacokinetics, detoxication, and consequences-Cyp1 knockout mouse lines as a paradigm. Mol. Pharmacol. 2013, 84, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Shertzer, H.G.; Puga, A.; Chang, C.; Smith, P.; Nebert, D.W.; Setchell, K.D.; Dalton, T.P. Inhibition of CYP1A1 enzyme activity in mouse hepatoma cell culture by soybean isoflavones. Chem.-Biol. Interact. 1999, 123, 31–49. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | δC (ppm) | H | δH (ppm) | Multiplicity | Coupling Constant (Hz) | gHMBC | gCOSY | gHSQC |
|---|---|---|---|---|---|---|---|---|
| 1′ | 128.3 | - | - | - | - | - | - | - |
| 2′ | 110.5 | 2′ | 6.78 | d | J (2′, 6′) = 1.5 | C-3′, 6′, 1′, 8′ | H-5′, 6′, 12 | H-2′ |
| 3′ | 148.0 | - | - | - | - | - | - | - |
| 4′ | 147.8 | - | - | - | - | - | - | - |
| 5′ | 105.8 | 5′ | 6.98 | d | J (5′, 6′) = 7.8 | C-6′, 1′, 4′ | H-2′, 6′ | H-5′ |
| 6′ | 123.4 | 6′ | 6.77 | d | J (6′, 5′) = 7.8 J (6′, 2′) = 1.5 | C-4′, 8′, 2′, 5′ | H-5′, 2′ | H-6′ |
| 7′ | 139.5 | - | - | - | - | - | - | - |
| 8′ | 118.4 | - | - | - | - | - | - | - |
| 9′ | 169.9 | - | - | - | - | - | - | - |
| 10 | 101.2 | 10a 10b | 6.05 | s | - | C-4′, 3′ | H-2′ | H-10a H-10b |
| 1 | 133.1 | - | - | - | - | |||
| 2 | 128.8 | - | - | - | - | - | - | - |
| 3 | 108.1 | 5 | 7.08 | s | - | C-7, 2, 1 | H-6, 7, 11 | H-3 |
| 4 | 151.7 | - | - | - | - | - | - | - |
| 5 | 150.0 | - | - | - | - | - | - | - |
| 6 | 106.0 | 2 | 7.12 | s | - | C-5, 1, 2, 7 | H-7, 10 | H-6 |
| 7 | 118.2 | 7 | 7.61 | s | - | C-2, 8′, 6 | H-9, 6, 3 | H-7 |
| 8 | 139.4 | - | - | - | - | - | - | - |
| 9 | 67.9 | 9a 9b | 5.33 | s | - | C-8′, 2, 1, 8, 9′ | H-7 | H-9a H-9b |
| 11 | 56.0 | 11 | 3.70 | s | - | C-5 | H-7 | H-11 |
| 12 | 55.8 | 12 | 3.95 | s | - | C-4 | H-3 | H-12 |
| Molecular Mass | 364 | |||
|---|---|---|---|---|
| Cluster ions | [2M + Na]+ | [2M + NH4]+ | [2M + H]+ | |
| 751 c | 746 a | - | ||
| Quasi molecular ions | [M + Na]+ | [M + NH4]+ | [M + H]+ | |
| 387 a | - | 365 d | ||
| Fragment ions | [M + H − H2O]+ | [M + H − CH2O]+ | [M + H − CO2]+ | |
| - | 335 c | 321 c | 303 b 291 b 289 b | |
| Column | A (Solvent) | B (Solvent) | Time (min) | Flow Rate (mL/min) | A% | B% | Rt * (min) | Ref. |
|---|---|---|---|---|---|---|---|---|
| YMC pack ODS-A (150 mm × 4.6 mm × 5 µm) (25 °C) | water | methanol | 0 | 0.8 | 90 | 10 | 49 | [91] |
| 60 | 10 | 90 | ||||||
| 70 | 10 | 90 | ||||||
| Nucleosil 100-C18 (40 + 250 mm × 4.6 mm × 5 µm) | water | acetonitrile | 0 | 0.8 | 55 | 45 | 15.51 | [8] ** |
| 17 | 33 | 67 | ||||||
| 18 | 50 | 50 | ||||||
| 25 | 55 | 45 | ||||||
| Nucleodur 100-5 C18 (125 mm × 4.6 mm × 5 µm) | water | methanol | 0 | 1.4 | 70 | 30 | 26.5 | [90] |
| 30 | 30 | 70 | ||||||
| 45 | 0 | 100 | ||||||
| 55 | 0 | 100 | ||||||
| GROM-Sil C18 (20 + 250 mm × 4 mm × 5 µm) | (0.1% H3PO4) in water | acetonitrile | 0 | 0.8 | 75 | 25 | - | [74] |
| 25 | 0.8 | 62 | 38 | |||||
| 43 | 1 | 57 | 43 | |||||
| 46 | 1 | 45 | 55 | |||||
| 54 | 1 | 30 | 70 | |||||
| 56 | 1 | 75 | 25 | |||||
| 60 | 0.8 | 75 | 25 | |||||
| Shiseido Capcell Pak C18 (250 mm × 4.6 mm × 5 µm) (35 °C) | water | acetonitrile | 0 | 0.8 | 80 | 20 | 54 | [11] |
| 40 | 54 | 46 | ||||||
| 60 | 45 | 55 | ||||||
| 70 | 40 | 60 | ||||||
| 75 | 60 | 40 |
| Parameters | Conditions | ||
|---|---|---|---|
| HPLC-ESI/MS | |||
| Column: Knauer Eurosphere RPC18 (250 mm × 2 mm × 5 µm) | |||
| Time (min) | A% (Water + 0.1% Formic Acid) | B% (Acetonitrile) | |
| Gradient | 0 | 70 | 30 |
| 30 | 30 | 70 | |
| 40 | 30 | 70 | |
| 55 | 0 | 100 | |
| 65 | 0 | 100 | |
| Flow rate | 0.4 mL/min | ||
| Ionization mode | ESI + and − mode | ||
| Capillary temp | 300 °C | ||
| Source voltage | 5 kV | ||
| Parameters | Conditions | ||
|---|---|---|---|
| HPLC-ESI/MSn | |||
| Column: Shiseido Capcell Pak C18 (250 mm × 4.6 mm × 5 µm) (35 °C) | |||
| Time (min) | A% (Water) | B% (Acetonitrile) | |
| Gradient | 0 | 80 | 20 |
| 40 | 54 | 46 | |
| 60 | 45 | 55 | |
| 70 | 60 | 40 | |
| 75 | 60 | 40 | |
| Flow rate | 0.8 mL/min | ||
| Ionization mode | ESI + mode | ||
| Capillary temp | 330 °C | ||
| Source voltage | 4.5 kV | ||
| Capillary voltage | 18 V | ||
| Sheath gas | (N2) flow, 50 AU | ||
| Auxillary gas | (N2) flow, 15 AU | ||
| Cell Line | Justicidin B | Diphyllin | Etoposide | Helanin |
|---|---|---|---|---|
| NSCLCN6 | 28 * | - | - | - |
| KB HeLa | 0.2 * | - | - | 0.2 * |
| Jurkat T | 3.2 * | - | - | 0.03 * |
| L6 | 3.3 * | - | - | - |
| PBM Cs | 4.7 * | - | - | 0.03 * |
| LAMA-84 | 1.11 | - | 0.79 | - |
| K-562 | 6.08 | - | 1.87 | - |
| SKW-3 | 1.62 | - | 0.82 | - |
| MDA-MB-231 | 106.9 | - | 63.1 | - |
| MCF-7 | 38.7 | - | 52.7 | - |
| HL-60 | 0.9 | - | - | - |
| L1210 | 6.3 | - | 14.2 | - |
| P388D1 | 7.3 | - | - | - |
| L0V0 | 6.081 | 8.120 | - | - |
| BGC-823 | 0.179 | 8.079 | - | - |
| Compound | HD-MY-Z | DOHH-2 | REH | HH | HuT-78 | OPM-2 | RPMI-8226 | U-266 |
|---|---|---|---|---|---|---|---|---|
| justicidin B | 144.5 | 5 | 8 | 16.2 | 6.1 | 1.5 | 0.17 | 17.2 |
| etoposide | >100 | 9.5 | 0.015 | 14.7 | 4.2 | 1.3 | 14.9 | 27.4 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemmati, S.; Seradj, H. Justicidin B: A Promising Bioactive Lignan. Molecules 2016, 21, 820. https://doi.org/10.3390/molecules21070820
Hemmati S, Seradj H. Justicidin B: A Promising Bioactive Lignan. Molecules. 2016; 21(7):820. https://doi.org/10.3390/molecules21070820
Chicago/Turabian StyleHemmati, Shiva, and Hassan Seradj. 2016. "Justicidin B: A Promising Bioactive Lignan" Molecules 21, no. 7: 820. https://doi.org/10.3390/molecules21070820
APA StyleHemmati, S., & Seradj, H. (2016). Justicidin B: A Promising Bioactive Lignan. Molecules, 21(7), 820. https://doi.org/10.3390/molecules21070820