Effect of Abscisic Acid on Accumulation of Five Active Components in Root of Glycyrrhiza uralensis

Abstract
:1. Introduction
2. Results
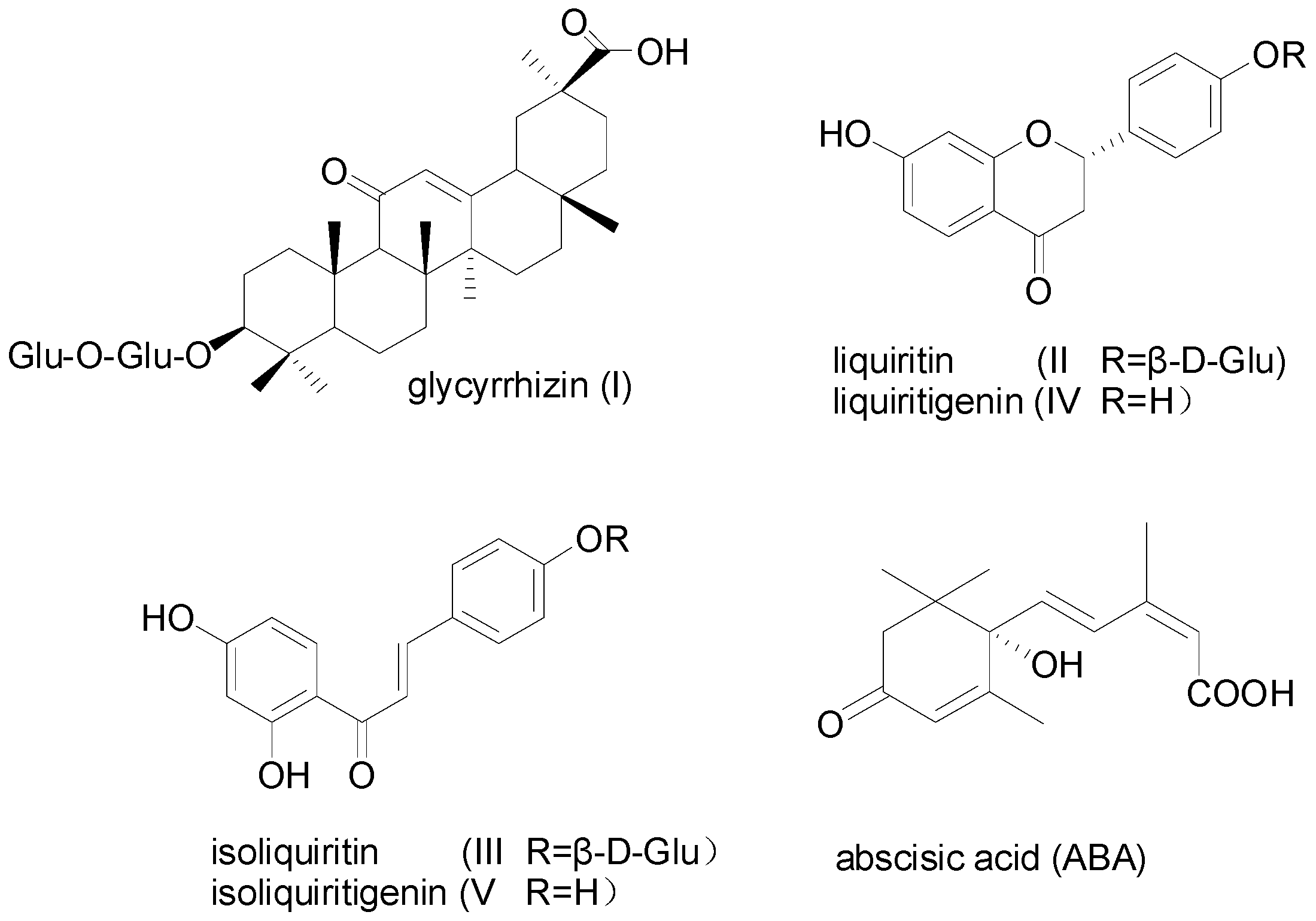
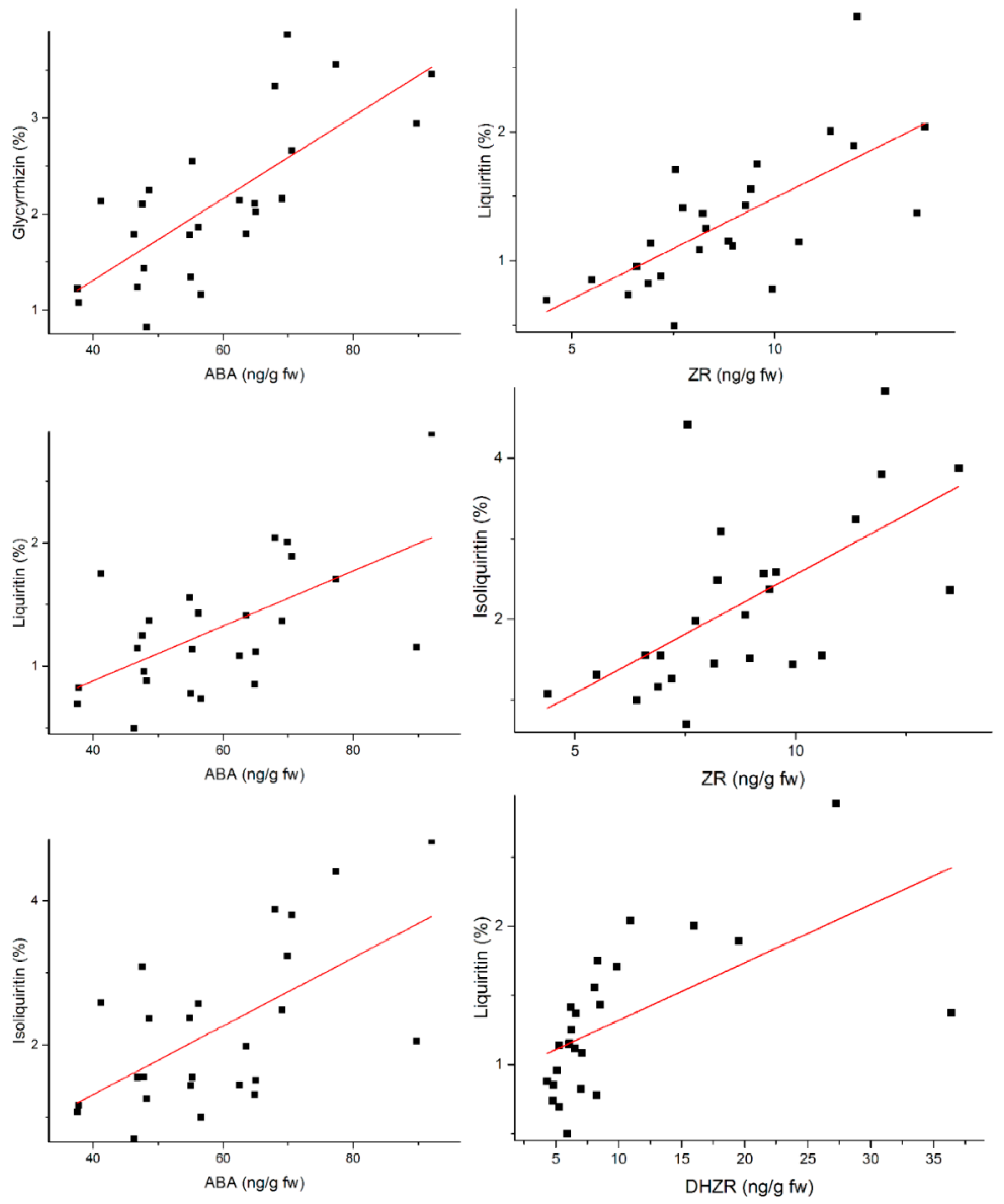
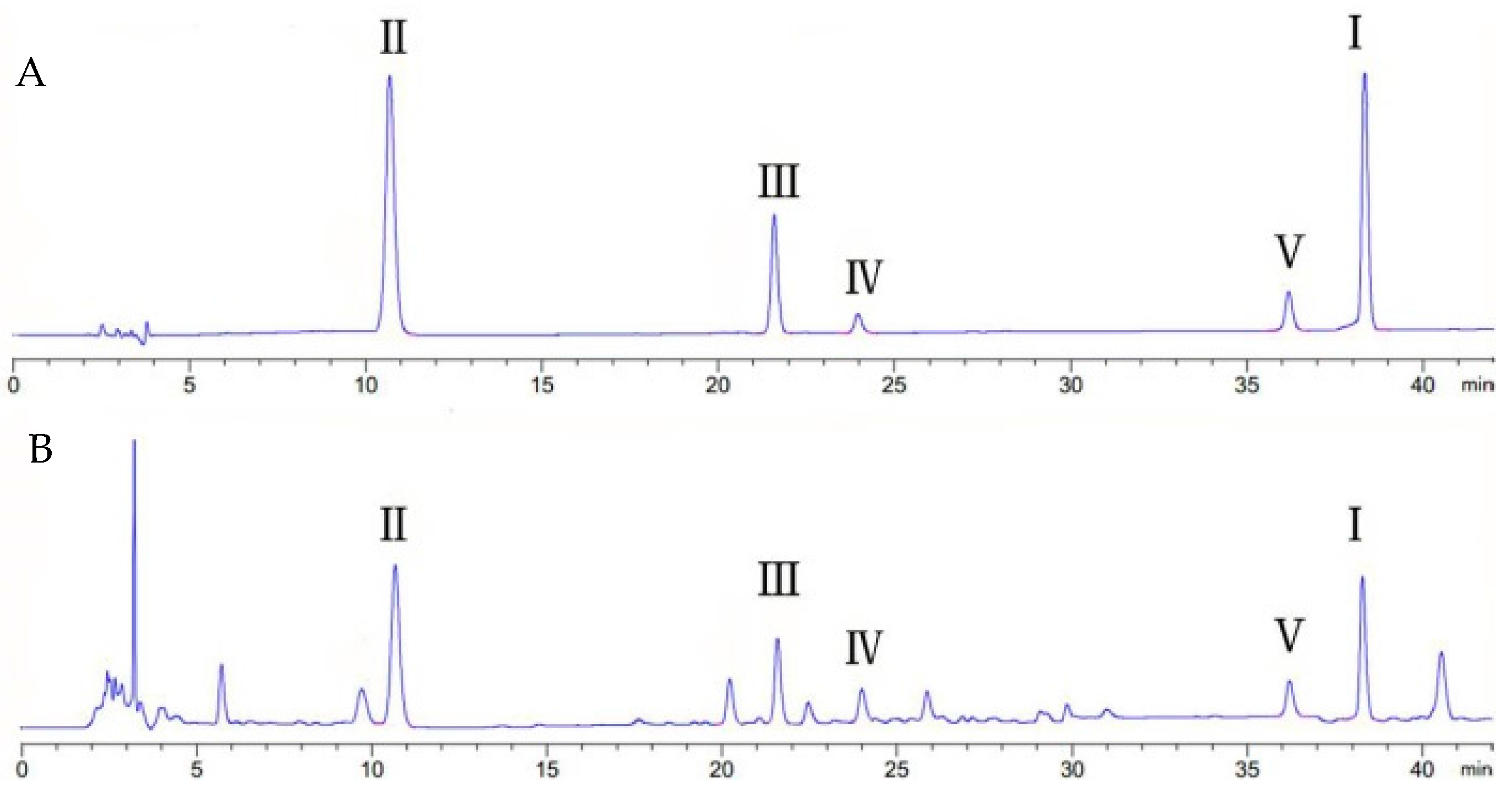
2.1. The Determination of Active Components and Phytohormones in Root of G. uralensis
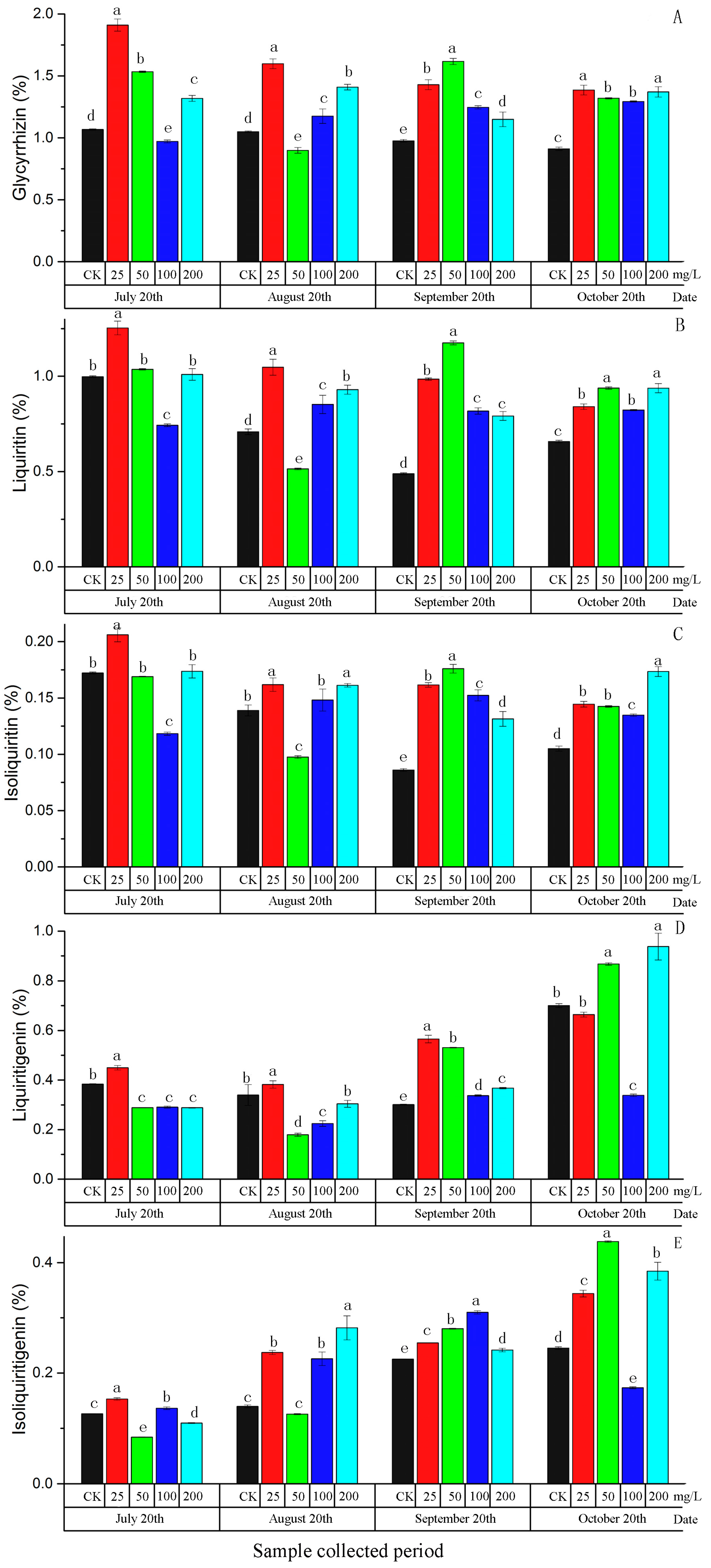
2.2. Morphological Characteristics and the Content of Five Active Components in Root of G. uralensis Affected by Exogenous ABA
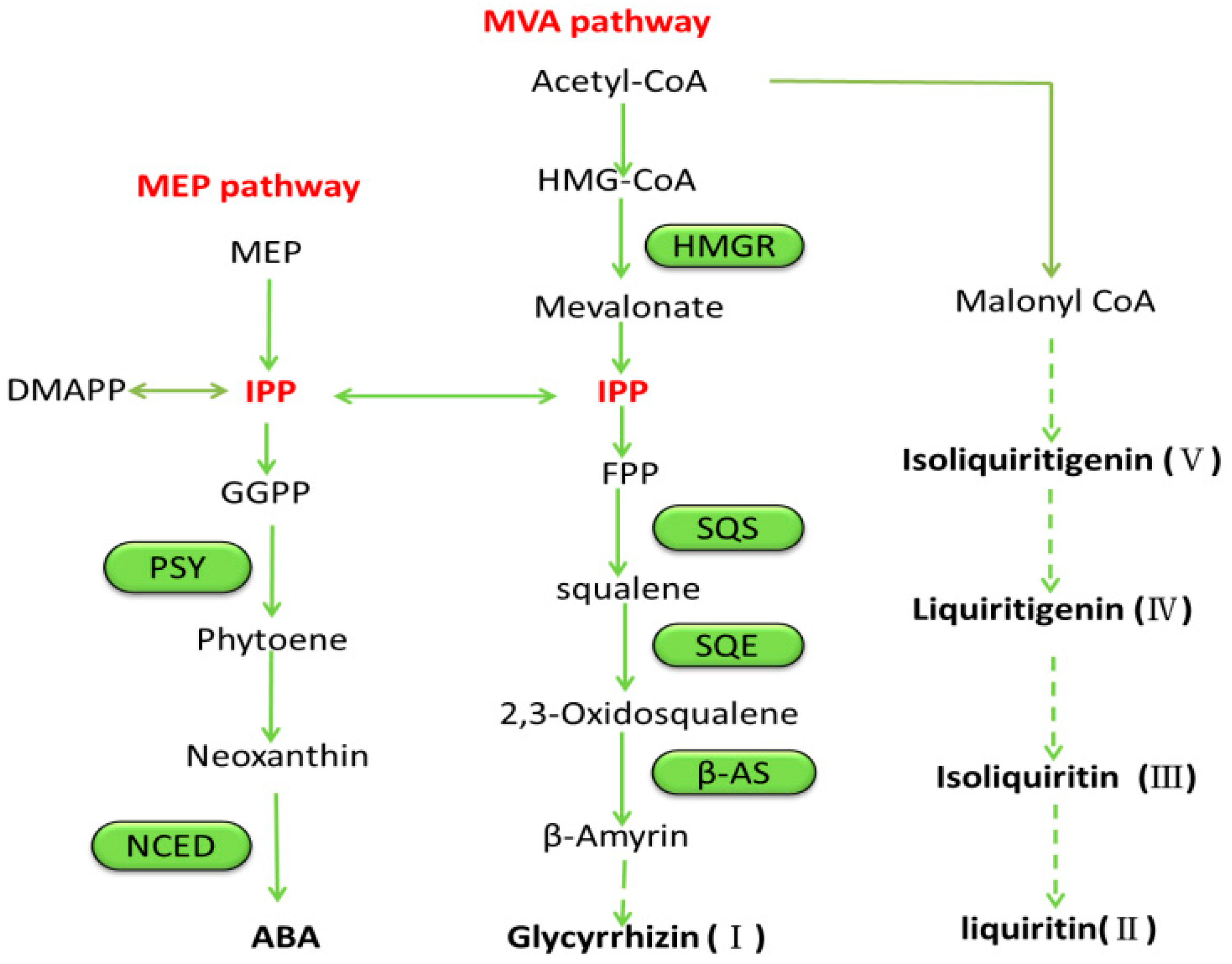
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Materials and ABA Applications
4.3. HPLC Analysis
4.4. Phytohormone Measurements
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China (in Chinese); China Medical Science Press: Beijing, China, 2010; Volume I, pp. 22–23. ISBN 978-75-0674-437-9. [Google Scholar]
- Vaya, J.; Belinky, P.A.; Aviram, M. Antioxidant constituents from licorice roots: Isolation, structure elucidation and antioxidative capacity toward LDL oxidation. Free Radic. Biol. Med. 1997, 23, 302–313. [Google Scholar] [CrossRef]
- Hikino, H. Recent research on oriental medicinal plants. In Economic and Medicinal Plant Research; Academic Press: London, UK, 1985; Volume 1, pp. 204–205. [Google Scholar]
- Seki, H.; Muranaka, T. Triterpene functional genomics in licorice for identification of CYP72A154 involved in the biosynthesis of glycyrrhizin. Plant Cell 2011, 23, 4112–4123. [Google Scholar] [CrossRef] [PubMed]
- Rizzato, G.; Scalabrin, E.; Radaelli, M.; Capodaglio, G.; Piccolo, O. A new exploration of licorice metabolome. Food Chem. 2017, 221, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, X.; Zhao, Z.; Liu, P.; Hu, Y.; Zhou, J.; Wang, Z.; Guo, D.; Guo, H. Antidepressant-like effects of liquiritin and isoliquiritin from Glycyrrhiza uralensis, in the forced swimming test and tail suspension test in mice. Prog. Neuro-Psychopharmacol. 2008, 32, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Kim, S.C.; Kim, Y.W. Composition Comprising Liquiritigenin for Preventing and Treating Liver Disease. Patent Eur. Pat. 1971354A1, 24 September 2008. [Google Scholar]
- Tawata, M.; Aida, K.; Noguchi, T.; Ozaki, Y.; Kume, S.; Sasaki, H.; Chin, M.; Onaya, T. Anti-platelet action of isoliquiritigenin, an aldose reductase inhibitor in licorice. Eur. J. Pharmacol. 1992, 212, 87–92. [Google Scholar] [CrossRef]
- Kimura, Y.; Okuda, T.; Okuda, H. Effects of flavonoids isolated from licorice roots (Glycyrrhiza inflata bat.) on degranulation in human polymorphonuclear neutrophils. Phytother. Res. 1993, 7, 335–340. [Google Scholar] [CrossRef]
- Chen, J.F.; Xu, P.; Zhu, S.Y.; Hu, Y.J. Advance in research for pharmacokinetics and drug interactions of licorice. Chin. J. Clin. Pharm. 2010, 10, 1174–1182. [Google Scholar]
- Miao, X.; Liu, R.; Liu, H.; Yu, F.; Wang, J.; Wang, W.; Wei, S. Genetic and environmental effect on the growth characteristics and bioactive components of eight-year-old Glycyrrhiza uralensis fisch. Agri Gene 2017, 3, 57–62. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Tani, T. Growth and glycyrrhizin contents in Glycyrrhiza uralensis roots cultivated for four years in eastern nei-meng-gu of china. J. Tradit. Med. 2002, 19, 87–92. [Google Scholar]
- Su, W.H.; Zhang, G.F.; Liu, X.H.; Ou, X.K. Relationship between accumulation of secondary metabolism in medicinal plant and environmental condition. Chin. Tradit. 2005, 9, 1415–1418. [Google Scholar]
- Li, W.D.; Hou, J.L.; Wang, W.Q.; Tang, X.M.; Liu, C.L.; Xing, D. Effect of water deficit on biomass production and accumulation of secondary metabolites in roots of Glycyrrhiza uralensis. Russ. J. Plant Physiol. 2011, 583, 538–542. [Google Scholar] [CrossRef]
- Daszkowskagolec, A.; Szarejko, I. Open or close the gate—Stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar]
- Seo, M.; Koshiba, T. Complex regulation of aba biosynthesis in plants. Trends Plant Sci. 2002, 7, 41–48. [Google Scholar] [CrossRef]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. Omics approaches toward defining the comprehensive abscisic acid signaling network in plants. Plant Cell Physiol. 2015, 56, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, A.K.; Gray, D.J.; Lu, J.; Gu, L. Effects of exogenous abscisic acid on antioxidant capacities, anthocyanins, and flavonol contents of muscadine grape (vitis rotundifolia) skins. Food Chem. 2011, 126, 982–988. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, X.; Sandhu, A.K.; Gu, L.W. Effects of exogenous abscisic acid on yield, antioxidant capacities, and phytochemical contents of greenhouse grown lettuces. J. Agric. Food Chem. 2010, 58, 6503–6509. [Google Scholar] [CrossRef] [PubMed]
- Kojoma, M.; Hayashi, S.; Shibata, T.; Yamamoto, Y.; Sekizaki, H. Variation of glycyrrhizin and liquiritin contents within a population of 5-year-old licorice (Glycyrrhiza uralensis) plants cultivated under the same conditions. Biol. Pharm. Bull. 2011, 34, 1334–1337. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Wu, L.J.; Yu, Z.L. Effect of salt and drought stress on antioxidant enzymes activities and sod isoenzymes of liquorice (Glycyrrhiza uralensis fisch). Plant Growth Regul. 2006, 49, 157–165. [Google Scholar] [CrossRef]
- Nasrollahi, V.; Mirzaie-Asl, A.; Piri, K.; Nazeri, S.; Mehrabi, R. The effect of drought stress on the expression of key genes involved in the biosynthesis of triterpenoid saponins in liquorice (Glycyrrhiza glabra). Phytochemistry 2014, 103, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Do Nascimento, N.C.; Menguer, P.K.; Henriques, A.T.; Fett-Neto, A.G. Accumulation of brachycerine, an antioxidant glucosidic indole alkaloid, is induced by abscisic acid, heavy metal, and osmotic stress inleaves of psychotria brachyceras. Plant Physiol. Biochem. 2013, 73, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.X.; Lv, B.S.; Wang, M.M.; Ma, H.Y.; Yang, H.Y.; Liu, X.L.; Jiang, C.J.; Liang, Z.W. Priming effect of abscisic acid on alkaline stress tolerance in rice (Oryza sativa L.) seedlings. Plant Physiol. Biochem. 2015, 90, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Balint, G.; Reynolds, A.G. Impact of exogenous abscisic acid on vine physiology and grape composition of cabernet sauvignon. Am. J. Enol. Vitic. 2012, 64, 74–87. [Google Scholar] [CrossRef]
- Gagné, S.; Cluzet, S.; Mérillon, J.M.; Gény, L. ABAinitiates anthocyanin production in grape cell cultures. J. Plant Growth Regul. 2011, 30, 1–10. [Google Scholar] [CrossRef]
- Ma, L.N.; Xi, Z.M.; Huo, S.S.; Luan, L.Y.; Gao, X.; Zhao, X.L. Studies on the regulation of anthocyanin biosynthesis in grape berry by brassinosteroid and abscisic acid. J. Fruit Sci. 2012, 5, 830–836. [Google Scholar]
- Leung, J.; Giraudat, J. Abscisic acid signal transduction. Annu. Rev. Plant Phys. 1998, 49, 199–222. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.P.; Yu, C.X.; Qiao, J.; Zang, Y.M.; Xiang, Y.; Ren, G.X.; Wang, L.; Zhang, X.Y.; Liu, C.S. Effect of exogenous phytohormones treatment on glycyrrhizic acid accumulation and preliminary exploration of the chemical control network based on glycyrrhizic acid in root of Glycyrrhiza uralensis. Rev. Bras. Farmacogn. 2016, 26, 490–496. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds glycyrrhizin (I), liquiritin (II), isoliquiritin (III), liquiritigenin (IV), isoliquiritigenin (V) are available from the corresponding author. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | I | II | III | IV | V | ZR | iPA | DHZR | GA3 | MeJA | ABA | BR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3.46 | 2.89 | 4.83 | 0.10 | 0.03 | 12.03 | 10.50 | 27.26 | 14.39 | 43.67 | 92.04 | 8.38 |
| 2 | 2.66 | 1.89 | 3.80 | 0.14 | 0.03 | 11.94 | 5.92 | 19.49 | 12.34 | 40.95 | 70.57 | 10.51 |
| 3 | 3.33 | 2.04 | 3.88 | 0.08 | 0.03 | 13.69 | 10.14 | 10.91 | 12.59 | 28.68 | 67.97 | 7.83 |
| 4 | 2.10 | 1.25 | 3.09 | 0.04 | 0.01 | 8.31 | 5.50 | 6.23 | 7.36 | 49.18 | 47.55 | 8.95 |
| 5 | 1.43 | 0.96 | 1.55 | 0.13 | 0.03 | 6.59 | 4.65 | 5.09 | 8.43 | 24.59 | 47.77 | 8.68 |
| 6 | 2.15 | 1.08 | 1.45 | 0.07 | 0.02 | 8.15 | 5.64 | 7.06 | 7.63 | 31.63 | 62.45 | 9.22 |
| 7 | 2.11 | 0.85 | 1.31 | 0.08 | 0.02 | 5.50 | 5.52 | 4.83 | 6.84 | 29.90 | 64.85 | 8.55 |
| 8 | 1.08 | 0.82 | 1.16 | 0.06 | 0.01 | 6.88 | 5.42 | 7.00 | 6.50 | 25.31 | 37.77 | 7.65 |
| 9 | 2.14 | 1.75 | 2.59 | 0.07 | 0.03 | 9.57 | 6.16 | 8.35 | 9.85 | 29.29 | 41.20 | 8.95 |
| 10 | 2.55 | 1.14 | 1.55 | 0.07 | 0.02 | 6.94 | 6.74 | 5.27 | 11.68 | 27.40 | 55.27 | 7.17 |
| 11 | 1.34 | 0.78 | 1.44 | 0.03 | 0.02 | 9.94 | 5.36 | 8.25 | 8.29 | 23.83 | 55.01 | 7.87 |
| 12 | 3.56 | 1.71 | 4.41 | 0.13 | 0.03 | 7.56 | 5.64 | 9.86 | 12.01 | 27.51 | 77.34 | 9.27 |
| 13 | 2.16 | 1.37 | 2.48 | 0.08 | 0.02 | 8.23 | 5.40 | 6.59 | 8.14 | 31.76 | 69.09 | 9.29 |
| 14 | 1.16 | 0.74 | 1.00 | 0.06 | 0.02 | 6.39 | 4.83 | 4.78 | 5.57 | 14.40 | 56.58 | 8.21 |
| 15 | 1.86 | 1.43 | 2.57 | 0.09 | 0.02 | 9.28 | 8.79 | 8.53 | 12.29 | 41.98 | 56.18 | 9.36 |
| 16 | 2.94 | 1.15 | 2.05 | 0.10 | 0.02 | 8.85 | 5.58 | 6.06 | 6.97 | 27.28 | 89.69 | 9.20 |
| 17 | 0.82 | 0.88 | 1.26 | 0.04 | 0.01 | 7.19 | 5.35 | 4.33 | 6.10 | 34.23 | 48.23 | 8.30 |
| 18 | 1.24 | 1.15 | 1.55 | 0.20 | 0.03 | 10.59 | 11.26 | 6.00 | 10.65 | 29.04 | 46.77 | 9.79 |
| 19 | 2.02 | 1.12 | 1.51 | 0.13 | 0.02 | 8.96 | 4.68 | 6.51 | 8.79 | 37.05 | 65.00 | 7.91 |
| 20 | 2.25 | 1.37 | 2.36 | 0.09 | 0.02 | 13.50 | 22.60 | 36.41 | 25.54 | 55.95 | 48.57 | 8.64 |
| 21 | 3.87 | 2.01 | 3.24 | 0.27 | 0.04 | 11.36 | 11.24 | 15.99 | 21.81 | 37.44 | 69.91 | 9.84 |
| 22 | 1.22 | 0.70 | 1.07 | 0.19 | 0.03 | 4.39 | 6.80 | 5.25 | 6.39 | 21.57 | 37.59 | 4.27 |
| 23 | 1.79 | 0.50 | 0.70 | 0.03 | 0.01 | 7.52 | 7.20 | 5.89 | 11.10 | 21.88 | 46.34 | 10.07 |
| 24 | 1.80 | 1.41 | 1.98 | 0.13 | 0.02 | 7.74 | 4.55 | 6.18 | 8.82 | 28.92 | 63.49 | 8.49 |
| 25 | 1.78 | 1.56 | 2.37 | 0.14 | 0.03 | 9.41 | 8.12 | 8.08 | 9.18 | 30.09 | 54.88 | 6.03 |
| Pearson Coefficient | ZR | iPA | DHZR | GA3 | MeJA | ABA | BR |
|---|---|---|---|---|---|---|---|
| I | 0.519 ** | 0.258 | 0.460 * | 0.560 ** | 0.343 | 0.746 ** | 0.318 |
| II | 0.693 ** | 0.325 | 0.600 ** | 0.519 ** | 0.496 * | 0.601 ** | 0.209 |
| III | 0.625 ** | 0.253 | 0.552 ** | 0.464 * | 0.506 ** | 0.609 ** | 0.271 |
| IV | 0.162 | 0.230 | 0.156 | 0.371 | 0.062 | 0.160 | −0.042 |
| V | 0.374 | 0.232 | 0.286 | 0.402 * | −0.014 | 0.306 | −0.043 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, J.; Luo, Z.; Li, Y.; Ren, G.; Liu, C.; Ma, X. Effect of Abscisic Acid on Accumulation of Five Active Components in Root of Glycyrrhiza uralensis. Molecules 2017, 22, 1982. https://doi.org/10.3390/molecules22111982
Qiao J, Luo Z, Li Y, Ren G, Liu C, Ma X. Effect of Abscisic Acid on Accumulation of Five Active Components in Root of Glycyrrhiza uralensis. Molecules. 2017; 22(11):1982. https://doi.org/10.3390/molecules22111982
Chicago/Turabian StyleQiao, Jing, Zuliang Luo, Yanpeng Li, Guangxi Ren, Chunsheng Liu, and Xiaojun Ma. 2017. "Effect of Abscisic Acid on Accumulation of Five Active Components in Root of Glycyrrhiza uralensis" Molecules 22, no. 11: 1982. https://doi.org/10.3390/molecules22111982
APA StyleQiao, J., Luo, Z., Li, Y., Ren, G., Liu, C., & Ma, X. (2017). Effect of Abscisic Acid on Accumulation of Five Active Components in Root of Glycyrrhiza uralensis. Molecules, 22(11), 1982. https://doi.org/10.3390/molecules22111982