Modulation of Cytochrome P450, P-glycoprotein and Pregnane X Receptor by Selected Antimalarial Herbs—Implication for Herb-Drug Interaction

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
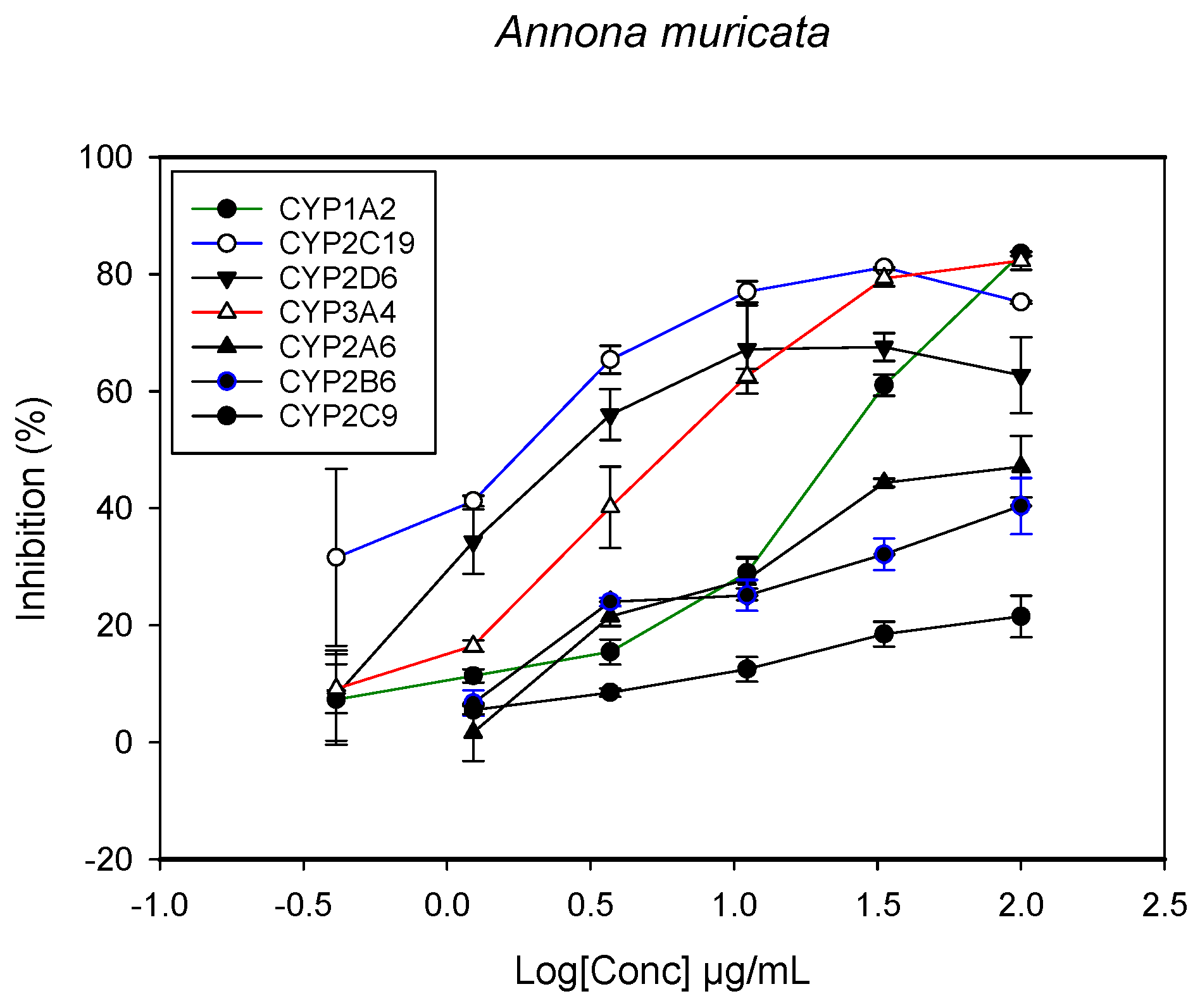
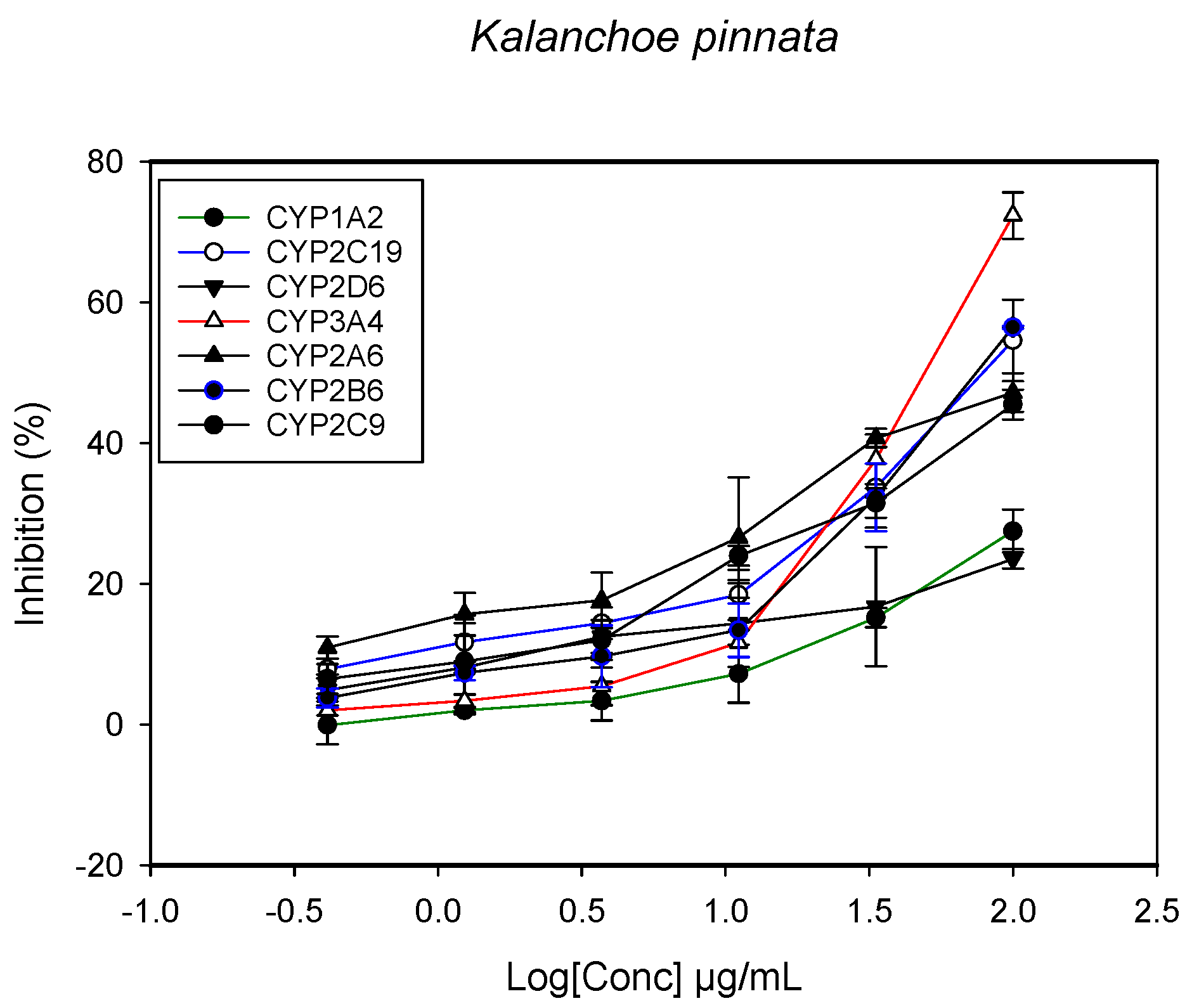
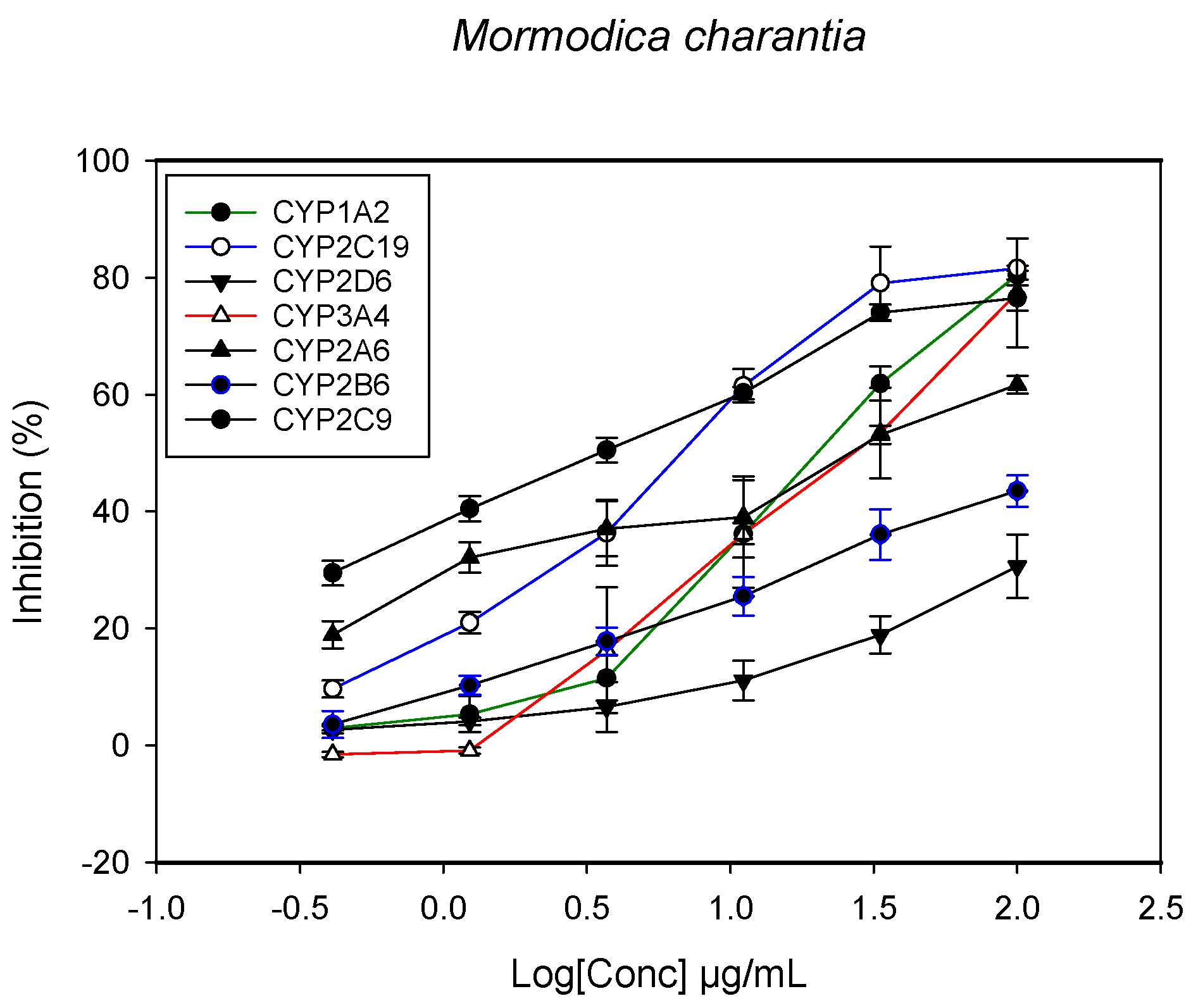
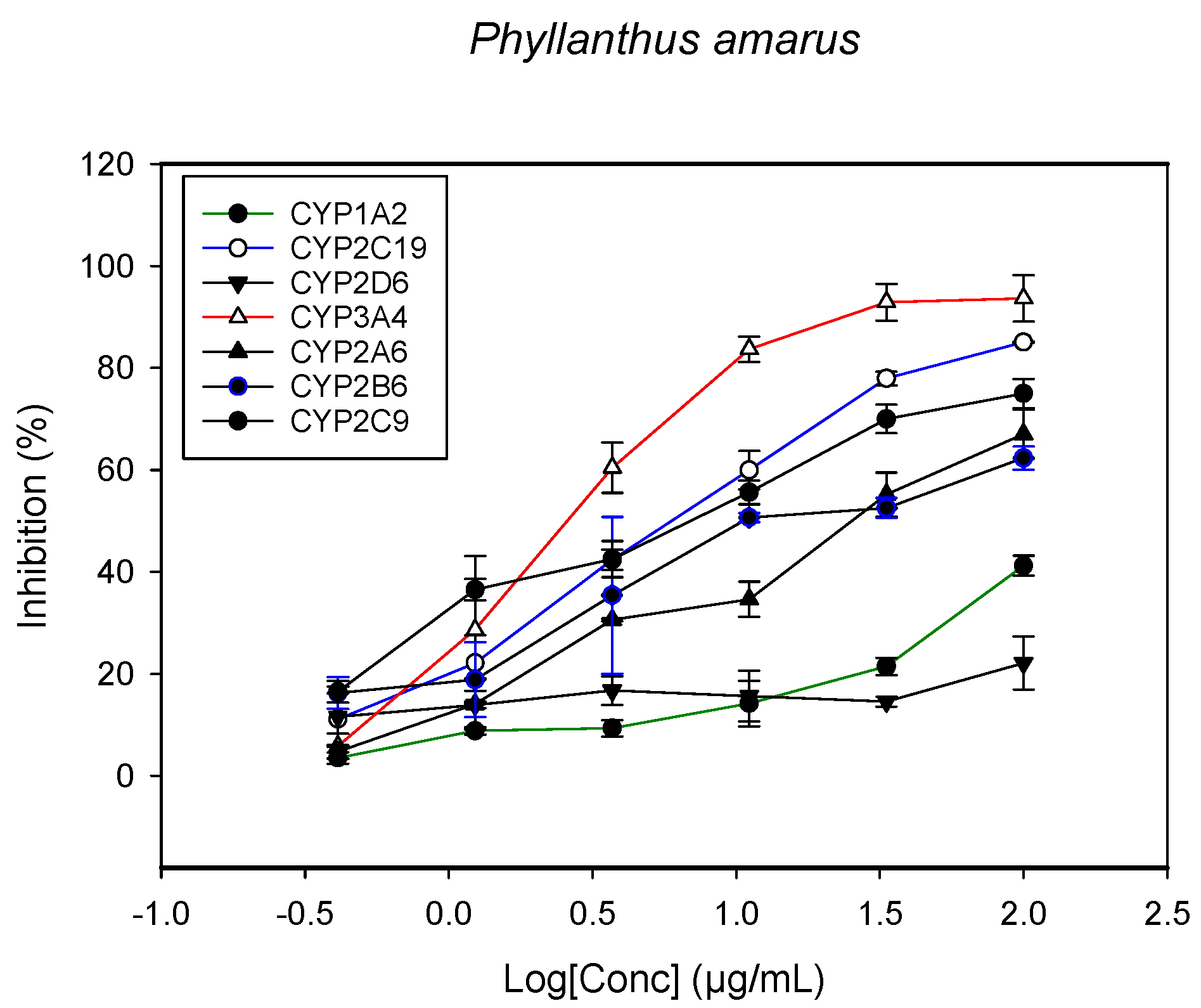
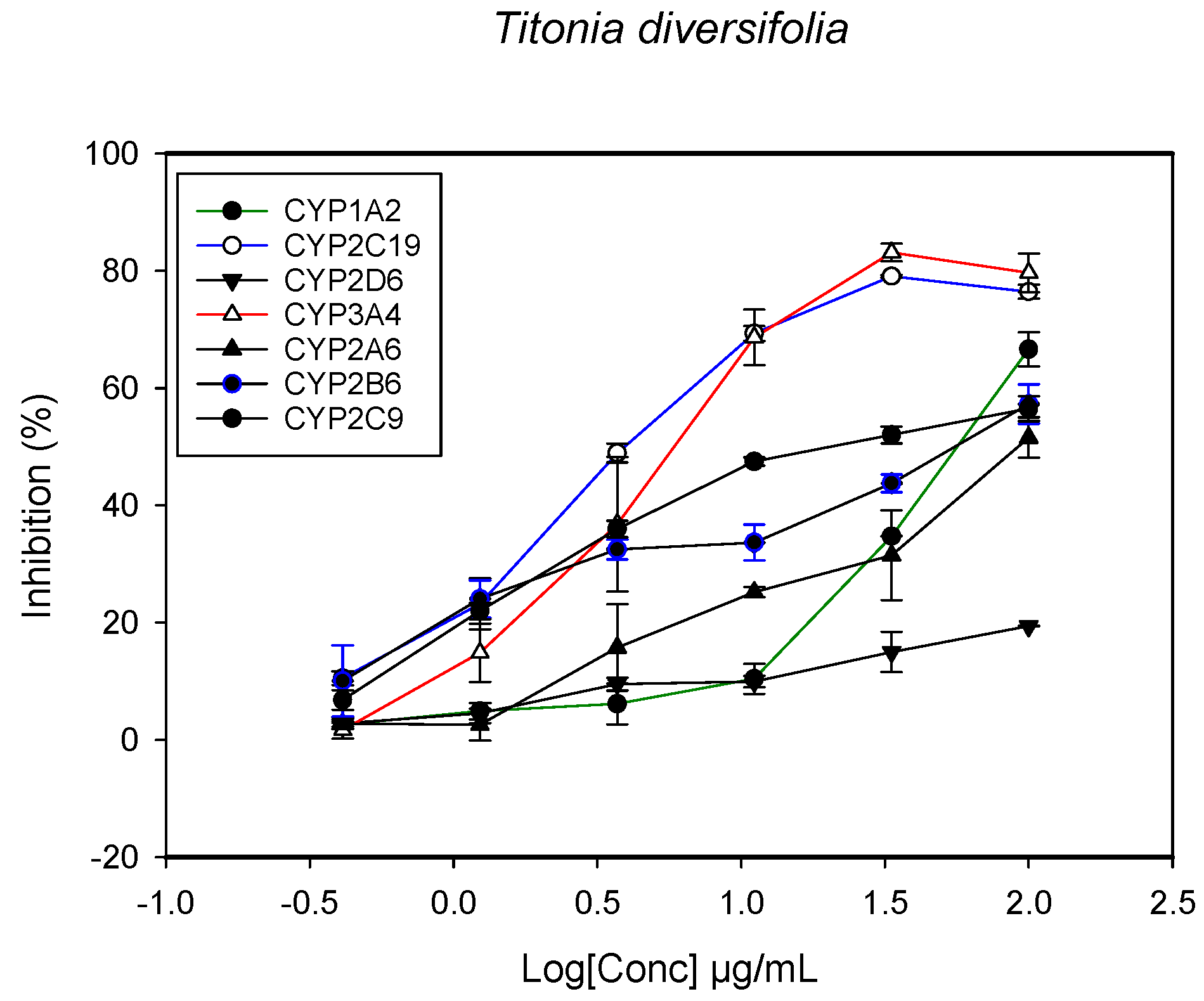
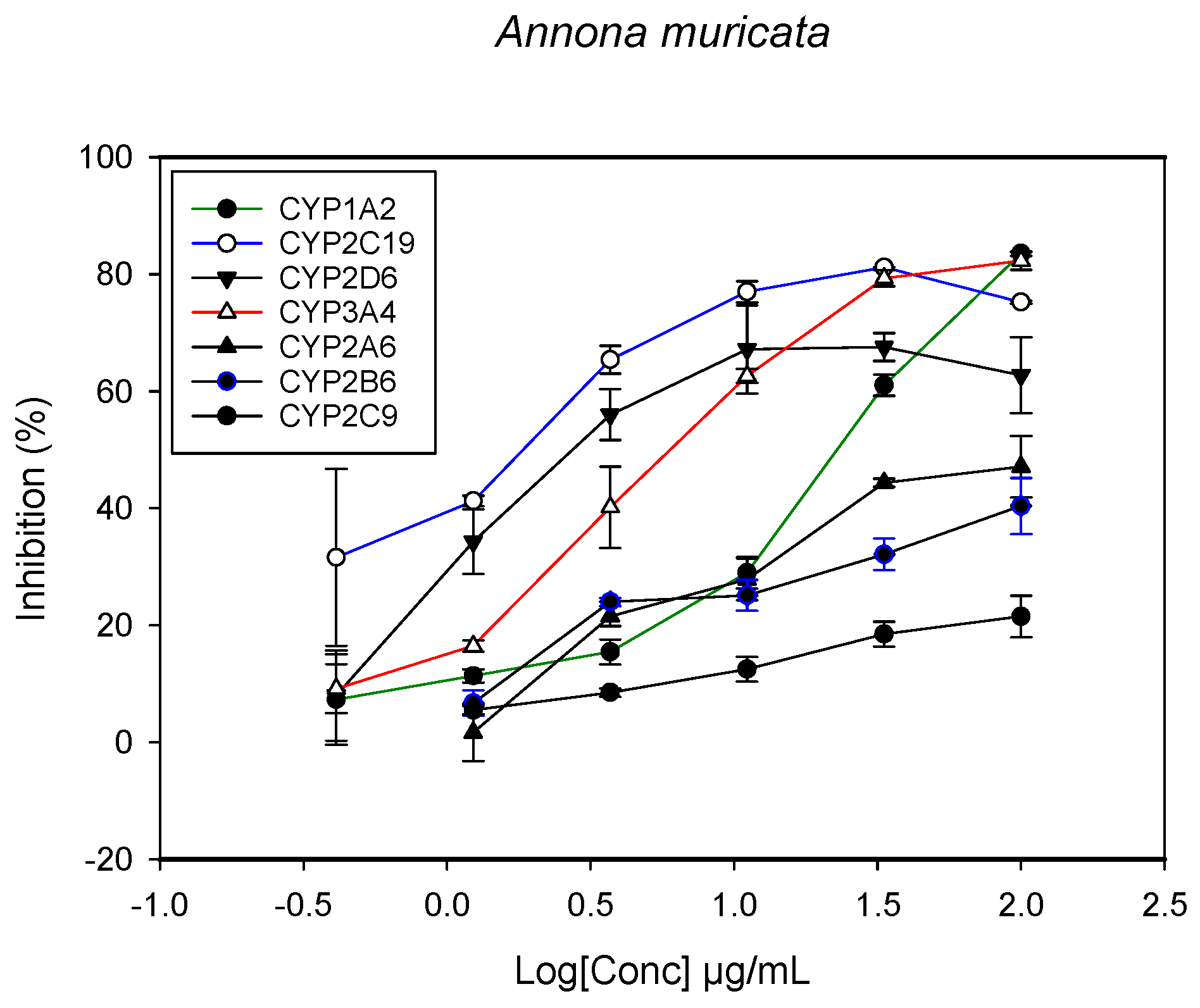
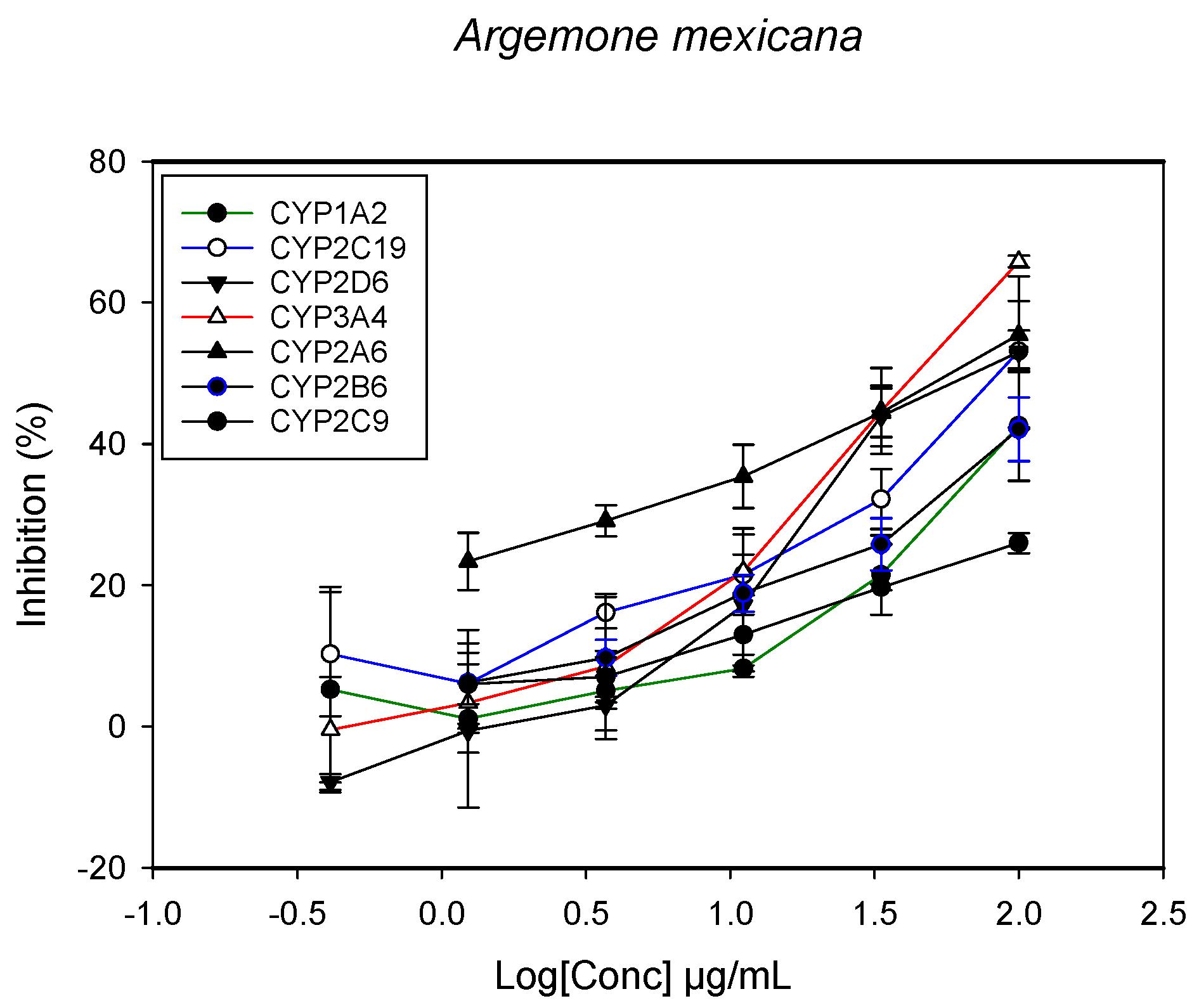
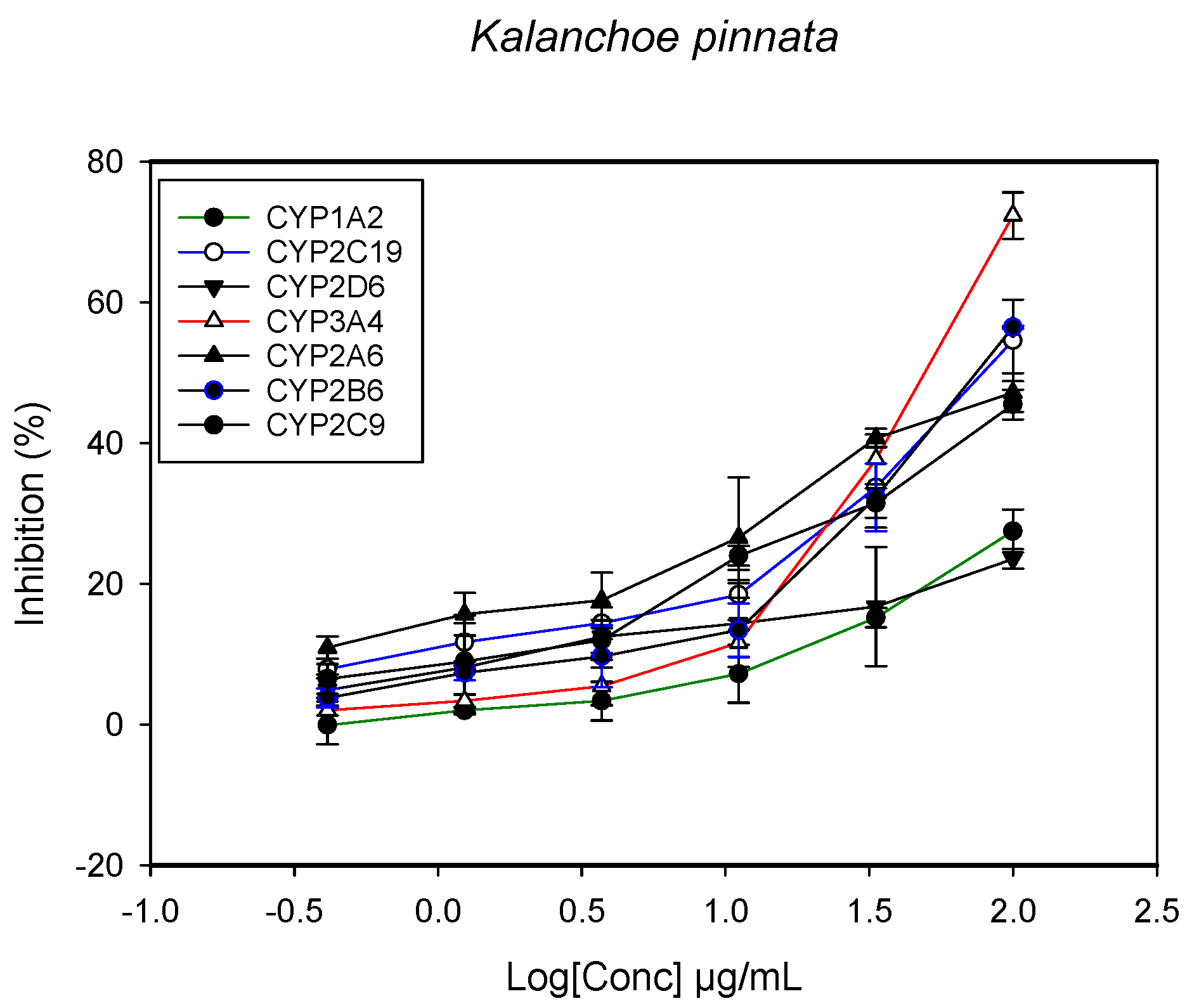
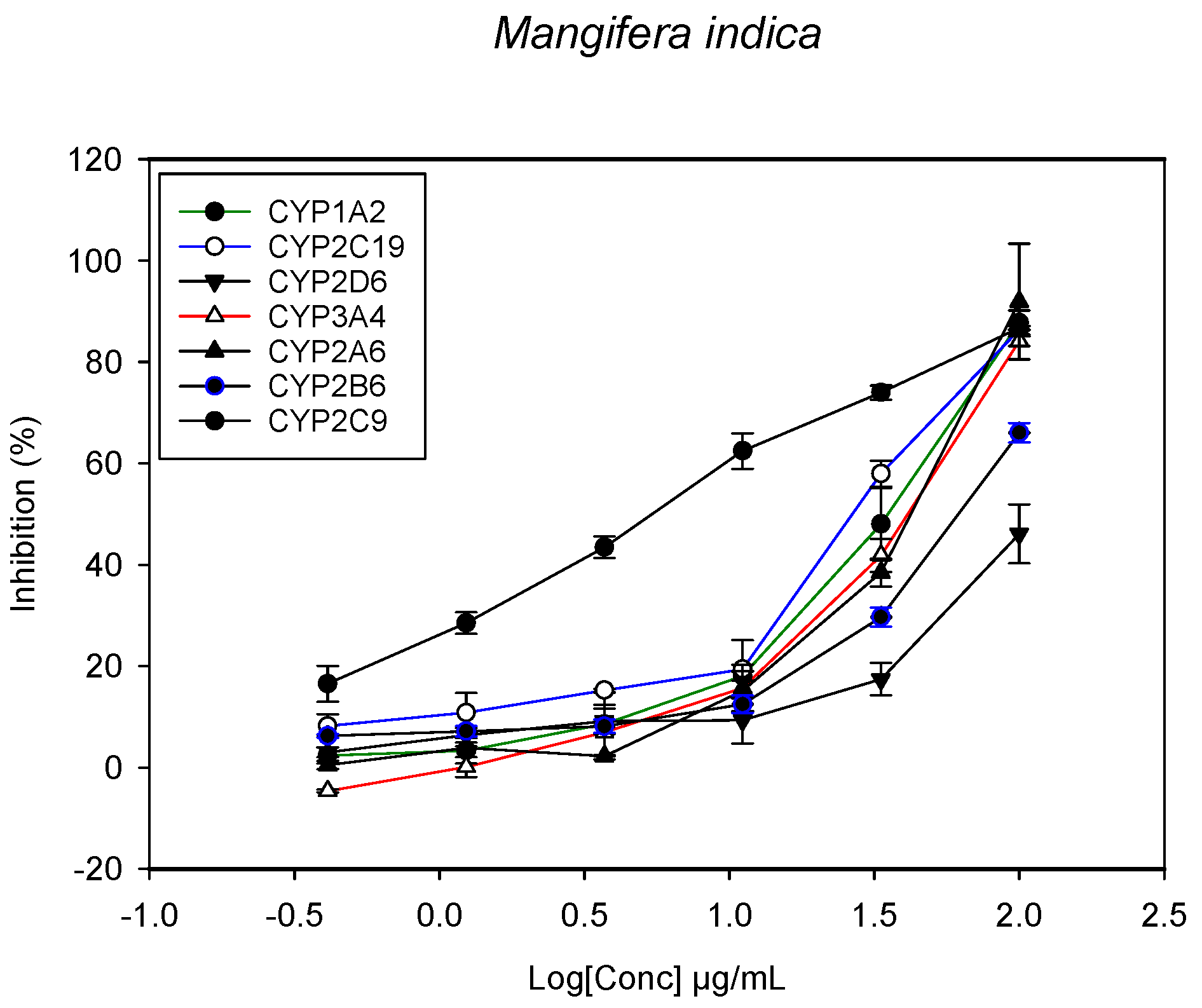
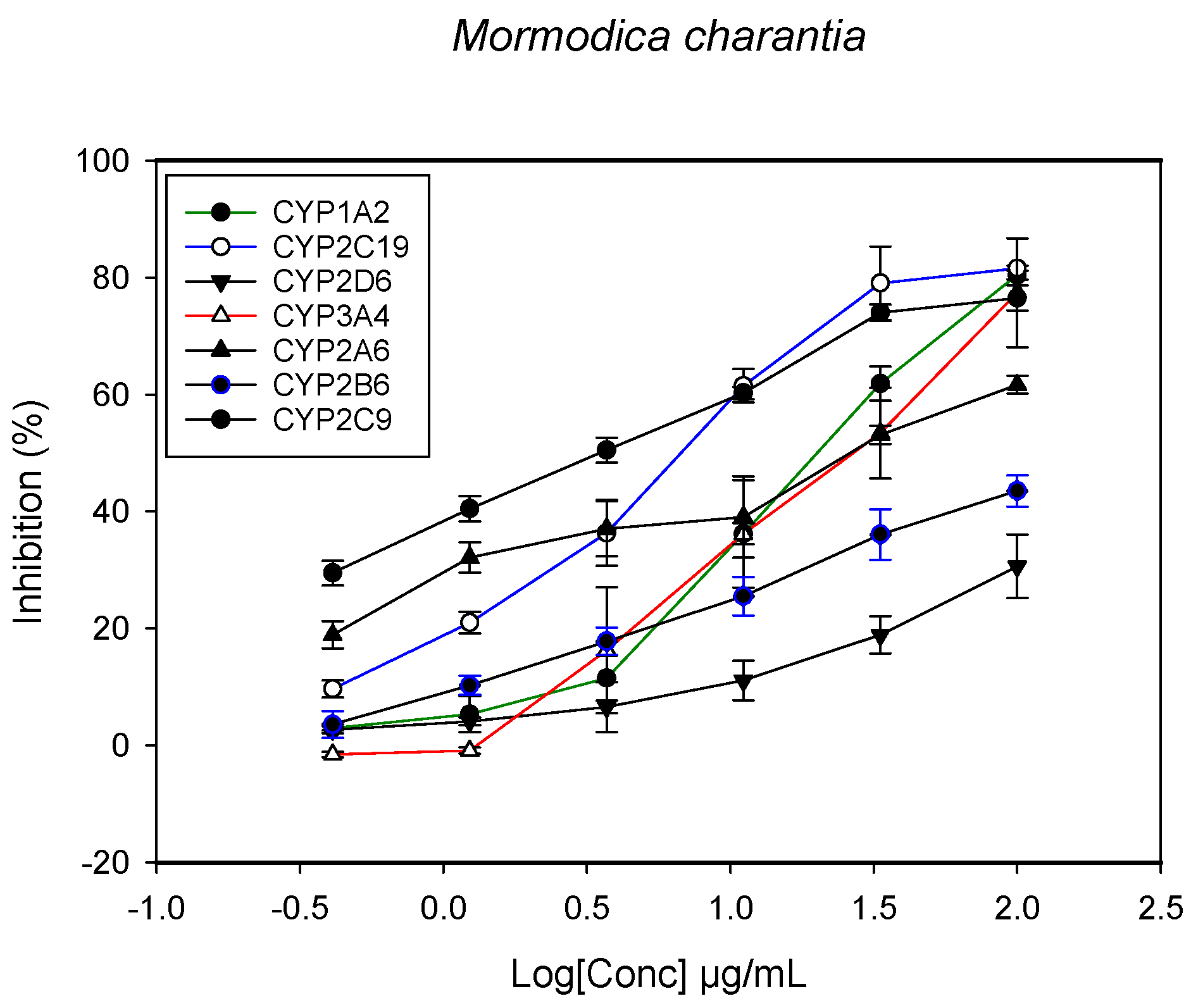
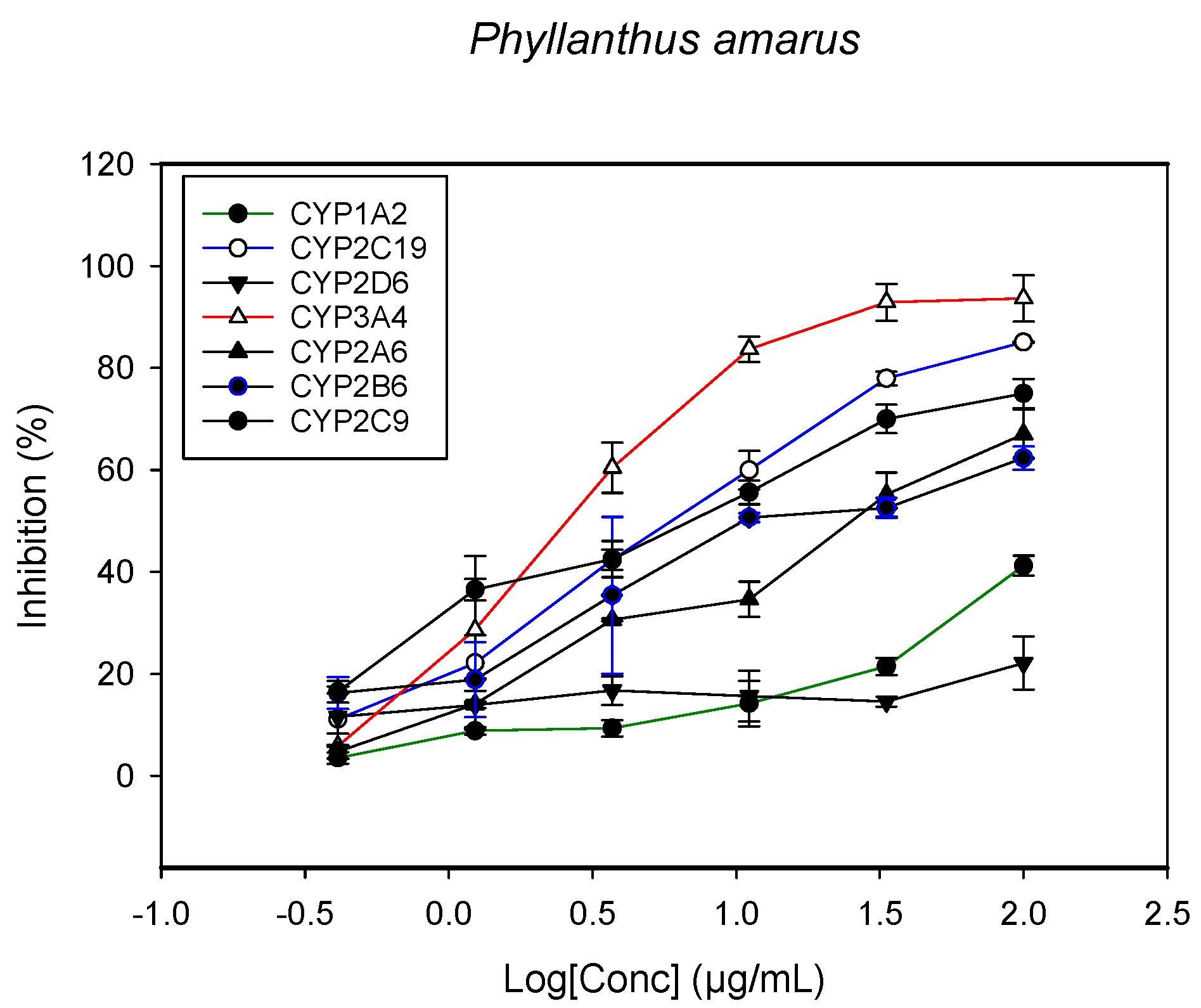
2.1. Inhibition of CYPs by the Herbal Extracts
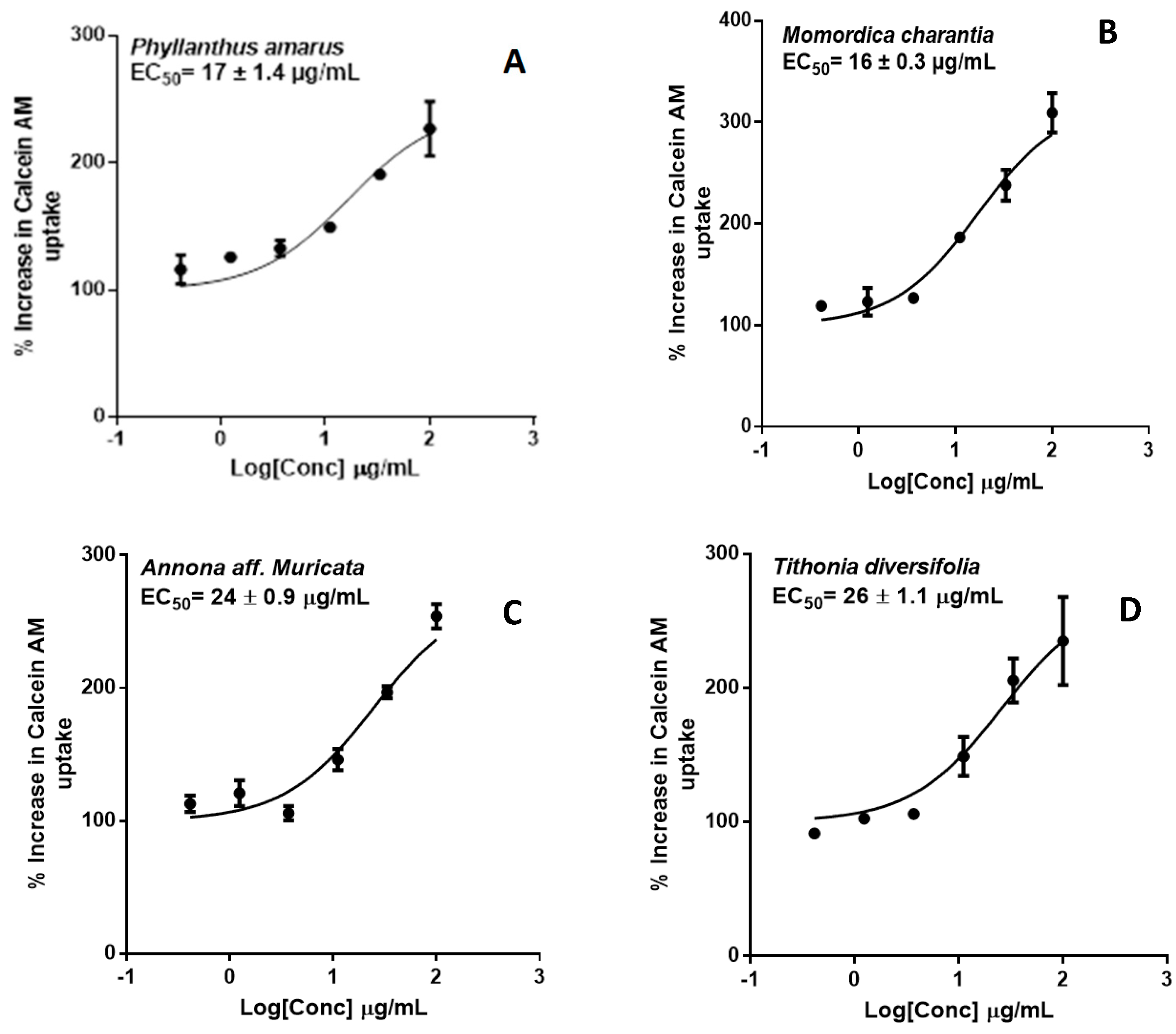
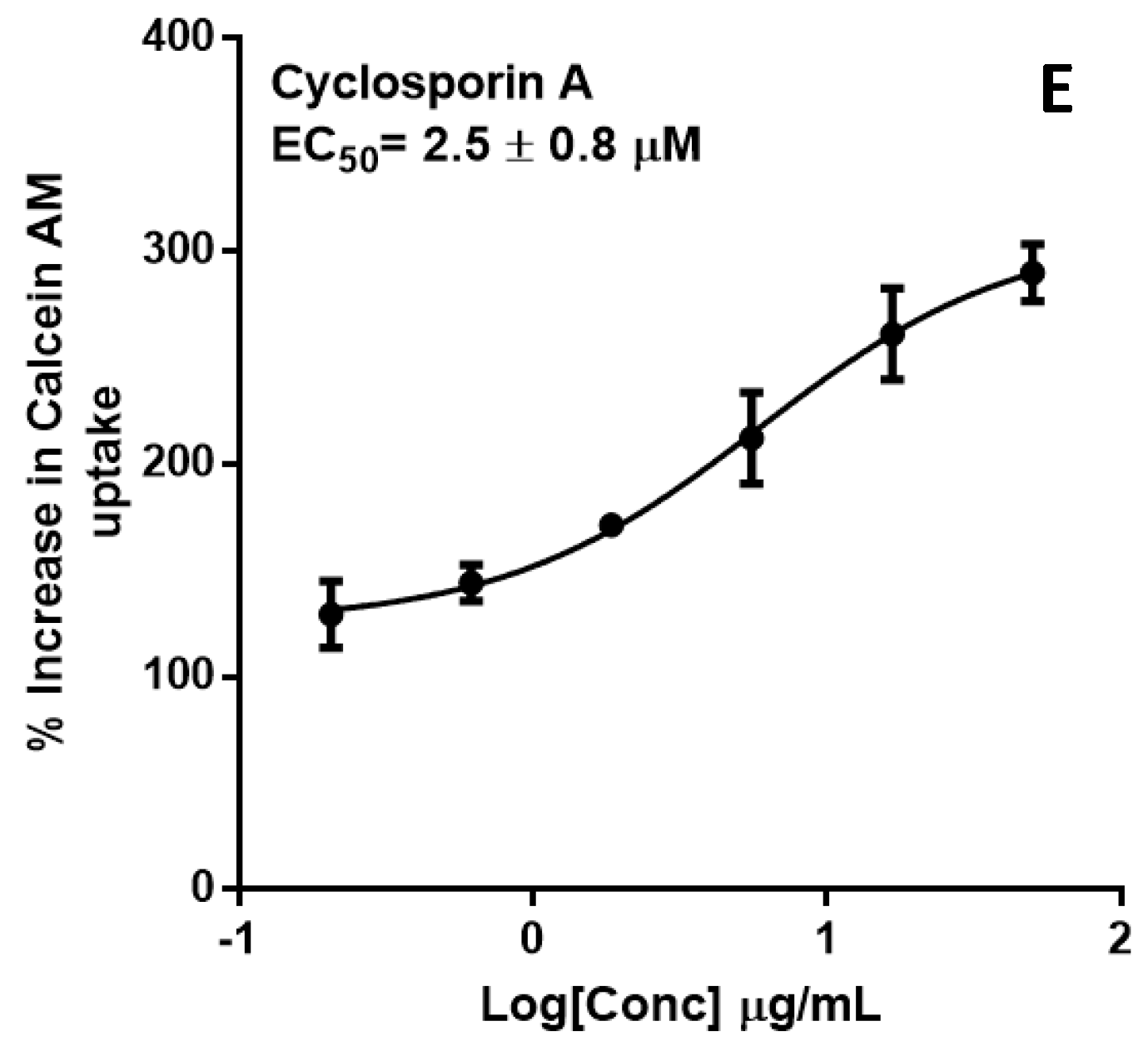
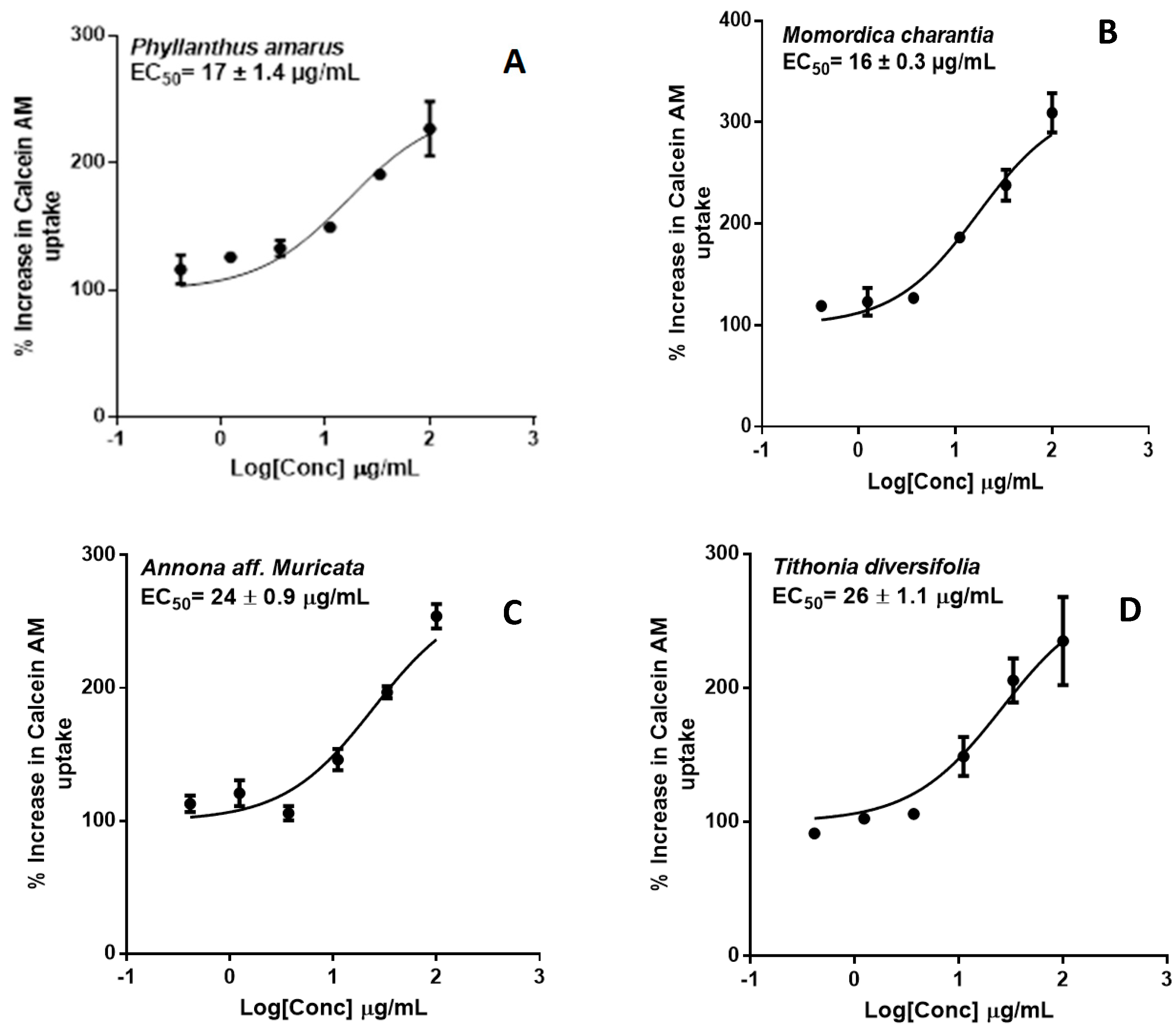
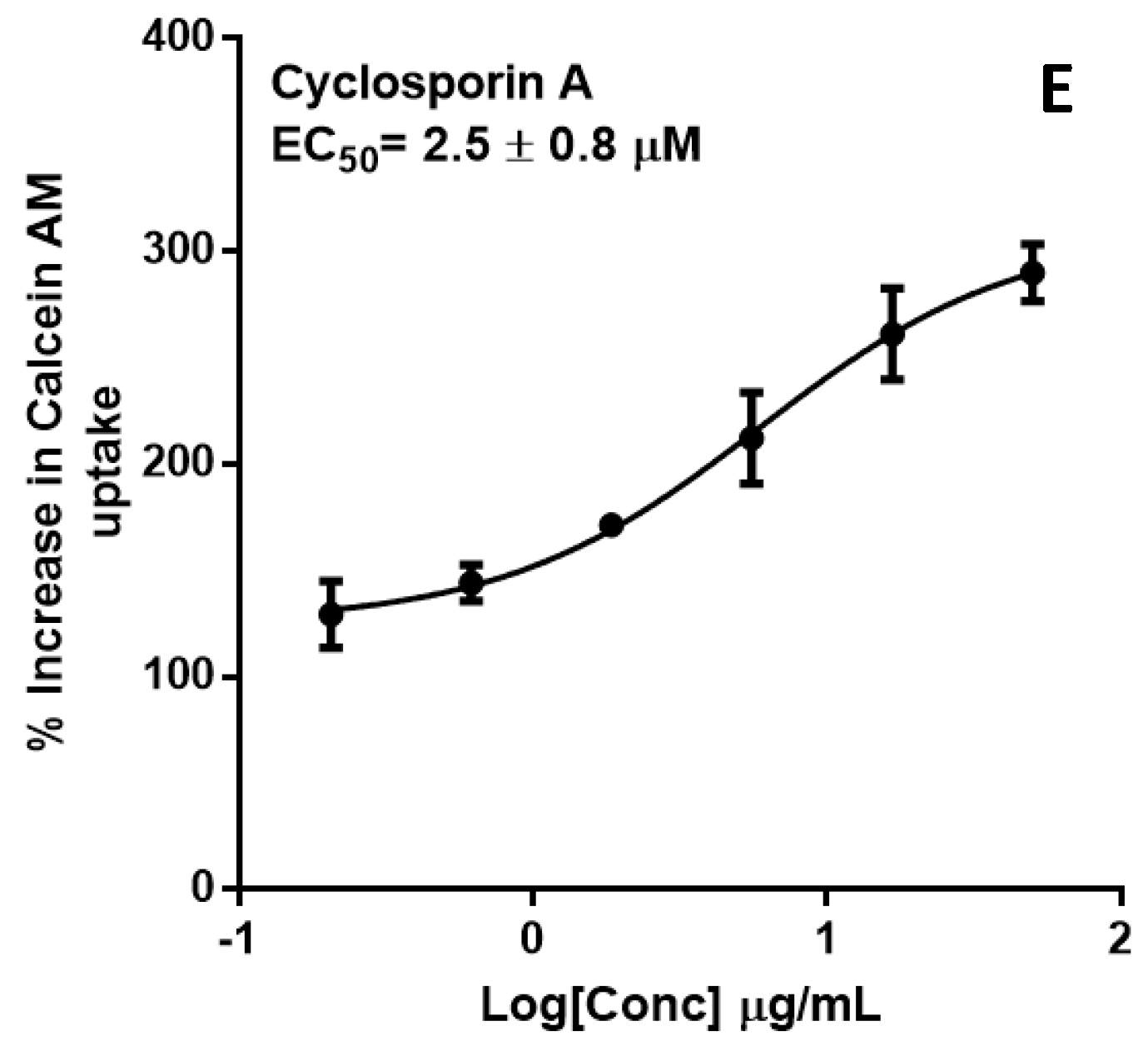
2.2. P-glycoprotein Inhibition
2.3. Pregnane-X Receptor Modulation
2.4. Discussion
3. Materials and Methods
3.1. Materials
3.2. Inhibition Assay with Recombinant CYPs
3.3. Assay for P-glycoprotein Inhibition by Calcein-AM Uptake in Parental and Transfected MDCK-II Cells
3.4. Assay for PXR Modulation
3.5. Calculation of IC50 Inhibition Values
3.6. Statistical Methods
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Fact Sheet on the World Malaria Report. 2016. Available online: http://www.who.int/malaria/publications/world-malaria-report-2016/report/en/ (accessed on 22 November 2017).
- Komlaga, G.; Agyare, C.; Dickson, R.A.; Mensah, M.L.; Annan, K.; Loiseau, P.M.; Champy, P. Medicinal plants and finished marketed herbal products used in the treatment of malaria in the Ashanti region, Ghana. J. Ethnopharmacol. 2015, 172, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Ahorlu, C.K.; Dunyo, S.K.; Afari, E.A.; Koram, K.A.; Nkrumah, F.K. Malaria-related beliefs and behaviour in Southern Ghana: Implications for treatment, prevention and control. Trop. Med. Int. Health 1997, 5, 488–499. [Google Scholar] [CrossRef]
- Fasinu, P.S.; Bouic, P.J.; Rosenkranz, B. An overview of the evidence and mechanisms of herb–drug interactions. Front. Pharmacol. 2012, 3, 69. [Google Scholar] [CrossRef] [PubMed]
- Fasinu, P.S.; Gurley, B.J.; Walker, L.A. Clinically relevant pharmacokinetic herb-drug interactions in antiretroviral therapy. Curr. Drug Metab. 2016, 17, 52–64. [Google Scholar] [CrossRef]
- Morris, C.A.; Duparc, S.; Borghini-Fuhrer, I.; Jung, D.; Shin, C.S.; Fleckenstein, L. Review of the clinical pharmacokinetics of artesunate and its active metabolite dihydroartemisinin following intravenous, intramuscular, oral or rectal administration. Malar. J. 2011, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Fasinu, P.; Bouic, P.J.; Rosenkranz, B. Liver-based in vitro technologies for drug biotransformation studies—A review. Curr. Drug Metab. 2012, 13, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ménan, H.; Banzouzi, J.T.; Hocquette, A.; Pélissier, Y.; Blache, Y.; Koné, M.; Mallié, M.; Assi, L.A.; Valentin, A. Antiplasmodial activity and cytotoxicity of plants used in West African traditional medicine for the treatment of Malaria. J. Ethnopharmacol. 2006, 105, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Watt, J.M.; Breyer-Brandwijk, M.J. The Medicinal and Poisonous Plants of Southern and Eastern Afica, 2nd ed.; E. & S. Livingstone Ltd.: Edinburgh, UK; London, UK, 1962; pp. 58–59. [Google Scholar]
- Rieser, M.J.; Kozlowski, J.F.; Wood, K.V.; McLaughlin, J.L. Muricatacin: A simple biologically active acetogenin derivative from the seeds of Annona muricata (annonaceae). Tetrahedron Lett. 1991, 32, 1137–1140. [Google Scholar] [CrossRef]
- Kim, G.S.; Zeng, L.; Alali, F.; Rogers, L.L.; Wu, F.E.; Sastrodihardjo, S.; McLaughlin, J.L. Muricoreacin and murihexocin C, mono-tetrahydrofuran acetogenins, from the leaves of annona muricata in honour of professor GH Neil Towers 75th birthday. Phytochemistry 1998, 49, 565–571. [Google Scholar] [CrossRef]
- Gbeassor, M.; Kedjagni, A.Y.; Koumaglo, K.; De Souza, C.; Agbo, K.; Aklikokou, K.; Amegbo, K.A. In vitro antimalarial activity of six medicinal plants. Phytother. Res. 1990, 4, 115–117. [Google Scholar] [CrossRef]
- Jaramillo, M.C.; Arango, G.J.; Gonzalez, M.C.; Robledo, S.M.; Velez, I.D. Cytotoxicity and antileishmanial activity of Annona muricata pericarp. Fitoterapia 2000, 71, 183–186. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chang, F.R.; Khalil, A.T.; Hsieh, P.W.; Wu, Y.C. Cytotoxic benzophenanthridine and benzylisoquinoline alkaloids from Argemone mexicana. Z. Naturforsch. C 2003, 58, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.A.; Rahman, F.H.A.; Ibrahim, N.A.; Taha, N.M. Isolation and structure determination of new nematicidal triglyceride from Argemone mexicana. J. Chem. Ecol. 1987, 13, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.L.; Graz, B.; Falquet, J.; Sidibe, O.; Forster, M.; Diallo, D. Argemone mexicana decoction for the treatment of uncomplicated Falciparum malaria. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 1190–1198. [Google Scholar] [CrossRef] [PubMed]
- Graz, B.; Willcox, M.L.; Diakite, C.; Falquet, J.; Dackuo, F.; Sidibe, O.; Giani, S.; Diallo, D. Argemone mexicana decoction versus artesunate-amodiaquine for the management of malaria in Mali: Policy and public-health implications. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.; Sanogo, R.; Diakite, C.; Giani, S.; Paulsen, B.S.; Diallo, D. Improved traditional medicines in Mali. J. Altern. Complement. Med. 2012, 18, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Supratman, U.; Fujita, T.; Akiyama, K.; Hayashi, H.; Murakami, A.; Sakai, H.; Koshimizu, K.; Ohigashi, H. Anti-tumor Promoting Activity of Bufadienolides from Kalanchoe pinnata and K. daigremontiana× butiflora. Biosci. Biotechnol. Biochem. 2001, 65, 947–949. [Google Scholar] [CrossRef] [PubMed]
- Núñez Sellés, A.J.; Vélez Castro, H.T.; Agüero-Agüero, J.; González-González, J.; Naddeo, F.; De Simone, F.; Rastrelli, L. Isolation and quantitative analysis of phenolic antioxidants, free sugars, and polyols from mango (Mangifera indica L.) stem bark aqueous decoction used in Cuba as a nutritional supplement. J. Agric. Food Chem. 2002, 50, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Awe, S.O.; Olajide, O.A.; Oladiran, O.O.; Makinde, J.M. Antiplasmodial and antipyretic screening of Mangifera indica extract. Phytother. Res. 1998, 12, 437–438. [Google Scholar] [CrossRef]
- Kumar, D.S.; Sharathnath, K.V.; Yogeswaran, P.; Harani, A.; Sudhakar, K.; Sudha, P.; Banji, D. A medicinal potency of Momordica charantia. Int. J. Pharm. Sci. Rev. Res. 2010, 1, 95–100. [Google Scholar]
- Elufioye, T.O.; Agbedahunsi, J.M. Antimalarial activities of Tithonia diversifolia (Asteraceae) and Crossopteryx febrifuga (Rubiaceae) on mice in vivo. J. Ethnopharmacol. 2004, 93, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Appiah-Opong, R.; Nyarko, A.K.; Dodoo, D.; Gyang, F.N.; Koram, K.A.; Ayisi, N.K. Antiplasmodial activity of extracts of Tridax procumbens and Phyllanthus amarus in in vitro Plasmodium falciparum culture systems. Ghana Med. J. 2011, 45, 143–150. [Google Scholar] [PubMed]
- Manda, V.K.; Ibrahim, M.A.; Dale, O.R.; Kumarihamy, M.; Cutler, S.J.; Khan, I.A.; Walker, L.A.; Muhammad, I.; Khan, S.I. Modulation of CYPs, P-gp, and PXR by Eschscholzia californica (California poppy) and its alkaloids. Planta Med. 2016, 82, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, U.I.; Büschel, B.; Kirch, W. Unwanted pregnancy on self-medication with St John’s wort despite hormonal contraception. Br. J. Clin. Pharmacol. 2003, 55, 112–113. [Google Scholar] [CrossRef] [PubMed]
- Bent, S.; Goldberg, H.; Padula, A.; Avins, A.L. Spontaneous bleeding associated with Ginkgo biloba. J. Gen. Intern. Med. 2005, 20, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Svensson, U.S.; Ashton, M.; Hai, T.N.; Bertilsson, L.; Houng, D.X.; Houng, N.V.; Nieu, N.T.; Sy, N.D.; Lykkesfeldt, J.; Cong, L.D. Artemisinin induces omeprazole metabolism in human beings. Clin. Pharmacol. Ther. 1998, 64, 160–167. [Google Scholar] [CrossRef]
- Mihara, K.; Svensson, U.S.; Tybring, G.; Hai, T.N.; Bertilsson, L.; Ashton, M. Stereospecific analysis of omeprazole supports artemisinin as a potent inducer of CYP2C19. Fundam. Clin. Pharmacol. 1999, 13, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Simonsson, U.S.; Jansson, B.; Hai, T.N.; Huong, D.X.; Tybring, G.; Ashton, M. Artemisinin autoinduction is caused by involvement of cytochrome P450 2B6 but not 2C9. Clin. Pharmacol. Ther. 2003, 74, 32–43. [Google Scholar] [CrossRef]
- Giao, P.T.; de Vries, P.J. Pharmacokinetic interactions of antimalarial agents. Clin. Pharmacokinet. 2001, 40, 343–373. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Q.; Björkman, A.; Andersson, T.B.; Ridderström, M.; Masimirembwa, C.M. Amodiaquine Clearance and Its Metabolism toN-Desethylamodiaquine Is Mediated by CYP2C8: A New High Affinity and Turnover Enzyme-Specific Probe Substrate. J. Pharmacol. Exp. Ther. 2002, 300, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Rautio, J.; Humphreys, J.E.; Webster, L.O.; Balakrishnan, A.; Keogh, J.P.; Kunta, J.R.; Serabjit-Singh, C.J.; Polli, J.W. In vitro p-glycoprotein inhibition assays for assessment of clinical drug interaction potential of new drug candidates: A recommendation for probe substrates. Drug Metab. Dispos. 2006, 34, 786–792. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.; Einolf, H.J.; Evers, R.; Kumar, G.; Moore, D.; Ripp, S.; Silva, J.; Sinha, V.; Sinz, M.; Skerjanec, A. In vitro and in vivo induction of cytochrome p450: A survey of the current practices and recommendations: A pharmaceutical research and manufacturers of America perspective. Drug Metab. Dispos. 2009, 37, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Mannel, M. Drug interactions with St John’s wort: Mechanisms and clinical implications. Drug Saf. 2004, 27, 773–797. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stanton, J.D.; Tolson, A.H.; Luo, Y.; Wang, H. Bioactive terpenoids and flavonoids from Ginkgo biloba extract induce the expression of hepatic drug-metabolizing enzymes through pregnane X receptor, constitutive androstane receptor, and aryl hydrocarbon receptor mediated pathways. Pharm. Res. 2009, 26, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M. Interaction between food substances and the intestinal epithelium. Biosci. Biotechnol. Biochem. 2010, 74, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Crespi, C.L.; Miller, V.P.; Penman, B.W. Microtiter plate assays for inhibition of human, drug-metabolizing cytochromes P450. Anal. Biochem. 1997, 248, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Manda, V.K.; Avula, B.; Ali, Z.; Wong, Y.H.; Smillie, T.J.; Khan, I.A.; Khan, S.I. Characterization of in vitro ADME properties of diosgenin and dioscin from Dioscorea villosa. Planta Med. 2013, 79, 1421–1428. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Tee, H.W.; Go, M.L. Functionalized chalcones as selective inhibitors of P-glycoprotein and breast cancer resistance protein. Bioorg. Med. Chem. 2008, 16, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Manda, V.K.; Avula, B.; Ali, Z.; Khan, I.A.; Walker, L.A.; Khan, S.I. Evaluation of in vitro ADME properties of mitragynine, 7-hydroxymitragynine, and mitraphylline. Planta Med. 2014, 80, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Manda, V.K.; Dale, O.R.; Awortwe, C.; Ali, Z.; Khan, I.A.; Walker, L.A.; Khan, S.I. Evaluation of drug interaction potential of Labisia pumila (Kacip Fatimah) and its constituents. Front. Pharmacol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the herbal products used in this study are available at the repository of the National Center for Natural Products Research, School of Pharmacy, University of Mississippi, University, MS, USA. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbs (Part Used) | 1A2 | 2A6 | 2B6 | 2C9 | 2C19 | 2D6 | 3A4 |
|---|---|---|---|---|---|---|---|
| IC50 (µg/mL) | |||||||
| Annona muricata (leaves) | 26.1 ± 3.1 | - | >100 | - | 2.1 ± 1.5 | 1.85 ± 3.3 | 2.2 ± 4.4 |
| Argemone Mexicana (whole plant) | >100 | 93.1 ± 8.5 | >100 | - | 98.2 ± 9.3 | 96.1 ± 8.2 | 55.0 ± 5.1 |
| Kalanchoe pinnata (stem) | - | >100 | 96.1 ± 7.7 | - | 91.2 ± 5.8 | - | 55.3 ± 7.2 |
| Mangifera indica (stem bark) | 42.2 ± 4.7 | 47.3 ± 4.1 | 87.4 ± 8.1 | 3.3 ± 0.6 | 32.4 ± 5.1 | >100 | 44.2 ± 3.1 |
| Momordica charantia (leaves and stem) | 23.4 ± 4.3 | 34.1 ± 3.3 | - | 3.3 ± 0.5 | 6.4 ± 1.5 | - | 31.1 ± 5.2 |
| Phyllanthus amarus (leaves, root and stem) | >100 | 28.1 ± 2.6 | 11.2 ± 1.5 | 5.3 ± 1.1 | 3.8 ± 1.0 | - | 0.5 ± 0.2 |
| Tithonia diversifolia (leaves) | 64.3 ± 5.2 | >100 | 98.4 ± 5.5 | 21.2 ± 2.0 | 3.8 ± 4.5 | - | 3.5 ± 2.2 |
| Fold Induction in PXR Activity | |||||
|---|---|---|---|---|---|
| Sample/Conc (µg/mL) | 60 | 20 | 6.67 | 2.22 | 0.74 |
| Annona muricata (leaves) | 1.66 | 1.17 | 1.14 | 0.89 | 0.89 |
| Argemone Mexicana (whole plant) | 2.50 | 2.09 | 1.41 | 1.33 | 1.03 |
| Kalanchoe pinnata (stem) | - | - | - | - | - |
| Mangifera indica (stem bark) | - | - | - | - | - |
| Momordica charantia (leaves and stem) | 4.64 | 2.67 | 1.67 | 1.22 | 1.25 |
| Phyllanthus amarus (leaves, root and stem) | 4.71 | 3.67 | 2.34 | 1.44 | 1.04 |
| Tithonia diversifolia (leaves) | - | 1.47 | 2.41 | 1.74 | 1.35 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fasinu, P.S.; Manda, V.K.; Dale, O.R.; Egiebor, N.O.; Walker, L.A.; Khan, S.I. Modulation of Cytochrome P450, P-glycoprotein and Pregnane X Receptor by Selected Antimalarial Herbs—Implication for Herb-Drug Interaction. Molecules 2017, 22, 2049. https://doi.org/10.3390/molecules22122049
Fasinu PS, Manda VK, Dale OR, Egiebor NO, Walker LA, Khan SI. Modulation of Cytochrome P450, P-glycoprotein and Pregnane X Receptor by Selected Antimalarial Herbs—Implication for Herb-Drug Interaction. Molecules. 2017; 22(12):2049. https://doi.org/10.3390/molecules22122049
Chicago/Turabian StyleFasinu, Pius S., Vamshi K. Manda, Olivia R. Dale, Nosa O. Egiebor, Larry A. Walker, and Shabana I. Khan. 2017. "Modulation of Cytochrome P450, P-glycoprotein and Pregnane X Receptor by Selected Antimalarial Herbs—Implication for Herb-Drug Interaction" Molecules 22, no. 12: 2049. https://doi.org/10.3390/molecules22122049
APA StyleFasinu, P. S., Manda, V. K., Dale, O. R., Egiebor, N. O., Walker, L. A., & Khan, S. I. (2017). Modulation of Cytochrome P450, P-glycoprotein and Pregnane X Receptor by Selected Antimalarial Herbs—Implication for Herb-Drug Interaction. Molecules, 22(12), 2049. https://doi.org/10.3390/molecules22122049