Transcriptomic Analysis of Leaf in Tree Peony Reveals Differentially Expressed Pigments Genes


Abstract
:1. Introduction
2. Results
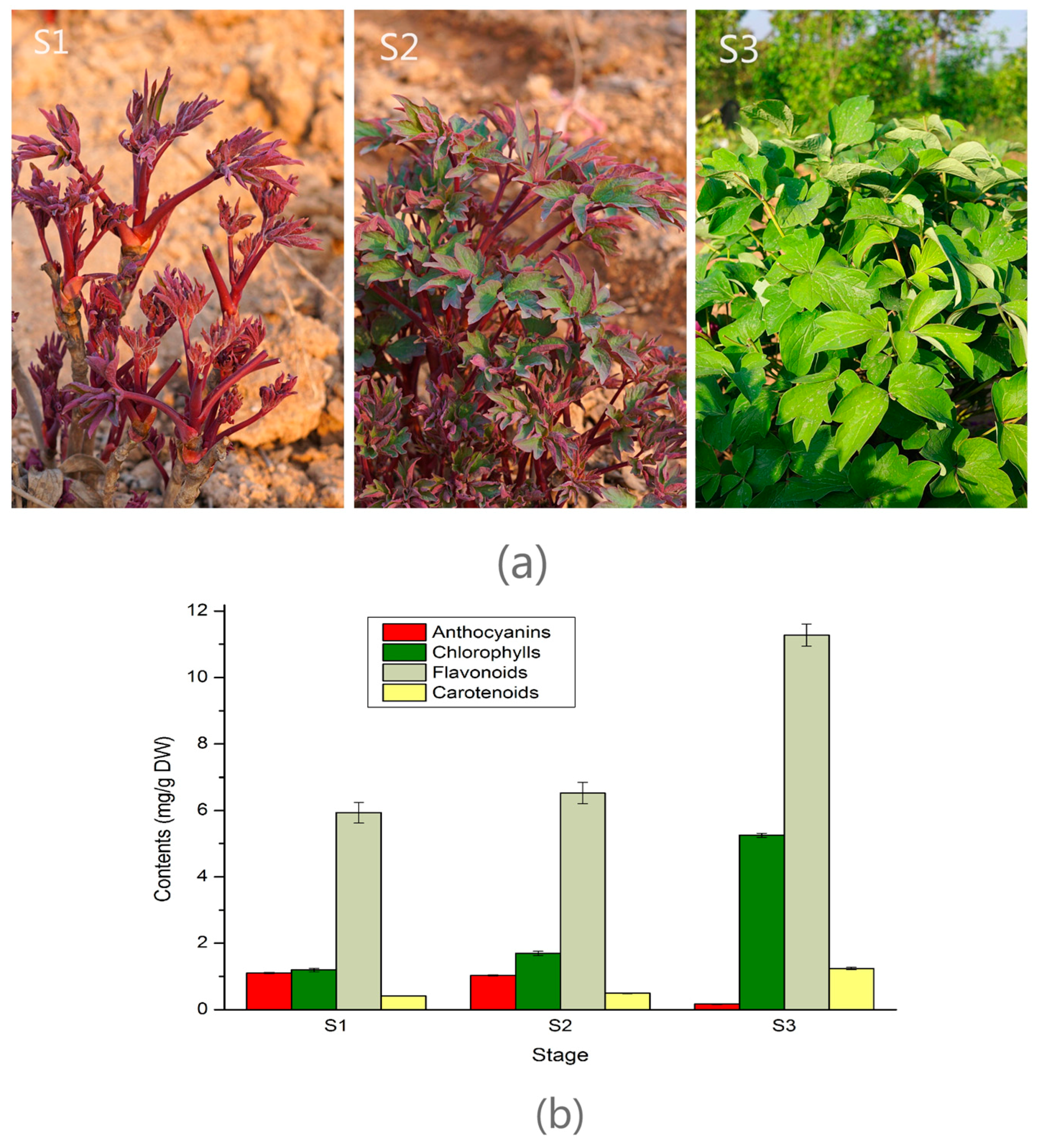
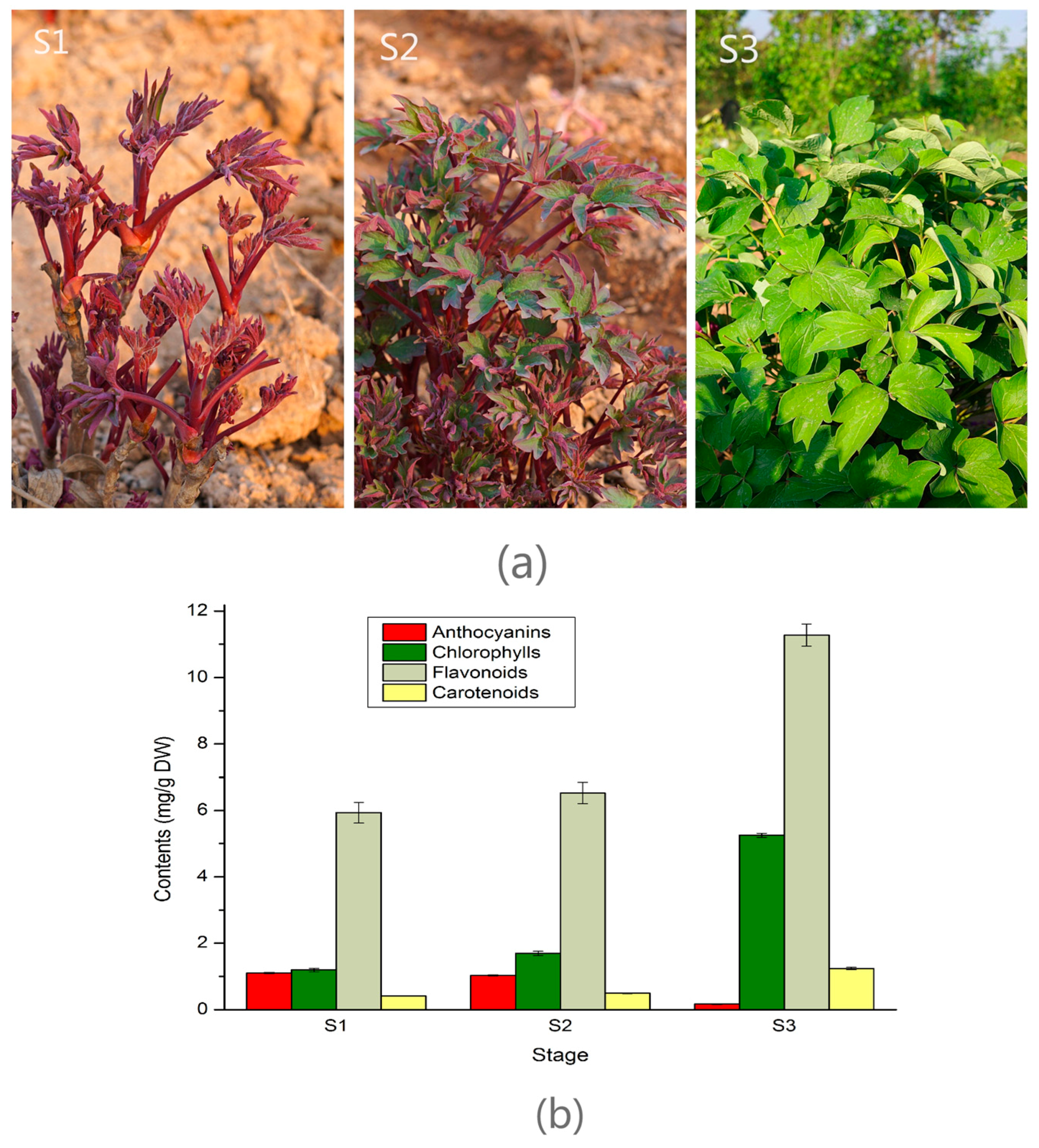
2.1. Pigments Level in the Leaf of Three Developmental Stages in Tree Peony
2.2. Library Construction and Transcriptome Sequencing
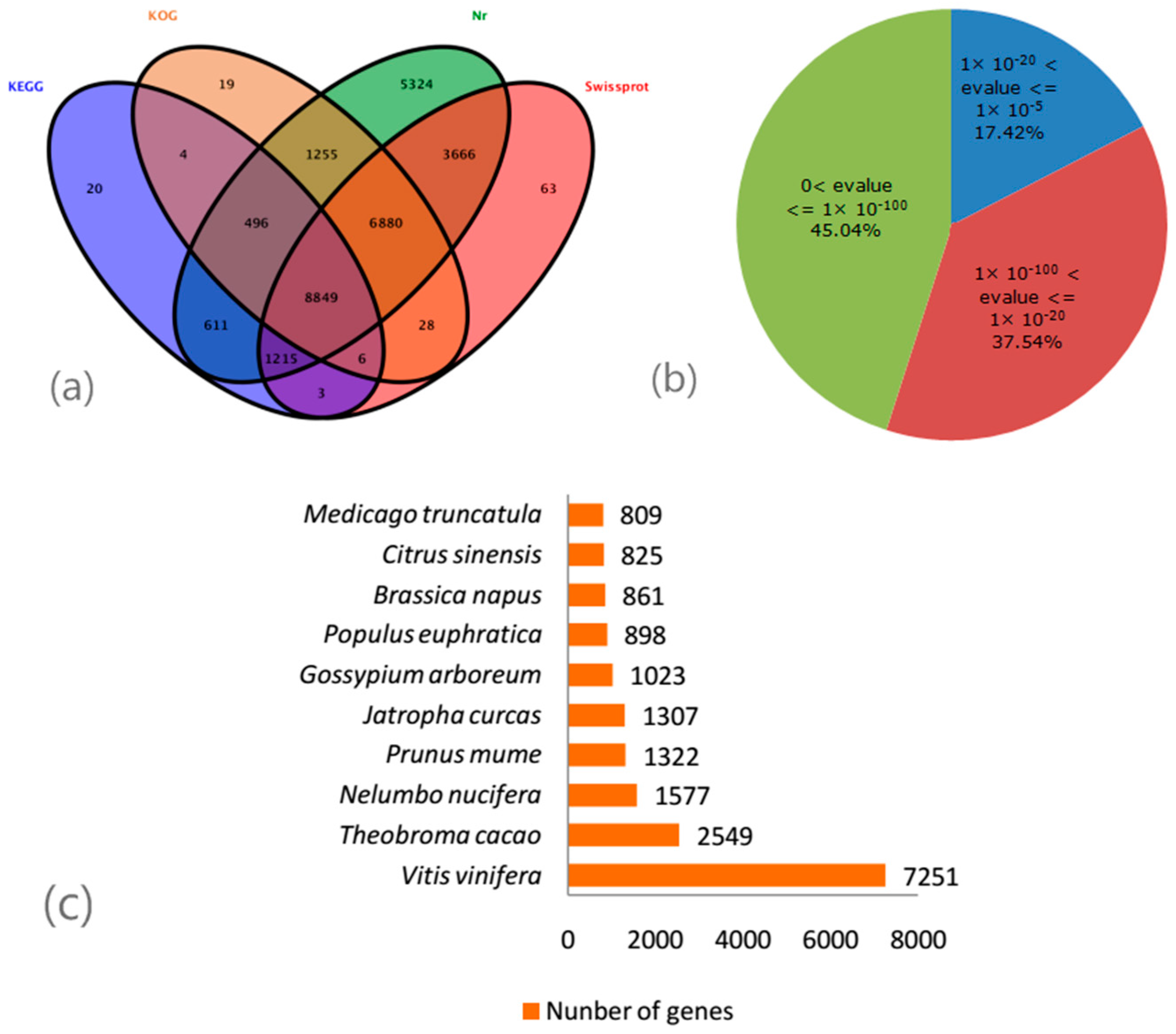
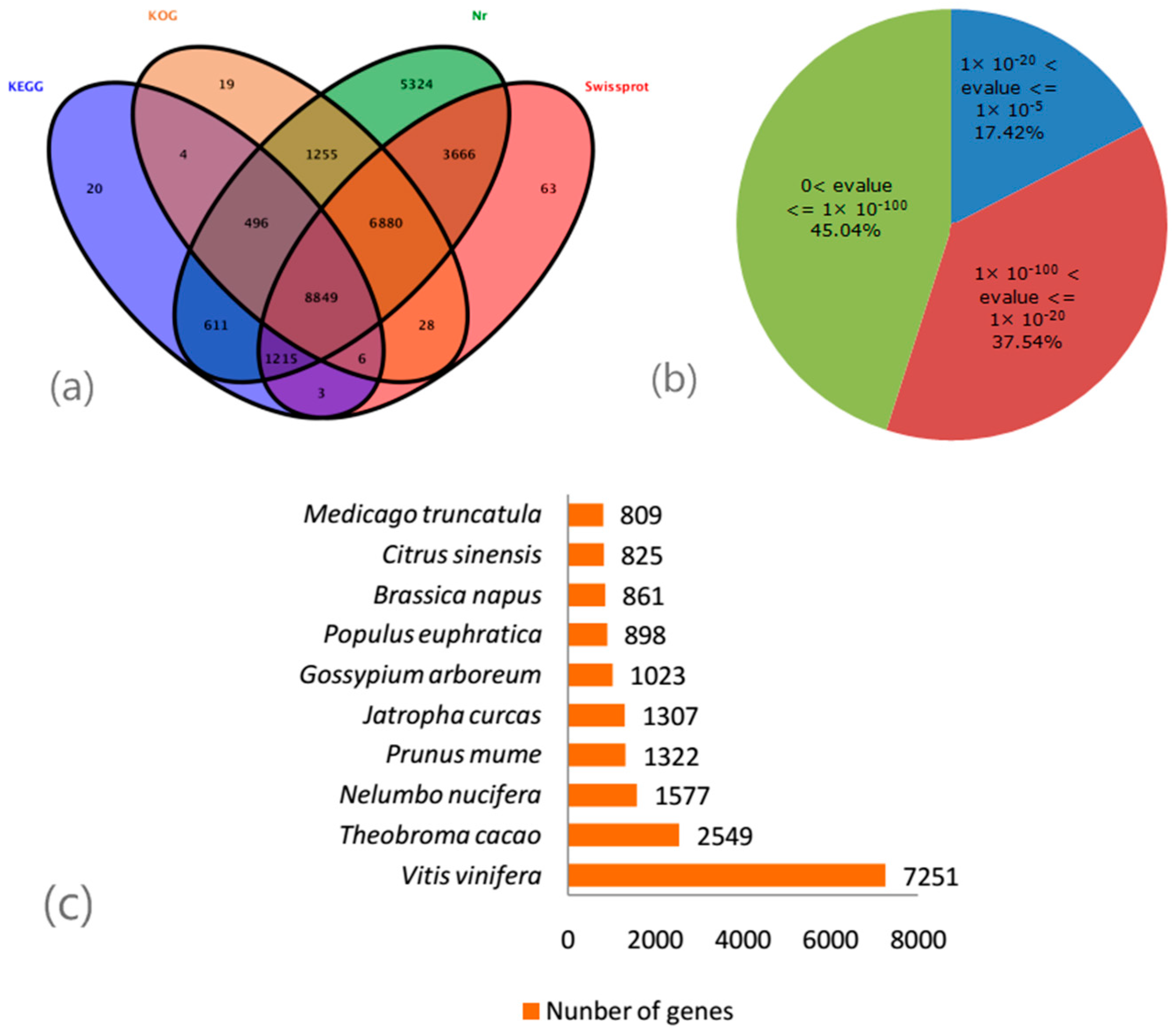
2.3. De Novo Assembly and Gene Annotation
2.4. Identification and Analysis of DEGs between Three Leaf Color Stages
2.5. Annotation of DEGs Probably Involved in Flavonoid/Anthocyanin Biosynthesis
2.6. Annotation of DEGs Probably Involved in Chlorophyll Biosynthesis
2.7. Annotation of DEGs Probably Involved in Carotenoid Biosynthesis
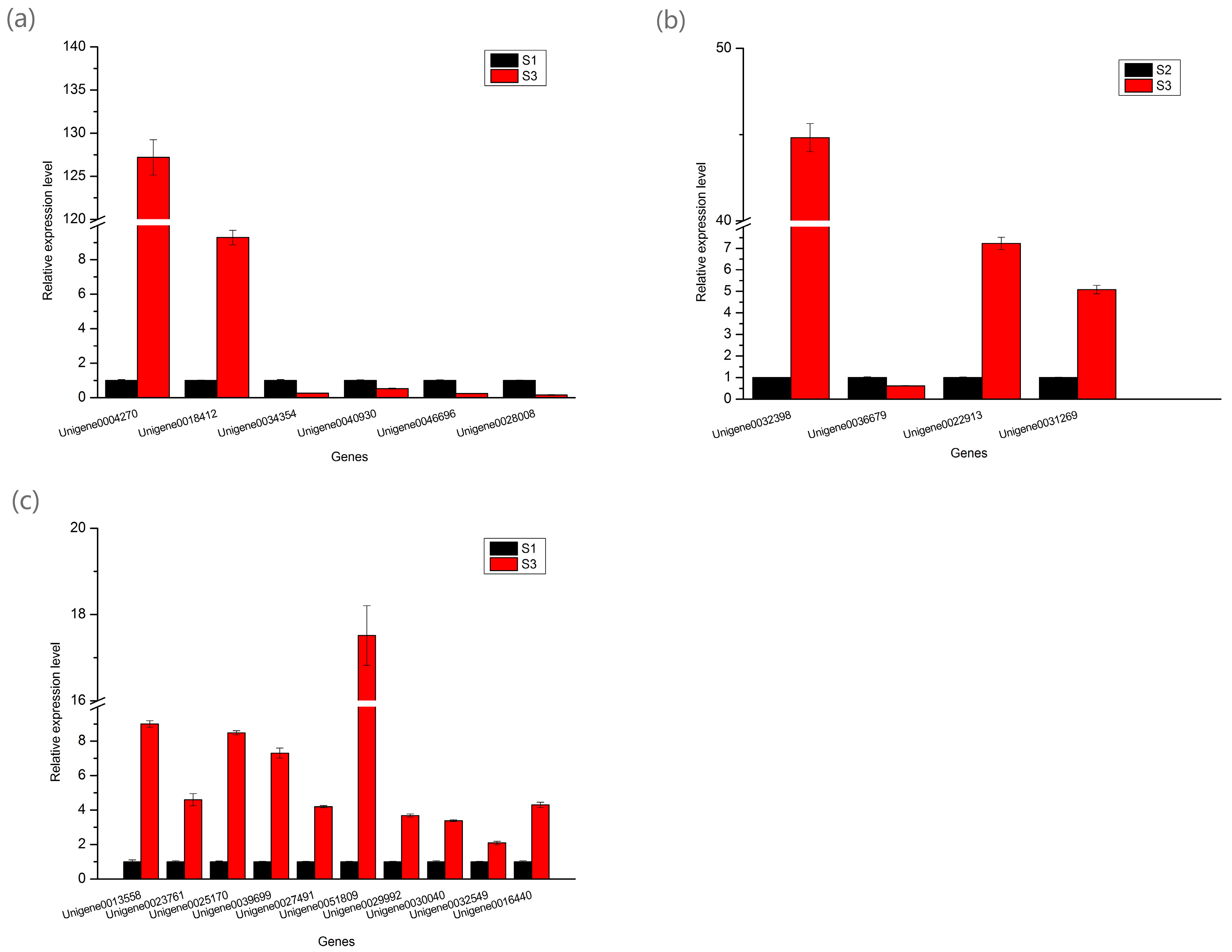
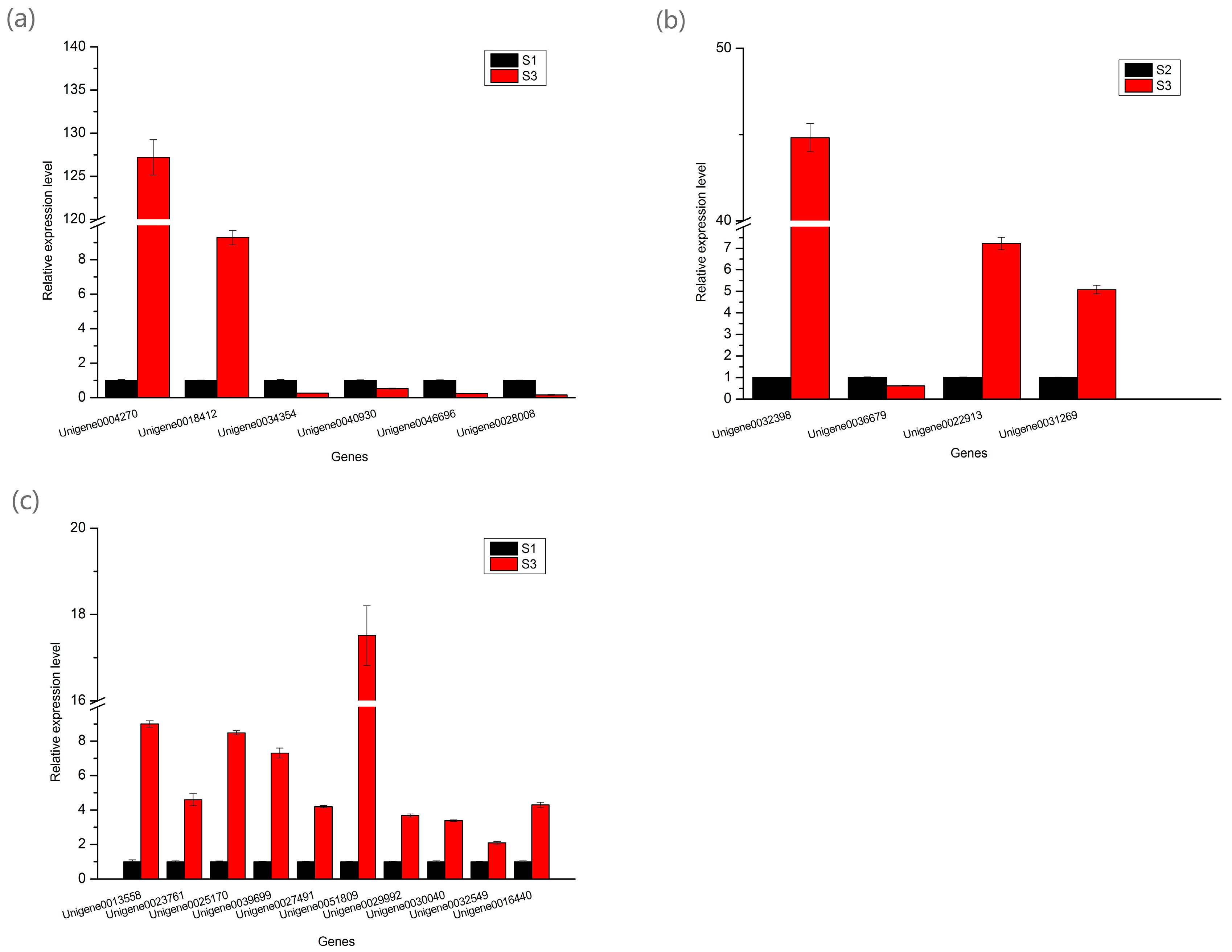
2.8. Real-Time Quantitative PCR Analysis of the DEGs Involved in Pigment Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Measurement of Leaf Pigments
4.2.1. Anthocyanin Level Measurement
4.2.2. Flavonoid Level Measurement
4.2.3. Carotenoid Level Measurement
4.2.4. Chlorophyll Level Measurement
4.3. RNA Extraction, Library Construction, and Transcriptome Sequencing
4.4. De Novo Assembly and Gene Annotation
4.5. Identification and Analysis of DEGs
4.6. Real-Time Quantitative PCR Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ji, L.; Wang, Q.; da Silva, J.A.T.; Yu, X.N. The genetic diversity of Paeonia L. Sci. Hortic. 2012, 143, 62–74. [Google Scholar] [CrossRef]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanin, betalains and carotenoid. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.Q.; Wei, M.R.; Liu, D.; Tao, J. Anatomical and biochemical analysis reveal the role of anthocyanin in flower coloration of herbaceous peony. Plant Physiol. Biochem. 2016, 102, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Li, Q.Z.; Rong, L.P.; Tang, L.; Wang, J.J.; Zhang, B. Analysis of the transcriptome of green and mutant golden-yellow leaves of Acer palmatum Thunb. using high-throughput RNA sequencing. J. Hortic. Sci. Biotechnol. 2015, 90, 388–394. [Google Scholar] [CrossRef]
- Fambrini, M.; Michelotti, V.; Pugliesi, C. Orange, yellow and white-cream: inheritance of carotenoid-based colour in sunflower pollen. Plant Biol. 2010, 12, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, L.; Dai, Y.; Wang, X.; Duan, Y.; Yang, G. De novo transcriptome analysis of Osmanthus serrulatus Rehd. flowers and leaves by Illumina sequencing. Biochem. Syst. Ecol. 2015, 61, 531–540. [Google Scholar] [CrossRef]
- Gao, Z.J.; Han, X.H.; Xiao, X.G. Purification and characterisation of polyphenol oxidase from red Swiss chard (Beta vulgaris subspecies cicla) leaves. Food Chem. 2009, 117, 342–348. [Google Scholar] [CrossRef]
- Wang, C.Q.; Song, H.; Gong, X.Z.; Hu, Q.G.; Liu, F.; Wang, B.-S. Correlation of tyrosinase activity and betacyanin biosynthesis induced by dark in C3 halophyte Suaeda salsa seedlings. Plant Sci. 2007, 173, 487–494. [Google Scholar] [CrossRef]
- Gandia-Herrero, F.; Garcia-Carmona, F. Biosynthesis of betalains: yellow and violet plant pigments. Trends Plant Sci. 2013, 18, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Shiraishi, A.; Hashimoto, F.; Aoki, N.; Shimizu, K.; Sakata, Y. Analysis of petal anthocyanin to investigate flower coloration of Zhongyuan (Chinese) and Daikon Island (Japanese) tree peony cultivars. J. Plant Res. 2001, 114, 33–43. [Google Scholar] [CrossRef]
- Wang, L.S.; Hashimoto, F.; Shiraishi, A.; Aoki, N.; Li, J.J.; Sakata, Y. Chemical taxonomy of the Xibei tree peony from China by floral pigmentation. J. Plant Res. 2004, 117, 47–55. [Google Scholar] [PubMed]
- Zhang, J.; Wang, L.; Shu, Q.; Liu, Z.A.; Li, C.; Zhang, J.; Wei, X.; Tian, D. Comparison of anthocyanin in non-blotches and blotches of the petals of Xibei tree peony. Sci. Hortic. 2007, 114, 104–111. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, W.; Wang, Y.; Gao, S.; Du, D.; Fu, J.; Dong, L. Anthocyanin biosynthesis and accumulation in developing flowers of tree peony (Paeonia suffruticosa) ‘Luoyang Hong’. Postharvest Biol. Technol. 2014, 97, 11–22. [Google Scholar] [CrossRef]
- Zhang, Y.; Cheng, Y.; Ya, H.; Xu, S.; Han, J. Transcriptome sequencing of purple petal spot region in tree peony reveals differentially expressed anthocyanin structural genes. Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Wang, Y.; Ren, L.; Shi, Q.; Zheng, B.; Miao, K.; Guo, X. Overexpression of Ps-CHI1, a homologue of the chalcone isomerase gene from tree peony (Paeonia suffruticosa), reduces the intensity of flower pigmentation in transgenic tobacco. Plant Cell Tiss. Org. 2013, 116, 285–295. [Google Scholar] [CrossRef]
- Shi, Q.; Zhou, L.; Wang, Y.; Li, K.; Zheng, B.; Miao, K. Transcriptomic analysis of Paeonia delavayi wild population flowers to identify differentially expressed genes involved in purple-red and yellow petal pigmentation. PLoS ONE 2015, 10, e0135038. [Google Scholar] [CrossRef] [PubMed]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.W.; Sagawa, J.M.; Frost, L.; Vela, J.P.; Bradshaw, H.D., Jr. Transcriptional control of floral anthocyanin pigmentation in monkey flowers (Mimulus). New Phytol. 2014, 204, 1013–1027. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Chen, Y.Y.; Tsai, W.C.; Chen, W.H.; Chen, H.H. Three R2R3-MYB transcription factors regulate distinct floral pigmentation patterning in Phalaenopsis spp. Plant Physiol. 2015, 168, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Kriangphan, N.; Vuttipongchaikij, S.; Kittiwongwattana, C.; Suttangkakul, A.; Pinmanee, P.; Sakulsathaporn, A.; Suwimon, R.; Suputtitada, S.; Chanvivattana, Y.; Apisitwanich, S. Effects of sequence and expression of eight anthocyanin biosynthesis genes on floral coloration in four Dendrobium hybrids. Hortic. J. 2015, 84, 83–92. [Google Scholar] [CrossRef]
- Tanaka, Y.; Ohmiya, A. Seeing is believing: engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotechnol. 2008, 19, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, X.; Chen, W.; Dong, M.; Yuan, W.; Liu, X.; Shang, F. Differential expression of carotenoid-related genes determines diversified carotenoid coloration in flower petal of Osmanthus fragrans. Tree Genet. Genomes 2013, 10, 329–338. [Google Scholar] [CrossRef]
- Saito, T.; Honma, D.; Tagashira, M.; Kanda, T.; Nesumi, A.; Maeda-Yamamoto, M. Anthocyanin from new red leaf tea “Sunrouge”. J. Agric. Food Chem. 2011, 59, 4779–4782. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Mu, H.; Jiang, J.; Liu, G. Transcriptomic analysis of purple leaf determination in birch. Gene 2013, 526, 251–258. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhang, Z.; Zhang, L. Anthocyanin accumulation, antioxidant ability and stability, and a transcriptional analysis of anthocyanin biosynthesis in purple heading Chinese cabbage (Brassica rapa L. ssp. pekinensis). J. Agric. Food Chem. 2016, 64, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xu, M.; Luo, Q.; Wang, J.; Li, H. De novo transcriptome analysis of Liriodendron chinense petals and leaves by Illumina sequencing. Gene 2014, 534, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lai, B.; Hu, B.; Qin, Y.H.; Zhao, J.T.; Wang, H.C.; Hu, G.B. Transcriptomic analysis of Litchi chinensis pericarp during maturation with a focus on chlorophyll degradation and flavonoid biosynthesis. BMC Genom. 2015, 16, 225. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Huang, X.M.; Hu, G.B.; Yang, Z.Y.; Huang, H.-B. A comparative study of chlorophyll loss and its related mechanism during fruit maturation in the pericarp of fast- and slow-degreening litchi pericarp. Sci. Hortic. 2005, 106, 247–257. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J.; Han, C.; Ge, J. Flower color diversity revealed by differential expression of flavonoid biosynthetic genes and flavonoid accumulation in herbaceous peony (Paeonia lactiflora Pall.). Mol. Biol. Rep. 2012, 39, 11263–11275. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Li, L.; Cheng, S.; Cao, F.; Wang, Y.; Yuan, H. Molecular cloning and function assay of a chalcone isomerase gene (GbCHI) from Ginkgo biloba. Plant Cell Rep. 2011, 30, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Jin, Z.; Qu, W.; Zhao, D.; Ma, F. Cloning of a cDNA encoding the Saussurea medusa chalcone isomerase and its expression in transgenic tobacco. Plant Physiol. Biochem. 2006, 44, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Huang, C.; Gong, Y.; Feng, Q.; Gao, F. Molecular Cloning and Expression Analysis of an ANS gene encoding anthocyanidin synthase from purple-fleshed sweet potato [Ipomoea batatas (L.) Lam]. Plant Mol. Biol. Rep. 2009, 28, 112–121. [Google Scholar] [CrossRef]
- Chen, S.M.; Li, C.H.; Zhu, X.R.; Deng, Y.M.; Sun, W.; Wang, L.S.; Chen, F.D.; Zhang, Z. The identification of flavonoids and the expression of genes of anthocyanin biosynthesis in the chrysanthemum. Biol. Plant. 2012, 56, 458–464. [Google Scholar] [CrossRef]
- Zhang, J.; Han, Z.Y.; Tian, J.; Zhang, X.; Song, T.T.; Yao, Y.C. The expression level of anthocyanidin synthase determines the anthocyanin content of crabapple (Malus sp.) petals. Acta Physiol. Plant. 2015, 37, 1–10. [Google Scholar] [CrossRef]
- Szankowski, I.; Flachowsky, H.; Li, H.; Halbwirth, H.; Treutter, D.; Regos, I.; Hanke, M.V.; Stich, K.; Fischer, T.C. Shift in polyphenol profile and sublethal phenotype caused by silencing of anthocyanidin synthase in apple (Malus sp.). Planta 2009, 229, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Beale, S.I. Green genes gleaned. Trends Plant Sci. 2005, 10, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Hortensteiner, S. Update on the biochemistry of chlorophyll breakdown. Plant Mol. Biol. 2013, 82, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Dong, L.; Zhang, C.; Wang, X.Q. Reference gene selection for real-time quantitative PCR normalization in tree peony (Paeonia suffruticosa Andr.). J. Agric. Biotechnol. 2012, 20, 521–528. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GeneID | log2 Ratio (S3/S1) a | FDR | Annotation |
|---|---|---|---|
| Unigene0004270 | 6.88 | 1.17 × 10−9 | Chalcone isomerase |
| Unigene0018412 | 2.90 | 0.007807 | Shikimate hydroxycinnamoyltransferase |
| Unigene0034354 | −1.49 | 0.000353 | Flavanone 3-hydroxylase |
| Unigene0040930 | −1.58 | 0.000112 | Flavonoid 3′-hydroxylase |
| Unigene0046696 | −1.65 | 0.000548 | Dihydroflavonol 4-reductase |
| Unigene0028008 | −3.40 | 6.79 × 10−12 | Anthocyanidin synthase |
| Unigene0036137 | −1.06 | 0.023175 | UDP-glycosyltransferase |
| Unigene0052871 | 1.90 | 0.001475 | Glucoside glucosyltransferase |
| GeneID | log2 Ratio (S3/S1) a | FDR | Annotation |
|---|---|---|---|
| Unigene0006988 | −10.28 | 0.002581722 | Transcription factor MYB39 [Aegilops tauschii] |
| Unigene0028536 | −4.40 | 3.34 × 10−12 | MYB-like protein X-like [Citrus sinensis] |
| Unigene0033848 | −3.43 | 3.58 × 10−6 | MYB domain protein 106 [Theobroma cacao] |
| Unigene0019117 | −3.15 | 5.67 × 10−5 | MYB domain protein 17 isoform 1 [Theobroma cacao] |
| Unigene0015077 | −2.98 | 1.59 × 10−8 | MYB-related protein 3R-1-like isoform X1 [Vitis vinifera] |
| Unigene0025015 | −2.92 | 3.02 × 10−7 | Transcription factor MYB90 [Vitis vinifera] |
| Unigene0015076 | −2.77 | 3.04 × 10−7 | MYB transcription factor family[Populus trichocarpa] |
| Unigene0017724 | −2.73 | 0.008752272 | MYB-related protein 3R-1-like isoform X1 [Vitis vinifera] |
| Unigene0023351 | −2.67 | 0.005438748 | MYB-related protein A-like [Populus euphratica] |
| Unigene0003954 | −2.57 | 0.007298376 | MYB-related protein 3R-1-like isoform X1 [Vitis vinifera] |
| Unigene0040613 | −2.57 | 0.000168857 | MYB family transcription factor APL isoform X2 [Vitis vinifera] |
| Unigene0031865 | −2.41 | 3.11 × 10−7 | MYB transcription factor family[Populus trichocarpa] |
| Unigene0023350 | −2.09 | 0.006013724 | MYB-related protein 3R-1-like [Prunus mume] |
| Unigene0003975 | −1.60 | 0.000354814 | Transcription factor MYB1R1-like [Citrus sinensis] |
| Unigene0031901 | −1.19 | 0.032182662 | MYB DNA-binding domain protein [Theobroma cacao] |
| Unigene0017529 | 1.10 | 0.010140177 | MYB transcription factor family [Populus trichocarpa] |
| Unigene0022785 | 1.32 | 0.041043118 | Transcription factor MYB44-like [Vitis vinifera] |
| Unigene0030933 | 1.48 | 0.01321785 | MYB-related protein 308-like [Populus euphratica] |
| Unigene0003846 | 1.57 | 0.02700583 | MYB transcription factor family [Populus trichocarpa] |
| Unigene0003228 | 2.00 | 0.009708022 | MYB-like protein J [Malus domestica] |
| Unigene0021304 | 2.21 | 0.028027338 | Transcription factor MYB59 [Vitis vinifera] |
| Unigene0009655 | 2.25 | 0.000336502 | MYB transcription factor family [Populus trichocarpa] |
| Unigene0018495 | 2.27 | 0.000636996 | Transcription factor MYB82 [Jatropha curcas] |
| Unigene0016379 | 2.49 | 0.000248643 | MYB family transcription factor APL [Prunus mume] |
| Unigene0013255 | 2.87 | 0.002952733 | R2R3 MYBA6 transcription factor splice variant 1 [Vitis vinifera] |
| Unigene0018284 | 3.07 | 0.002052337 | MYB family transcription factor At1g14600 [Vitis vinifera] |
| Unigene0029454 | 3.13 | 6.71 × 10−5 | MYB-like transcription factor [Betula luminifera] |
| Unigene0017856 | −6.92 | 2.88 × 10−20 | Transcription factor bHLH94-like [Nelumbo nucifera] |
| Unigene0011325 | −3.98 | 0.001209742 | bHLH family protein [Populus trichocarpa] |
| Unigene0011327 | −3.79 | 0.000339255 | bHLH family protein [Populus trichocarpa] |
| Unigene0001264 | −3.48 | 0.000243391 | bHLH family protein [Populus trichocarpa] |
| Unigene0023425 | −3.26 | 6.47 × 10−5 | Transcription factor bHLH118-like [Jatropha curcas] |
| Unigene0013501 | −2.60 | 0.033069525 | Transcription factor bHLH93 [Vitis vinifera] |
| Unigene0026438 | −2.44 | 0.000225777 | Transcription factor bHLH70 isoform X2 [Vitis vinifera] |
| Unigene0030075 | −2.32 | 0.000257132 | Transcription factor bHLH30-like [Jatropha curcas] |
| Unigene0031959 | −2.14 | 0.000135867 | Transcription factor bHLH66-like isoform X4 [Vitis vinifera] |
| Unigene0041894 | −1.94 | 5.05 × 10−5 | Transcription factor bHLH35-like [Pyrus x bretschneideri] |
| Unigene0041893 | −1.90 | 0.00691905 | Transcription factor bHLH35-like [Pyrus x bretschneideri] |
| Unigene0033453 | −1.66 | 0.000163627 | Myc anthocyanin regulatory protein isoform X1 [Vitis vinifera] |
| Unigene0009061 | −1.60 | 0.011318569 | Transcription factor bHLH104-like isoform X1 [Citrus sinensis] |
| Unigene0000218 | −1.28 | 0.026379909 | Transcription factor bHLH49 isoform X1 [Vitis vinifera] |
| Unigene0037994 | −1.22 | 0.03284377 | Transcription factor bHLH145-like [Prunus mume] |
| Unigene0033727 | 1.06 | 0.015085073 | Transcription factor bHLH63 [Vitis vinifera] |
| Unigene0014388 | 1.18 | 0.03179475 | Transcription factor bHLH51 [Vitis vinifera] |
| Unigene0020369 | 1.79 | 0.00037921 | Transcription factor bHLH149 [Solanum lycopersicum] |
| Unigene0023437 | 1.81 | 0.00014609 | Transcription factor bHLH63 [Prunus mume] |
| Unigene0027343 | 1.88 | 0.000282396 | Transcription factor bHLH61 isoform 2 [Theobroma cacao] |
| Unigene0032390 | −5.15 | 6.23 × 10−17 | F-box/WD repeat-containing 10 [Gossypium arboreum] |
| Unigene0019877 | −4.98 | 1.75 × 10−6 | WD repeat-containing 62 [Gossypium arboreum] |
| Unigene0052972 | −3.88 | 8.77 × 10−8 | WD repeat-containing protein 76 isoform X2 [Jatropha curcas] |
| Unigene0017625 | −3.76 | 0.000361154 | F-box/WD repeat-containing protein sel-10 [Jatropha curcas] |
| Unigene0038354 | −3.59 | 1.16 × 10−14 | WD repeat-containing protein 76 isoform X1 [Jatropha curcas] |
| Unigene0038352 | −3.35 | 6.74 × 10−8 | WD repeat-containing protein 76 isoform X2 [Jatropha curcas] |
| Unigene0038982 | −3.33 | 5.71 × 10−10 | WD repeat and HMG-box DNA-binding protein 1 [Vitis vinifera] |
| Unigene0004606 | −3.25 | 1.42 × 10−6 | WD repeat-containing protein 76 isoform X1 [Jatropha curcas] |
| Unigene0016452 | −2.38 | 2.55 × 10−6 | WD-40 repeat-containing protein MSI3 [Vitis vinifera] |
| Unigene0038353 | −2.35 | 0.000579262 | WD repeat-containing protein 76 [Gossypium raimondii] |
| Unigene0038355 | −2.28 | 0.004815745 | WD repeat-containing protein 76 isoform X2 [Jatropha curcas] |
| Unigene0033696 | −2.24 | 6.56 × 10−7 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0033696 | −2.24 | 6.56 × 10−7 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0031235 | −2.22 | 1.05 × 10−8 | WD-40 repeat-containing protein MSI4-like [Eucalyptus grandis] |
| Unigene0017354 | −2.05 | 0.000604273 | WD repeat-containing protein 5-like [Nelumbo nucifera] |
| Unigene0041973 | −1.94 | 4.57 × 10−5 | WD repeat-containing protein 43 isoform X1 [Vitis vinifera] |
| Unigene0011781 | −1.86 | 0.003509861 | Glutamate-rich WD repeat-containing protein 1 [Nelumbo nucifera] |
| Unigene0046703 | −1.71 | 0.023069191 | WD repeat-containing 6 [Gossypium arboreum] |
| Unigene0041702 | −1.68 | 0.002347404 | WD repeat-containing protein 3 [Vitis vinifera] |
| Unigene0018836 | −1.63 | 0.007290383 | WD repeat-containing protein 82 [Jatropha curcas] |
| Unigene0024647 | −1.61 | 2.61 × 10−5 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0028464 | −1.47 | 0.007517559 | WD repeat-containing protein [Gossypium arboreum] |
| Unigene0041538 | −1.39 | 0.0043636 | WD repeat-containing protein 87-like [Cicer arietinum] |
| Unigene0030123 | −1.34 | 0.001589388 | WD-40 repeat-containing protein MSI1 [Vitis vinifera] |
| Unigene0051975 | −1.32 | 0.002923971 | WD repeat-containing protein 61 [Nelumbo nucifera] |
| Unigene0031526 | −1.18 | 0.018270378 | Katanin p80 WD40 repeat-containing subunit B1 homolog isoform X2 [Vitis vinifera] |
| Unigene0039012 | −1.13 | 0.010137359 | WD repeat-containing protein 70 [Vitis vinifera] |
| Unigene0032745 | −1.12 | 0.014302649 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0045126 | −1.08 | 0.031686906 | WD40 protein [Paeonia suffruticosa] |
| Unigene0049600 | 1.24 | 0.010794516 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0019158 | 1.62 | 0.014228592 | WD repeat-containing protein LWD1-like [Gossypium raimondii] |
| Unigene0029541 | 1.84 | 0.000911988 | WD repeat-containing 70 [Gossypium arboreum] |
| Unigene0015065 | 2.21 | 3.62 × 10−6 | F-box/WD repeat-containing protein [Medicago truncatula] |
| Unigene0051483 | 2.58 | 0.008905399 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0033466 | 3.91 | 5.28 × 10−6 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| Unigene0033465 | 5.08 | 1.24 × 10−5 | Transducin/WD40 repeat-like superfamily protein [Theobroma cacao] |
| GeneID | log2 Ratio (S3/S2) a | FDR | Annotation |
|---|---|---|---|
| Unigene0031269 | 1.94 | 0.03382 | Mg-protoporphyrinogen IX monomethylester |
| Unigene0022913 | 1.02 | 0.047351 | Divinyl reductase |
| Unigene0036679 | 1.03 | 0.033455 | Chlorophyllide a oxygenase |
| Unigene0045551 | −1.42 | 0.009836 | Glutamyl-tRNA synthetase |
| Unigene0032398 | 1.09 | 0.02502 | Protochlorophyllide oxidoreductase |
| Unigene0039869 | −5.47 | 6.00 × 10−16 | Geranylgeranyl reductase |
| Unigene0047139 | 1.40 | 0.003807 | Pheophorbide a oxygenase |
| Unigene0026448 | 1.32 | 0.019098 | Protoheme ferro-lyase |
| GeneID | log2 Ratio (S3/S1) a | FDR | Annotation |
|---|---|---|---|
| Unigene0029992 | 2.07 | 2.10 × 10−7 | Phytoene synthase 2, chloroplastic-like |
| Unigene0030040 | 1.77 | 3.96 × 10−5 | Phytoene desaturase, partial |
| Unigene0025170 | 2.28 | 4.78 × 10−5 | Beta-carotene isomerase D27 |
| Unigene0039699 | 1.71 | 3.42 × 10−5 | Zeta-carotene desaturase |
| Unigene0013558 | 1.99 | 3.48 × 10−7 | Violaxanthin de-epoxidase, chloroplastic |
| Unigene0023761 | 1.08 | 0.014272 | Lycopene beta cyclase, chloroplastic |
| Unigene0016440 | 1.38 | 0.000987 | Lycopene epsilon cyclase |
| Unigene0032549 | 2.55 | 0.013492 | Beta-carotene hydroxylase, partial |
| Unigene0027491 | 1.60 | 5.36 × 10−5 | Zeaxanthin epoxidase, partial |
| Unigene0051809 | 2.88 | 2.71 × 10−13 | Carotenoid cleavage dioxygenase |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Shi, Q.; Niu, L.; Zhang, Y. Transcriptomic Analysis of Leaf in Tree Peony Reveals Differentially Expressed Pigments Genes. Molecules 2017, 22, 324. https://doi.org/10.3390/molecules22020324
Luo J, Shi Q, Niu L, Zhang Y. Transcriptomic Analysis of Leaf in Tree Peony Reveals Differentially Expressed Pigments Genes. Molecules. 2017; 22(2):324. https://doi.org/10.3390/molecules22020324
Chicago/Turabian StyleLuo, Jianrang, Qianqian Shi, Lixin Niu, and Yanlong Zhang. 2017. "Transcriptomic Analysis of Leaf in Tree Peony Reveals Differentially Expressed Pigments Genes" Molecules 22, no. 2: 324. https://doi.org/10.3390/molecules22020324
APA StyleLuo, J., Shi, Q., Niu, L., & Zhang, Y. (2017). Transcriptomic Analysis of Leaf in Tree Peony Reveals Differentially Expressed Pigments Genes. Molecules, 22(2), 324. https://doi.org/10.3390/molecules22020324