
Black Tea High-Molecular-Weight Polyphenol-Rich Fraction Promotes Hypertrophy during Functional Overload in Mice


Abstract
:1. Introduction
2. Results
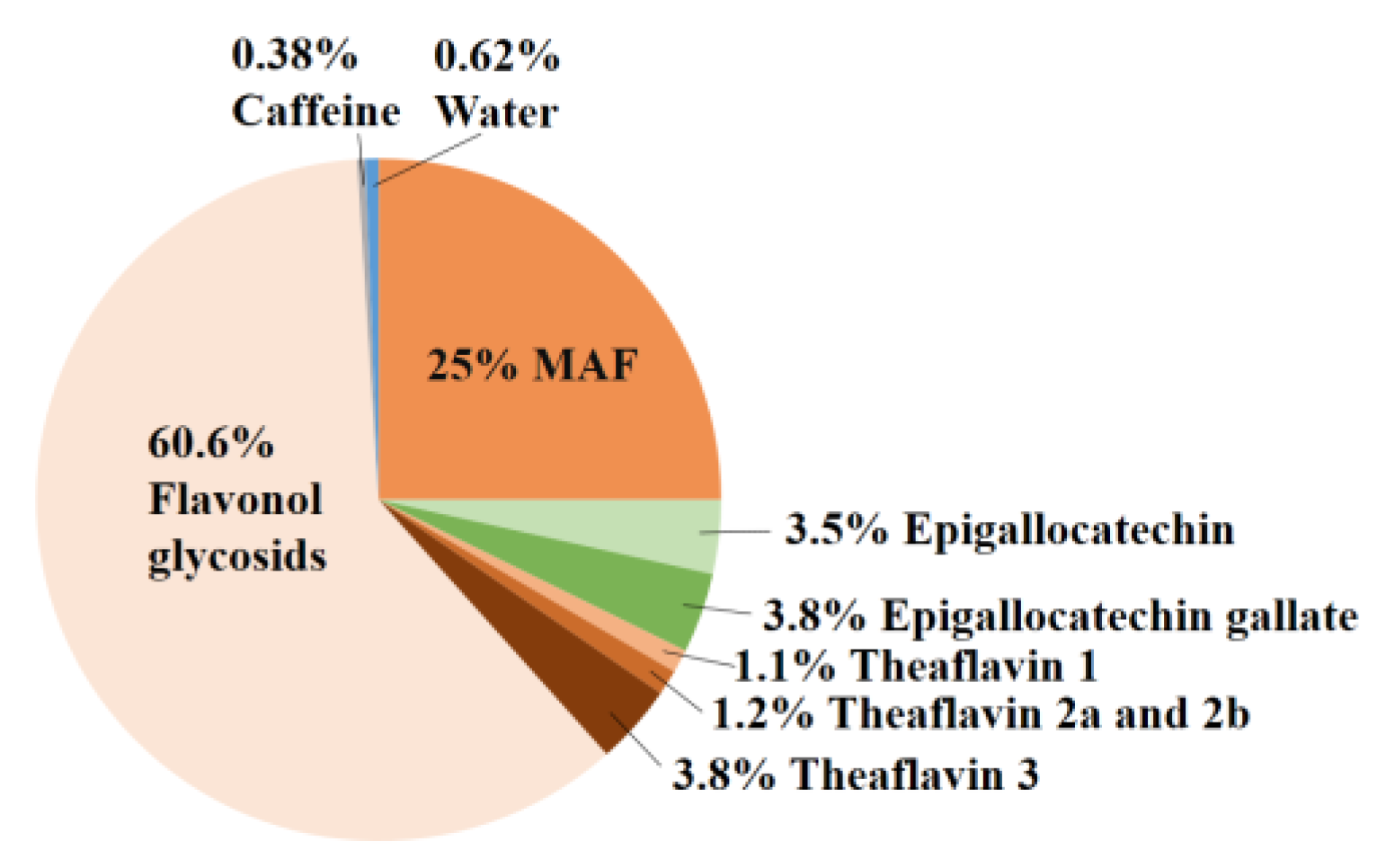
2.1. Composition of E80
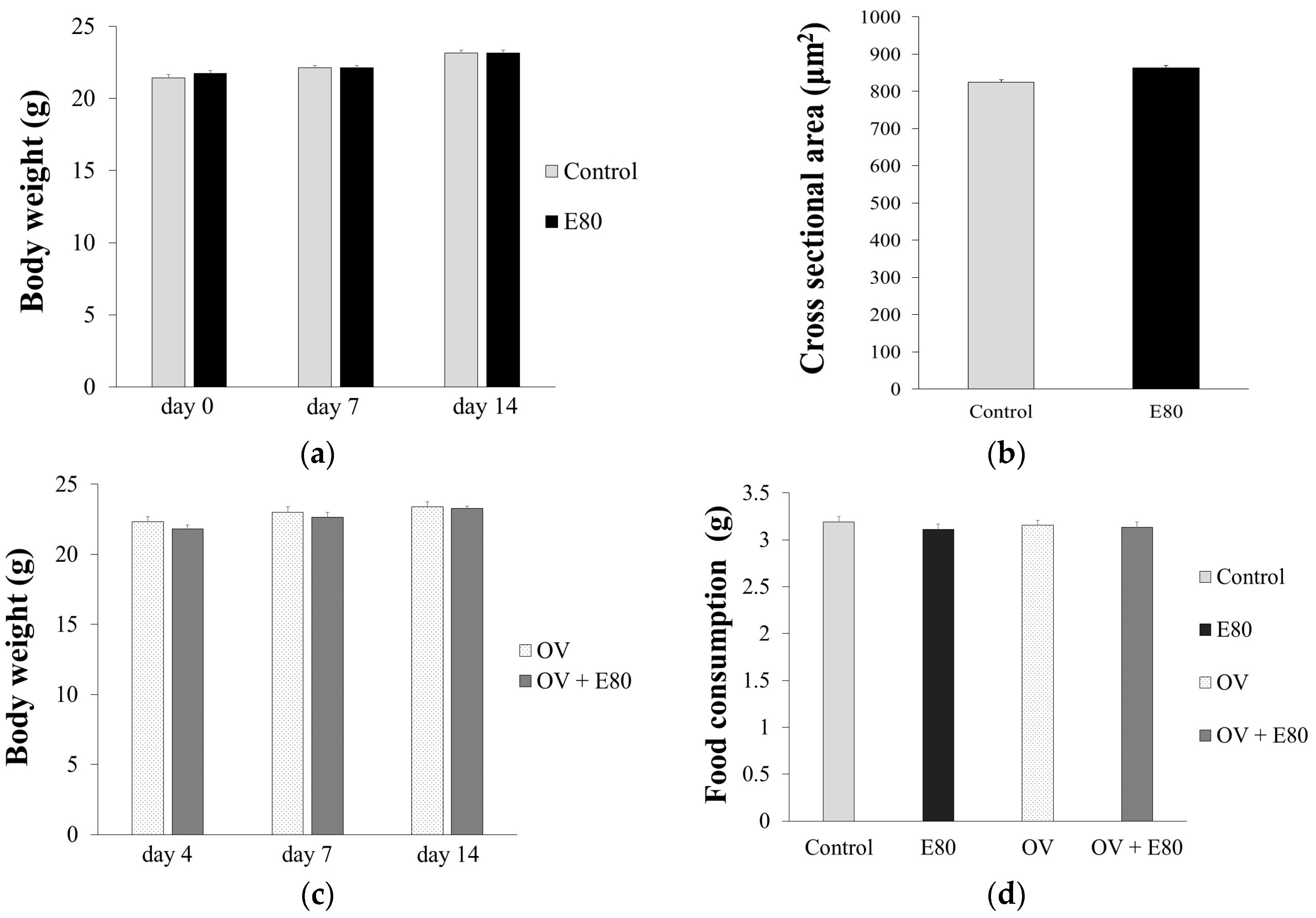
2.2. E80 Has No Effect on Food Intake and Growth
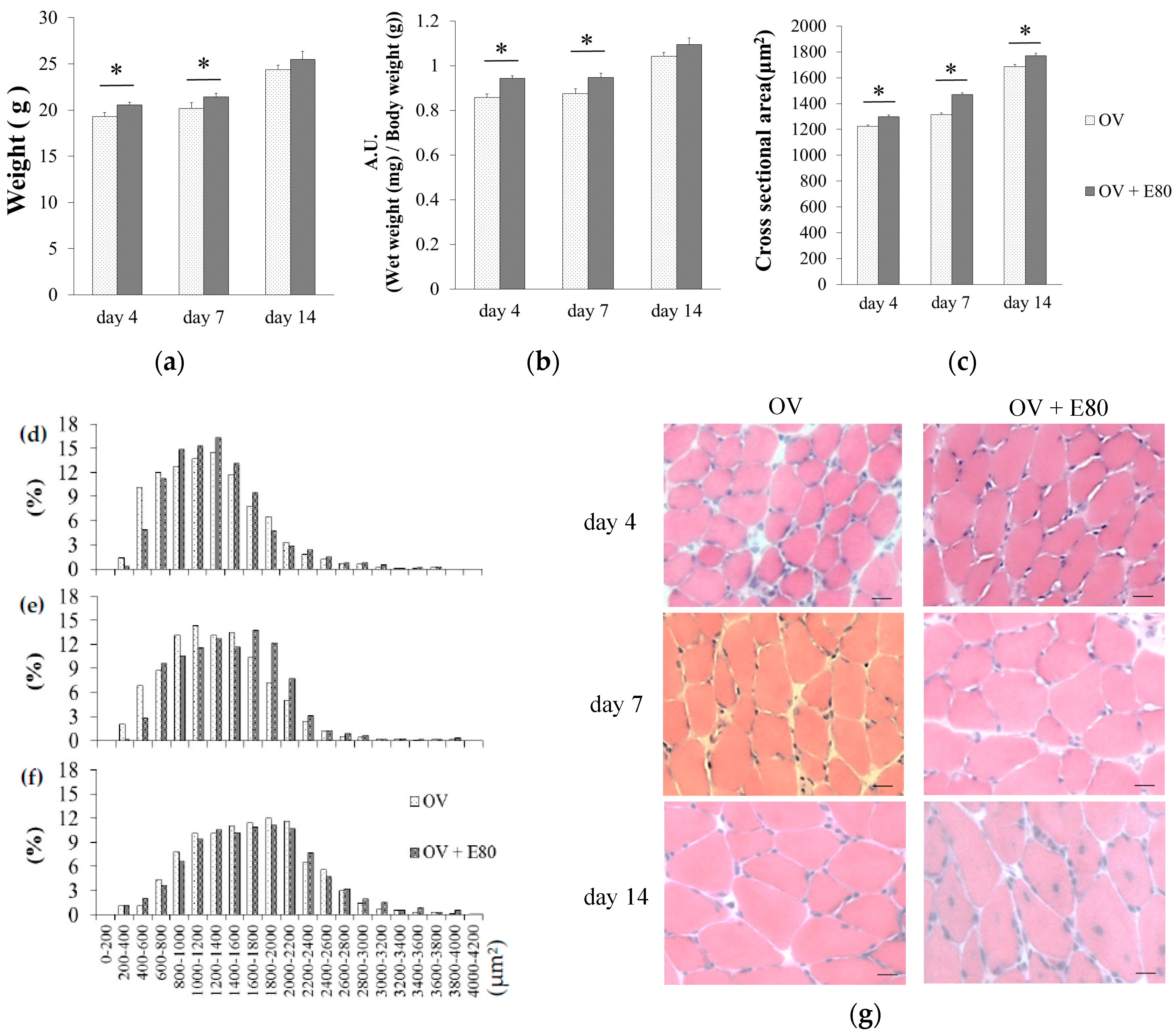
2.3. E80 Improves Overload-Induced Muscle Hypertrophy
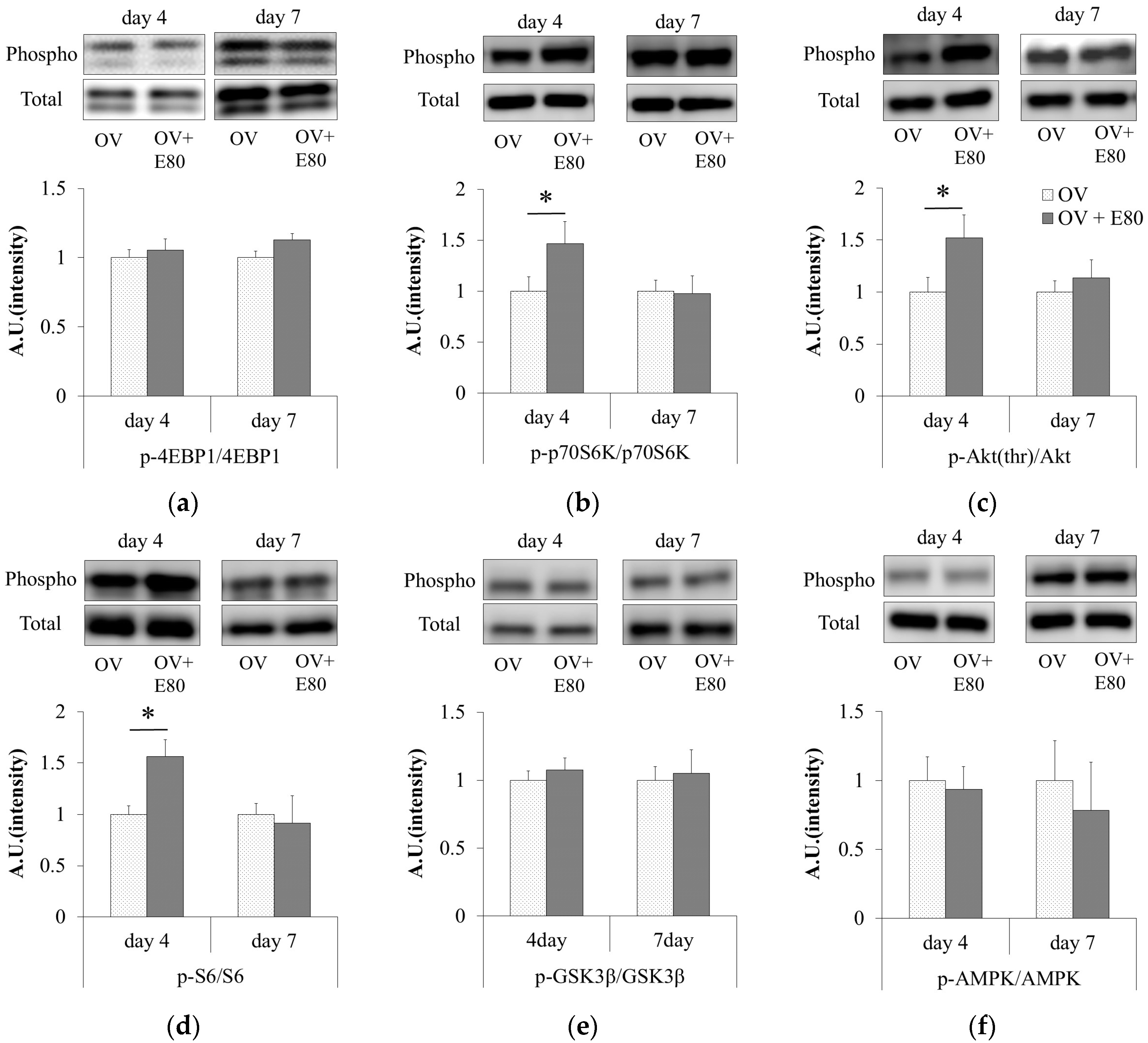
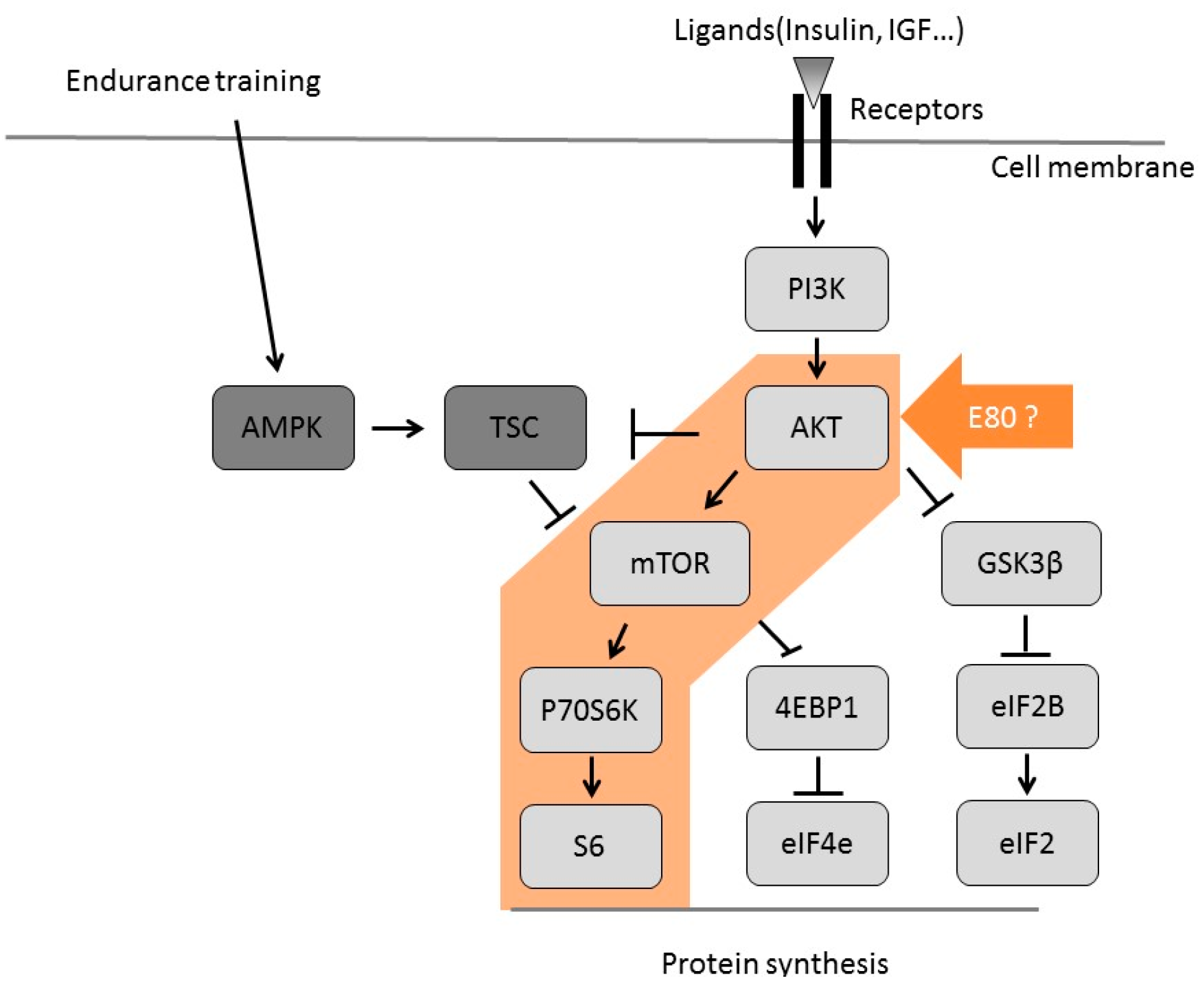
2.4. E80 Intake Activates the Akt/mTOR Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Mice and Functional Overload
4.2. Preparation of MAF-Rich Sample (E80) from Black Tea
4.3. Quantitative Analysis of Caffeine, Catechins, and Theaflavins in E80
4.4. Quantitative Analysis of Highly Polymerized Polyphenols in E80
4.5. Preparing Diet
4.6. Antibodies
4.7. Cross-Sectional Area Quantification
4.8. Western Blot Analysis
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 4EBP1 | Eukaryotic translation initiation factor 4E-binding protein 1 |
| AMPK | 5′ adenosine monophosphate-activated protein kinase |
| ANOVA | Analysis of variance |
| CSA | Cross sectional area |
| eIF2 | Eukaryotic Initiation Factor 2 |
| eIF2B | Eukaryotic Initiation Factor 2B |
| eIF4E | Eukaryotic Initiation Factor 4E |
| GSK3β | Glycogen synthase kinase 3 beta |
| HPLC | High performance liquid chromatography |
| IGF | Insulin-like growth factors |
| MAF | Mitochondria activation factor |
| mTOR | Mammalian target of rapamycin |
| p70S6K | P70 ribosomal protein S6 kinase |
| PGC1-α | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| PI3K | Phosphoinositide 3-kinase |
| PVDF | Polyvinylidene Difluoride |
| S6 | Ribosomal protein S6 kinase |
| TSC | Tuberous sclerosis complex |
References
- Weisburger, J.H. Tea and health: A historical perspective. Cancer Lett. 1997, 114, 315–317. [Google Scholar] [CrossRef]
- Toschi, T.G.; Bordoni, A.; Hrelia, S.; Bendini, A.; Lercker, G.; Biagi, P.L. The protective role of different green tea extracts after oxidative damage is related to their catechin composition. J. Agric. Food Chem. 2000, 48, 3973–3978. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, Y.; Xie, Z.; Zhou, Y.; Zhang, Y.; Wan, X. The anti-obesity effects of green tea in human intervention and basic molecular studies. Eur. J. Clin. Nutr. 2014, 68, 1075–1087. [Google Scholar] [PubMed]
- Han, L.K.; Takaku, T.; Li, J.; Kimura, Y.; Okuda, H. Anti-obesity action of oolong tea. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Murase, T.; Nagasawa, A.; Suzuki, J.; Hase, T.; Tokimitsu, I. Beneficial effects of tea catechins on diet-induced obesity: Stimulation of lipid catabolism in the liver. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Sae-Tan, S.; Grove, K.A.; Kennett, M.J.; Lambert, J.D. (−)-epigallocatechin-3-gallate increases the expression of genes related to fat oxidation in the skeletal muscle of high fat-fed mice. Food Funct. 2011, 2, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Alway, S.E.; Bennett, B.T.; Wilson, J.C.; Edens, N.K.; Pereira, S.L. Epigallocatechin-3-gallate improves plantaris muscle recovery after disuse in aged rats. Exp. Gerontol. 2014, 50, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Meador, B.M.; Mirza, K.A.; Tian, M.; Skelding, M.B.; Reaves, L.A.; Edens, N.K.; Tisdale, M.J.; Pereira, S.L. The green tea polyphenol epigallocatechin-3-gallate (egcg) attenuates skeletal muscle atrophy in a rat model of sarcopenia. J. Frailty Aging 2015, 4, 209–215. [Google Scholar] [PubMed]
- Gupta, S.; Hastak, K.; Ahmad, N.; Lewin, J.S.; Mukhtar, H. Inhibition of prostate carcinogenesis in tramp mice by oral infusion of green tea polyphenols. Proc. Natl. Acad Sci. USA 2001, 98, 10350–10355. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Tajima, K.; Mizutani, M.; Iwata, H.; Iwase, T.; Miura, S.; Hirose, K.; Hamajima, N.; Tominaga, S. Regular consumption of green tea and the risk of breast cancer recurrence: Follow-up study from the hospital-based epidemiologic research program at aichi cancer center (herpacc), Japan. Cancer Lett. 2001, 167, 175–182. [Google Scholar] [CrossRef]
- Stewart, C.E.; Rittweger, J. Adaptive processes in skeletal muscle: Molecular regulators and genetic influences. J. Musculoskelet. Neuronal Interact. 2006, 6, 73–86. [Google Scholar] [PubMed]
- Pette, D.; Staron, R.S. Myosin isoforms, muscle fiber types, and transitions. Microsc. Res. Tech. 2000, 50, 500–509. [Google Scholar] [CrossRef]
- Hamilton, M.T.; Booth, F.W. Skeletal muscle adaptation to exercise: A century of progress. J. Appl. Physiol. (1985) 2000, 88, 327–331. [Google Scholar]
- Kirkendall, D.T.; Garrett, W.E., Jr. The effects of aging and training on skeletal muscle. Am. J. Sports Med. 1998, 26, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, T.; Ursula, S.P.R.; Stephano, M.F.S.; Cleber, A.R.; Edilamaar, O.M. Signaling pathways that mediate skeletal muscle hypertrophy: Effects of exercise training. INTECH Open Access Publ. 2012. [Google Scholar] [CrossRef]
- Fujihara, T.; Nakagawa-Izumi, A.; Ozawa, T.; Numata, O. High-molecular-weight polyphenols from oolong tea and black tea: Purification, some properties, and role in increasing mitochondrial membrane potential. Biosci. Biotechnol. Biochem. 2007, 71, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, T.; Kumagai, C.; Fujihara, T.; Takemasa, T.; Ozawa, T.; Numata, O. Black tea high-molecular-weight polyphenol stimulates exercise training-induced improvement of endurance capacity in mouse via the link between ampk and glut4. PLoS ONE 2013, 8, e69480. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, C.; Tagesson, C. Influence of organic solvent mixtures on biological membranes. Br. J. Ind. Med. 1985, 42, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Campana, C.; Griffin, P.L.; Simon, E.L. Caffeine overdose resulting in severe rhabdomyolysis and acute renal failure. Am. J. Emerg. Med. 2014, 32, 111.e3–111.e4. [Google Scholar] [PubMed]
- Baar, K.; Esser, K. Phosphorylation of p70(s6k) correlates with increased skeletal muscle mass following resistance exercise. Am. J. Physiol. 1999, 276, C120–C127. [Google Scholar] [PubMed]
- Terzis, G.; Spengos, K.; Mascher, H.; Georgiadis, G.; Manta, P.; Blomstrand, E. The degree of p70 s6k and s6 phosphorylation in human skeletal muscle in response to resistance exercise depends on the training volume. Eur. J. Appl. Physiol. 2010, 110, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Jager, S.; Handschin, C.; St-Pierre, J.; Spiegelman, B.M. Amp-activated protein kinase (ampk) action in skeletal muscle via direct phosphorylation of pgc-1alpha. Proc. Natl. Acad Sci. USA 2007, 104, 12017–12022. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.K.; Mukai, K.; Lally, J.S.; Maher, A.C.; Gurd, B.J.; Heigenhauser, G.J.; Spriet, L.L.; Holloway, G.P. Amp-activated protein kinase is required for exercise-induced peroxisome proliferator-activated receptor co-activator 1 translocation to subsarcolemmal mitochondria in skeletal muscle. J. Physiol. 2013, 591, 1551–1561. [Google Scholar] [PubMed]
- Lantier, L.; Fentz, J.; Mounier, R.; Leclerc, J.; Treebak, J.T.; Pehmoller, C.; Sanz, N.; Sakakibara, I.; Saint-Amand, E.; Rimbaud, S.; et al. Ampk controls exercise endurance, mitochondrial oxidative capacity, and skeletal muscle integrity. FASEB J. 2014, 28, 3211–3224. [Google Scholar] [CrossRef] [PubMed]
- Lee-Young, R.S.; Griffee, S.R.; Lynes, S.E.; Bracy, D.P.; Ayala, J.E.; McGuinness, O.P.; Wasserman, D.H. Skeletal muscle amp-activated protein kinase is essential for the metabolic response to exercise in vivo. J. Biol. Chem. 2009, 284, 23925–23934. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Zhu, T.; Guan, K.L. Tsc2 mediates cellular energy response to control cell growth and survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef]
- Gwinn, D.M.; Shackelford, D.B.; Egan, D.F.; Mihaylova, M.M.; Mery, A.; Vasquez, D.S.; Turk, B.E.; Shaw, R.J. Ampk phosphorylation of raptor mediates a metabolic checkpoint. Mol. Cell 2008, 30, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Mounier, R.; Lantier, L.; Leclerc, J.; Sotiropoulos, A.; Foretz, M.; Viollet, B. Antagonistic control of muscle cell size by ampk and mtorc1. Cell Cycle 2011, 10, 2640–2646. [Google Scholar] [CrossRef] [PubMed]
- Mounier, R.; Lantier, L.; Leclerc, J.; Sotiropoulos, A.; Pende, M.; Daegelen, D.; Sakamoto, K.; Foretz, M.; Viollet, B. Important role for ampkalpha1 in limiting skeletal muscle cell hypertrophy. FASEB J. 2009, 23, 2264–2273. [Google Scholar] [PubMed]
- McGee, S.L.; Mustard, K.J.; Hardie, D.G.; Baar, K. Normal hypertrophy accompanied by phosphoryation and activation of amp-activated protein kinase alpha1 following overload in lkb1 knockout mice. J. Physiol. 2008, 586, 1731–1741. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Selby, A.; Rankin, D.; Patel, R.; Atherton, P.; Hildebrandt, W.; Williams, J.; Smith, K.; Seynnes, O.; Hiscock, N.; et al. Age-related differences in the dose-response relationship of muscle protein synthesis to resistance exercise in young and old men. J. Physiol. 2009, 587, 211–217. [Google Scholar] [PubMed]
- Evans, W.J. Skeletal muscle loss: Cachexia, sarcopenia, and inactivity. Am. J. Clin. Nutr. 2010, 91, 1123S–1127S. [Google Scholar] [CrossRef] [PubMed]
- Fielding, R.A.; Vellas, B.; Evans, W.J.; Bhasin, S.; Morley, J.E.; Newman, A.B.; Abellan van Kan, G.; Andrieu, S.; Bauer, J.; Breuille, D.; et al. Sarcopenia: An undiagnosed condition in older adults. Current consensus definition: Prevalence, etiology, and consequences. International working group on sarcopenia. J. Am. Med. Dir. Assoc. 2011, 12, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Spangenburg, E.E.; Booth, F.W. Leukemia inhibitory factor restores the hypertrophic response to increased loading in the lif(-/-) mouse. Cytokine 2006, 34, 125–130. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) | Food Consumption (g) | CSA (µm2) | |||
|---|---|---|---|---|---|
| day 4 | day 7 | day 14 | |||
| Control | 21.44 ± 0.36 | 22.13 ± 0.4 | 23.17 ± 0.37 | 3.19 ± 0.06 | 824.75 ± 5.44 |
| E80 | 21.74 ± 0.31 | 22.14 ± 0.34 | 23.19 ± 0.15 | 3.11 ± 0.06 | 862.83 ± 6.23 |
| OV | 22.33 ± 0.38 | 22.99 ± 0.34 | 23.4 ± 0.36 | 3.15 ± 0.06 | |
| OV + E80 | 21.8 ± 0.33 | 22.64 ± 0.35 | 23.27 ± 0.32 | 3.13 ± 0.06 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aoki, Y.; Ozawa, T.; Takemasa, T.; Numata, O. Black Tea High-Molecular-Weight Polyphenol-Rich Fraction Promotes Hypertrophy during Functional Overload in Mice. Molecules 2017, 22, 548. https://doi.org/10.3390/molecules22040548
Aoki Y, Ozawa T, Takemasa T, Numata O. Black Tea High-Molecular-Weight Polyphenol-Rich Fraction Promotes Hypertrophy during Functional Overload in Mice. Molecules. 2017; 22(4):548. https://doi.org/10.3390/molecules22040548
Chicago/Turabian StyleAoki, Yuki, Tetsuo Ozawa, Tohru Takemasa, and Osamu Numata. 2017. "Black Tea High-Molecular-Weight Polyphenol-Rich Fraction Promotes Hypertrophy during Functional Overload in Mice" Molecules 22, no. 4: 548. https://doi.org/10.3390/molecules22040548
APA StyleAoki, Y., Ozawa, T., Takemasa, T., & Numata, O. (2017). Black Tea High-Molecular-Weight Polyphenol-Rich Fraction Promotes Hypertrophy during Functional Overload in Mice. Molecules, 22(4), 548. https://doi.org/10.3390/molecules22040548