
Screening, Purification and Characterization of Anionic Antimicrobial Proteins from Foeniculum Vulgare



Abstract
:1. Introduction
2. Results and Discussion
2.1. Comparison of Protein Concentration Extracted in Different Buffers after Dialysis
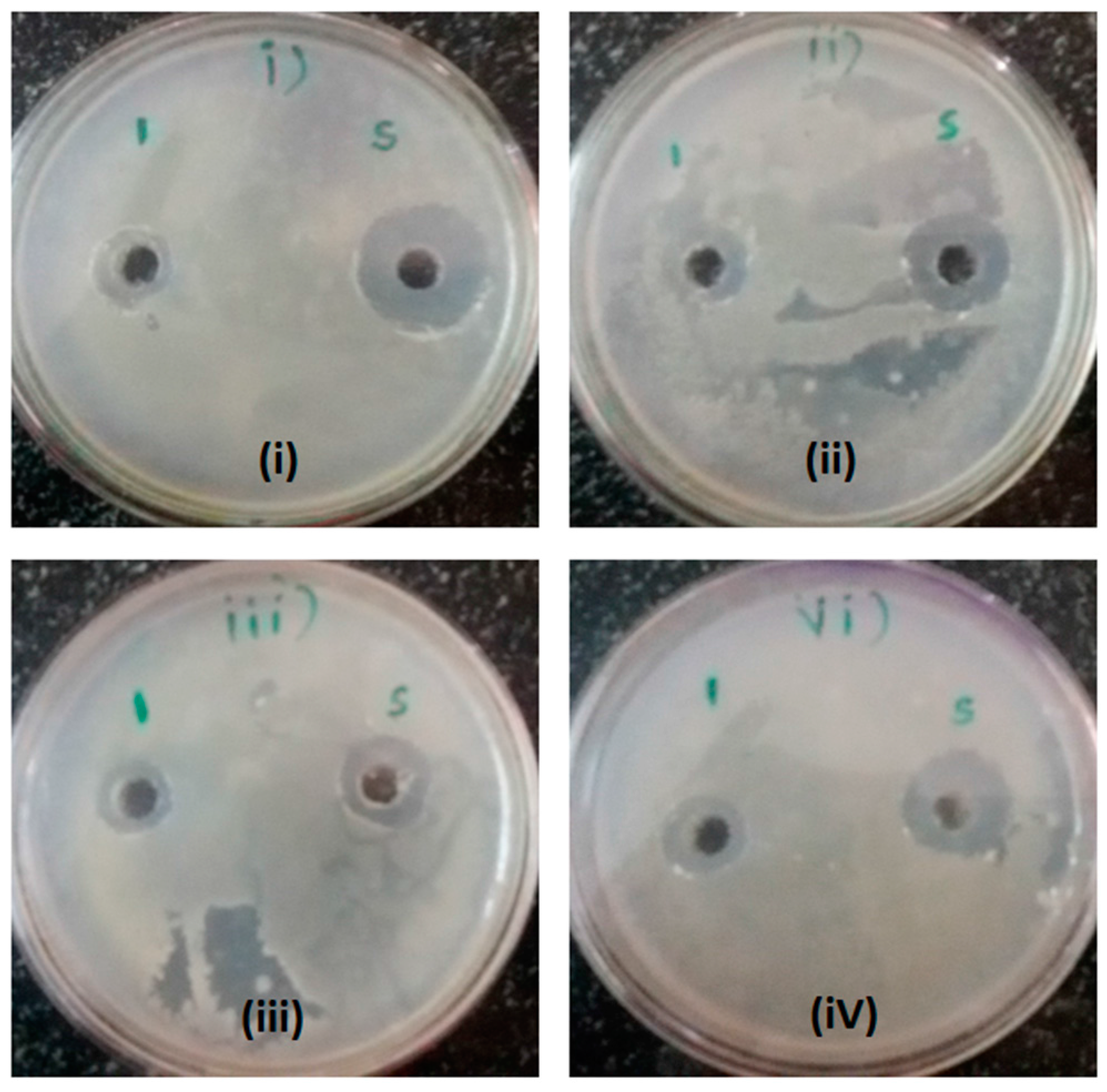
2.2. Antibacterial Activity of Sodium Acetate Buffer pH-6.0 Extracts after Dialysis
2.3. Protein Concentration of Ion Exchange Chromatography Elutes
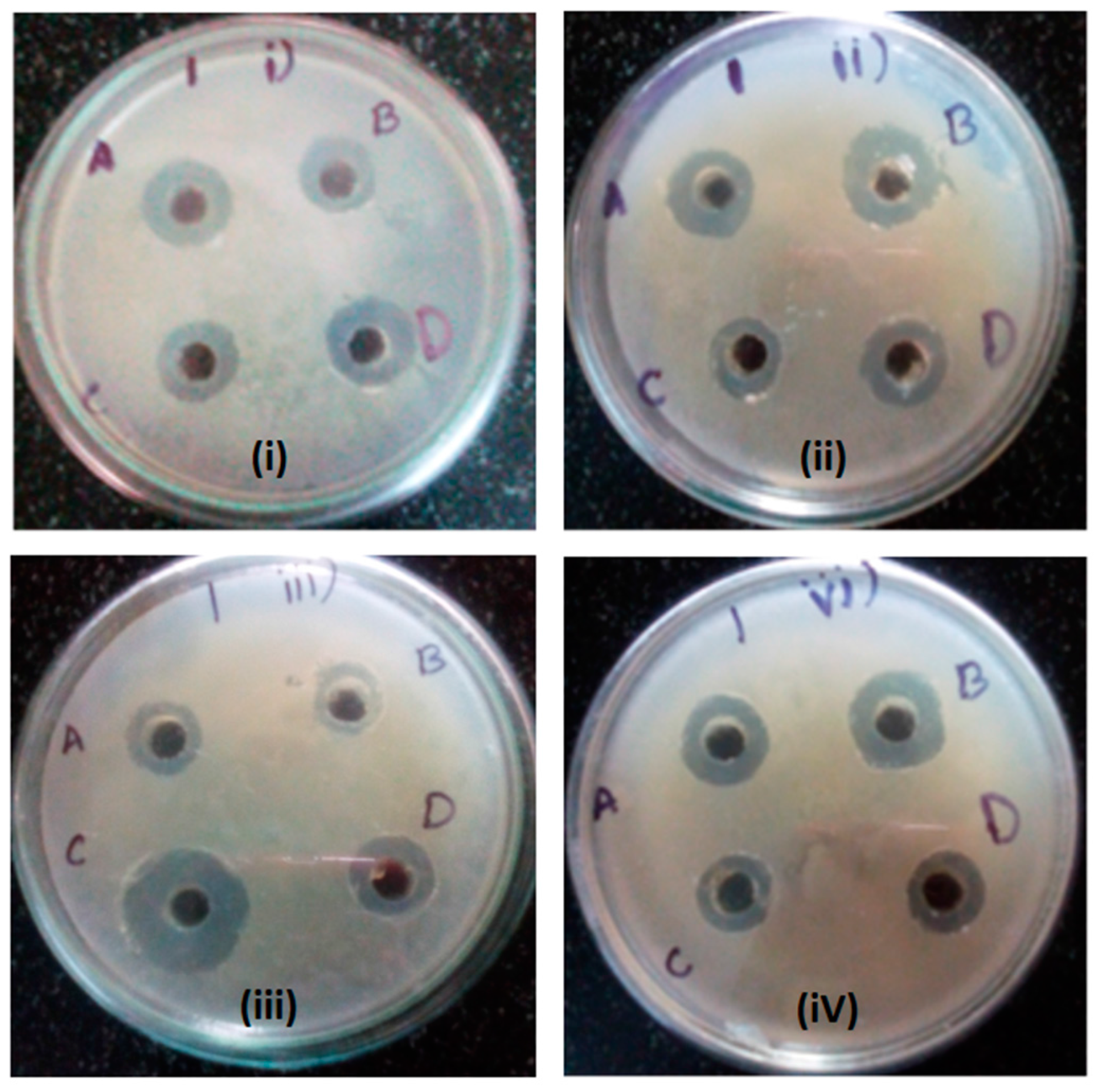
2.4. Antibacterial Activity
2.5. 2-D Gel Electrophoresis Results
3. Experimental Section
3.1. Materials
3.2. Preparation of Plant Seeds Extract
3.3. DEAE-Ion Exchange Chromatography
3.4. Bacterial Strains
3.5. Culture Medium and Inoculum Preparation
3.6. Agar Well Diffusion Assay
3.7. Determination of the Minimal Inhibitory Concentration (MIC)
3.8. Statistical Analysis
3.9. Two-Dimensional Electrophoresis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stintzi, A.; Heitz, T.; Prasad, V.; Wiedemann-Merdinoglu, S.; Kauffmann, S.; Geoffroy, P.; Legrand, M.; Fritig, B. Plant ‘pathogenesis-related’ proteins and their role in defense against pathogens. Biochimie 1993, 75, 687–706. [Google Scholar] [CrossRef]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current overview of allergens of plant pathogenesis related protein families. Sci. World J. 2014, 2014, 543195. [Google Scholar] [CrossRef] [PubMed]
- Egorov, T.A.; Odintsova, T.I.; Pukhalsky, V.A.; Grishin, E.V. Diversity of wheat anti-microbial peptides. Peptides 2005, 26, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.; Yedery, R.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Montalbán-López, M.; Sánchez-Hidalgo, M.; Cebrián, R.; Maqueda, M. Discovering the bacterial circular proteins: Bacteriocins, cyanobactins, and pilins. J. Biol. Chem. 2012, 287, 27007–27013. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.P.; Wong, C.T. Chemical synthesis of circular proteins. J. Biol. Chem. 2012, 287, 27020–27025. [Google Scholar] [CrossRef] [PubMed]
- Hammami, R.; Hamida, J.B.; Vergoten, G.; Fliss, I. Phytamp: A database dedicated to antimicrobial plant peptides. Nucleic Acids Res. 2009, 37, D963–D968. [Google Scholar] [CrossRef] [PubMed]
- Badgujar, S.B.; Patel, V.V.; Bandivdekar, A.H. Foeniculum vulgare Mill.: A review of its botany, phytochemistry, pharmacology, contemporary application, and toxicology. BioMed Res. Int. 2014, 2014, 842674. [Google Scholar] [CrossRef] [PubMed]
- Mohsenzadeh, M. Evaluation of antibacterial activity of selected Iranian essential oils against Staphylococcus aureus and Escherichia coli in nutrient broth medium. Pak. J. Biol. Sci. 2007, 10, 3693–3697. [Google Scholar] [PubMed]
- Leah, R.; Tommerup, H.; Svendsen, I.; Mundy, J. Biochemical and molecular characterization of three barley seed proteins with antifungal properties. J. Biol. Chem. 1991, 266, 1564–1573. [Google Scholar] [PubMed]
- Melchers, L.S.; Groot-de, M.A.; Knaap, J.A.; Ponstein, A.S.; Sela-Buurlage, M.B.; Bol, J.F.; Cornelissen, B.J.; Elzen, P.J.; Linthorst, H.J. A new class of tobacco chitinases homologous to bacterial exo-chitinases displays antifungal activity. Plant J. 1994, 5, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Broekaert, W.F.; Terras, F.; Cammue, B.; Osborn, R.W. Plant defensins: Novel antimicrobial peptides as components of the host defense system. Plant Physiol. 1995, 108, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.; Cammue, B.P.; Thevissen, K. Plant defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Florack, D.; Stiekema, W. Thionins: Properties, possible biological roles and mechanisms of action. Plant Mol. Biol. 1994, 26, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Cammue, B.P.A.; Thevissen, K.; Hendriks, M.; Eggermont, K.; Goderis, I.J.; Proost, P.; Van Damme, J.; Osborn, R.W.; Guerbette, F.; Kader, J.-C. A potent antimicrobial protein from onion seeds showing sequence homology to plant lipid transfer proteins. Plant Physiol. 1995, 109, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-S.; Samuel, D.; Liu, Y.-J.; Shyu, J.-C.; Lai, S.-M.; Lin, K.-F.; Lyu, P.-C. Binding mechanism of nonspecific lipid transfer proteins and their role in plant defense. Biochemistry 2004, 43, 13628–13636. [Google Scholar] [CrossRef] [PubMed]
- Agizzio, A.P.; Carvalho, A.O.; Suzanna de Fátima, F.R.; Machado, O.L.; Alves, E.W.; Okorokov, L.A.; Samarão, S.S.; Bloch, C.; Prates, M.V.; Gomes, V.M. A 2S albumin-homologous protein from passion fruit seeds inhibits the fungal growth and acidification of the medium by Fusarium oxysporum. Arch. Biochem. Biophys. 2003, 416, 188–195. [Google Scholar] [CrossRef]
- Terras, F.; Schoofs, H.; De Bolle, M.; Van Leuven, F.; Rees, S.B.; Vanderleyden, J.; Cammue, B.; Broekaert, W.F. Analysis of two novel classes of plant antifungal proteins from radish (Raphanus sativus L.) seeds. J. Biol. Chem. 1992, 267, 15301–15309. [Google Scholar] [PubMed]
- Barbieri, L.; Battelli, M.G.; Stirpe, F. Ribosome-inactivating proteins from plants. BBA-Biomembranes 1993, 1154, 237–282. [Google Scholar] [CrossRef]
- Dong, T.; Ng, T.; Yeung, H.; Wong, R. Isolation and characterization of a novel ribosome-inactivating protein, β-kirilowin, from the seeds of Trichosanthes kirilowii. Biochem. Biophys. Res. Commun. 1994, 199, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-W.; Vepachedu, R.; Sharma, N.; Vivanco, J.M. Ribosome-inactivating proteins in plant biology. Planta 2004, 219, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Jach, G.; Görnhardt, B.; Mundy, J.; Logemann, J.; Pinsdorf, E.; Leah, R.; Schell, J.; Maas, C. Enhanced quantitative resistance against fungal disease by combinatorial expression of different barley antifungal proteins in transgenic tobacco. Plant J. 1995, 8, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Hoda, H.; Elmi, F.; Elmi, M.M. Electrophoretic pattern and antibacterial activity of proteins from vicia faba seed extract. Int. J. Mol. Cell. Med. 2013, 2, 46–49. [Google Scholar] [PubMed]
- Imade, P.E.; Izekor, P.E.; Eghafona, N.O.; Enabulele, O.I.; Ophori, E. Asymptomatic bacteriuria among pregnant women. N. Am. J. Med. Sci. 2010, 2, 263–266. [Google Scholar] [PubMed]
- Alviano, D.; Alviano, C. Plant extracts: Search for new alternatives to treat microbial diseases. Curr. Pharm. Biotechnol. 2009, 10, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Cseke, L.; Podila, G. MADS-box genes in dioecious aspen II: A review of MADS-box genes from trees and their potential in forest biotechnology. Hysiol. Mol. Biol. Plants 2004, 10, 7–28. [Google Scholar]
- Freed, J.K.; Smith, J.R.; Li, P.; Greene, A.S. Isolation of signal transduction complexes using biotin and crosslinking methodologies. Proteomics 2007, 7, 2371–2374. [Google Scholar] [CrossRef] [PubMed]
- Görg, A.; Weiss, W.; Dunn, M.J. Current two-dimensional electrophoresis technology for proteomics. Proteomics 2004, 4, 3665–3685. [Google Scholar] [CrossRef] [PubMed]
- Barciszewski, J.; Maciej, S.; Haertlé, T. Minireview: Analysis of rape seed napin structure and potential roles of the storage protein. J. Protein Chem. 2000, 19, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Gehrig, P.; Biemann, K. Assignment of the disulfide bonds in napin, a seed storage protein from Brassica napus, using matrix-assisted laser desorption ionization mass spectrometry. Pept. Res. 1995, 9, 308–314. [Google Scholar]
- Yamada, K.; Hirota, M.; Niimi, Y.; Nguyen, H.A.; Takahara, Y.; Kamio, Y.; Kaneko, J. Nucleotide sequences and organization of the genes for carotovoricin (Ctv) from Erwinia carotovora indicate that Ctv evolved from the same ancestor as Salmonella typhi prophage. Biosci. Biotechnol. Biochem. 2006, 70, 2236–2247. [Google Scholar] [CrossRef] [PubMed]
- Feil, H.; Feil, W.S.; Chain, P.; Larimer, F.; DiBartolo, G.; Copeland, A.; Lykidis, A.; Trong, S.; Nolan, M.; Goltsman, E. Comparison of the complete genome sequences of Pseudomonas syringae pv. syringae B728a and pv. tomato DC3000. Proc. Natl. Acad. Sci. USA 2005, 102, 11064–11069. [Google Scholar] [CrossRef] [PubMed]
- Chuang, D.-Y.; Chien, Y.-C.; Wu, H.-P. Cloning and expression of the Erwinia carotovora subsp. carotovora gene encoding the low-molecular-weight bacteriocin carocin S1. J. Bacteriol. 2007, 189, 620–626. [Google Scholar] [PubMed]
- Ochiai, H.; Inoue, Y.; Takeya, M.; Sasaki, A. Genome sequence of Xanthomonas oryzae pv. oryzae suggests contribution of large numbers of effector genes and insertion sequences to its race diversity. Jpn. Agric. Res. Q. JARQ 2005, 39, 275–287. [Google Scholar]
- Hawkey, P.; Lewis, D. Medical Bacteriology: A Practical Approach; OUP Oxford: Oxford, UK, 2003; Volume 265. [Google Scholar]
- Camporese, A.; Balick, M.; Arvigo, R.; Esposito, R.; Morsellino, N.; De Simone, F.; Tubaro, A. Screening of anti-bacterial activity of medicinal plants from Belize (central America). J. Ethnopharmacol. 2003, 87, 103–107. [Google Scholar] [CrossRef]
- Black, J.G. Microbiology: Principles and Explorations; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| S. No | Plant | Sodium Phosphate Citrate Buffer (pH-7.2) (μg/mL) | CTAB Buffer (pH-6.0) (μg/mL) | Sodium Acetate Buffer (pH-6.5) (μg/mL) |
|---|---|---|---|---|
| 1. | Foeniculum vulgare Mill. | 80 | 80 | 140 |
| S. No | S1 | Chl (25 mcg/mL) | Cipro (100 mcg/mL) |
|---|---|---|---|
| S. aureus | 12 | 21 | 16 |
| E. coli | 11 | 8 | 14 |
| P. aeruginosa | 12.5 | 8 | 12 |
| P. vulgaris | 12 | 8 | 14 |
| S. No | Elute 1 | Elute 2 | Elute 3 | Elute 4 |
|---|---|---|---|---|
| Foeniculum vulgare Mill. | 120 | 120 | 110 | 100 |
| S. No | Elute 1 | Elute 2 | Elute 3 | Elute 4 | Cipro (100 mcg) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| ZOI | IC50 Value | ZOI | IC50 Value | ZOI | IC50 Value | ZOI | IC50 Value | ZOI | IC50 Value | |
| S. aureus | 10 | 27.64 | 8 | 25.91 | 8 | 21.27 | 12 | 20.8 | 16 | 160.529 |
| E. coli | 13 | 67.56 | 12 | 64.12 | 6 | 60.52 | 8 | 41.06 | 14 | 92.489 |
| P. aeruginosa | 5 | 28.01 | 4 | 68.33 | 16 | 25.02 | 10 | 26.67 | 12 | 144.634 |
| P. vulgaris | 12 | 59.68 | 13 | 57.83 | 6 | 41.25 | 7 | 35.67 | 14 | 72.685 |
| Bacterial Strains | Elute 1 | Elute 2 | Elute 3 | Elute 4 | Cipro | t Value | p-Value * |
|---|---|---|---|---|---|---|---|
| IC50 | IC50 | IC50 | IC50 | IC50 | |||
| S. aureus | 27.64 | 25.91 | 21.27 | 20.8 | 160.529 | 6.2 | 0.025 |
| E. coli | 67.56 | 64.12 | 60.52 | 41.06 | 92.489 | 2.39 | 0.139 |
| P. aeruginosa | 28.01 | 68.33 | 25.02 | 26.67 | 144.634 | 3.4 | 0.077 |
| P. vulgaris | 59.68 | 57.83 | 41.25 | 35.67 | 72.685 | 6.5 | 0.023 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Akeel, R.; Mateen, A.; Syed, R.; Alyousef, A.A.; Shaik, M.R. Screening, Purification and Characterization of Anionic Antimicrobial Proteins from Foeniculum Vulgare. Molecules 2017, 22, 602. https://doi.org/10.3390/molecules22040602
Al Akeel R, Mateen A, Syed R, Alyousef AA, Shaik MR. Screening, Purification and Characterization of Anionic Antimicrobial Proteins from Foeniculum Vulgare. Molecules. 2017; 22(4):602. https://doi.org/10.3390/molecules22040602
Chicago/Turabian StyleAl Akeel, Raid, Ayesha Mateen, Rabbani Syed, Abdullah A. Alyousef, and Mohammed Rafi Shaik. 2017. "Screening, Purification and Characterization of Anionic Antimicrobial Proteins from Foeniculum Vulgare" Molecules 22, no. 4: 602. https://doi.org/10.3390/molecules22040602
APA StyleAl Akeel, R., Mateen, A., Syed, R., Alyousef, A. A., & Shaik, M. R. (2017). Screening, Purification and Characterization of Anionic Antimicrobial Proteins from Foeniculum Vulgare. Molecules, 22(4), 602. https://doi.org/10.3390/molecules22040602