
Optimization of EPS Production and Characterization by a Halophilic Bacterium, Kocuria rosea ZJUQH from Chaka Salt Lake with Response Surface Methodology


Abstract
:1. Introduction
2. Results
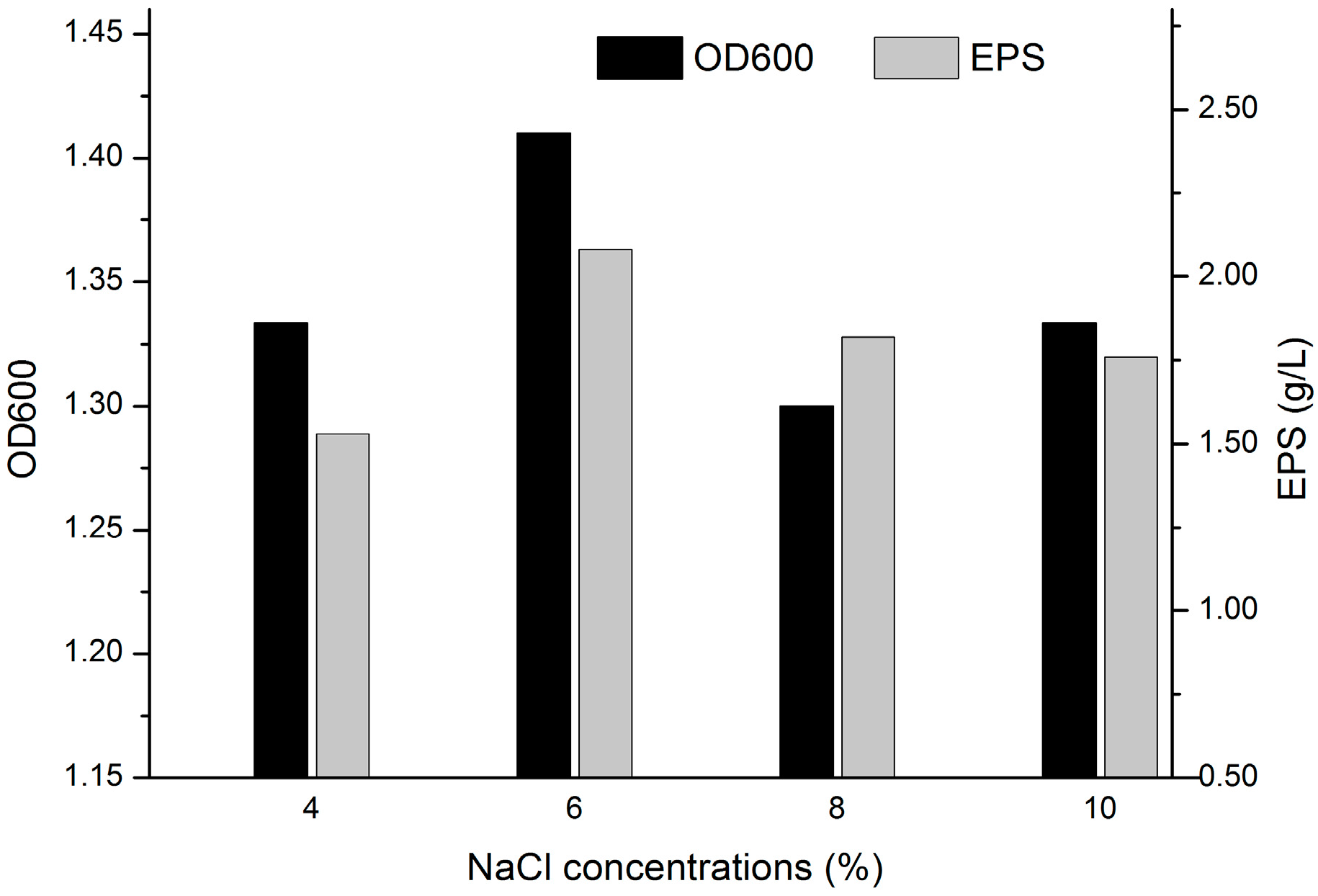
2.1. Determination of the Salt Tolerance
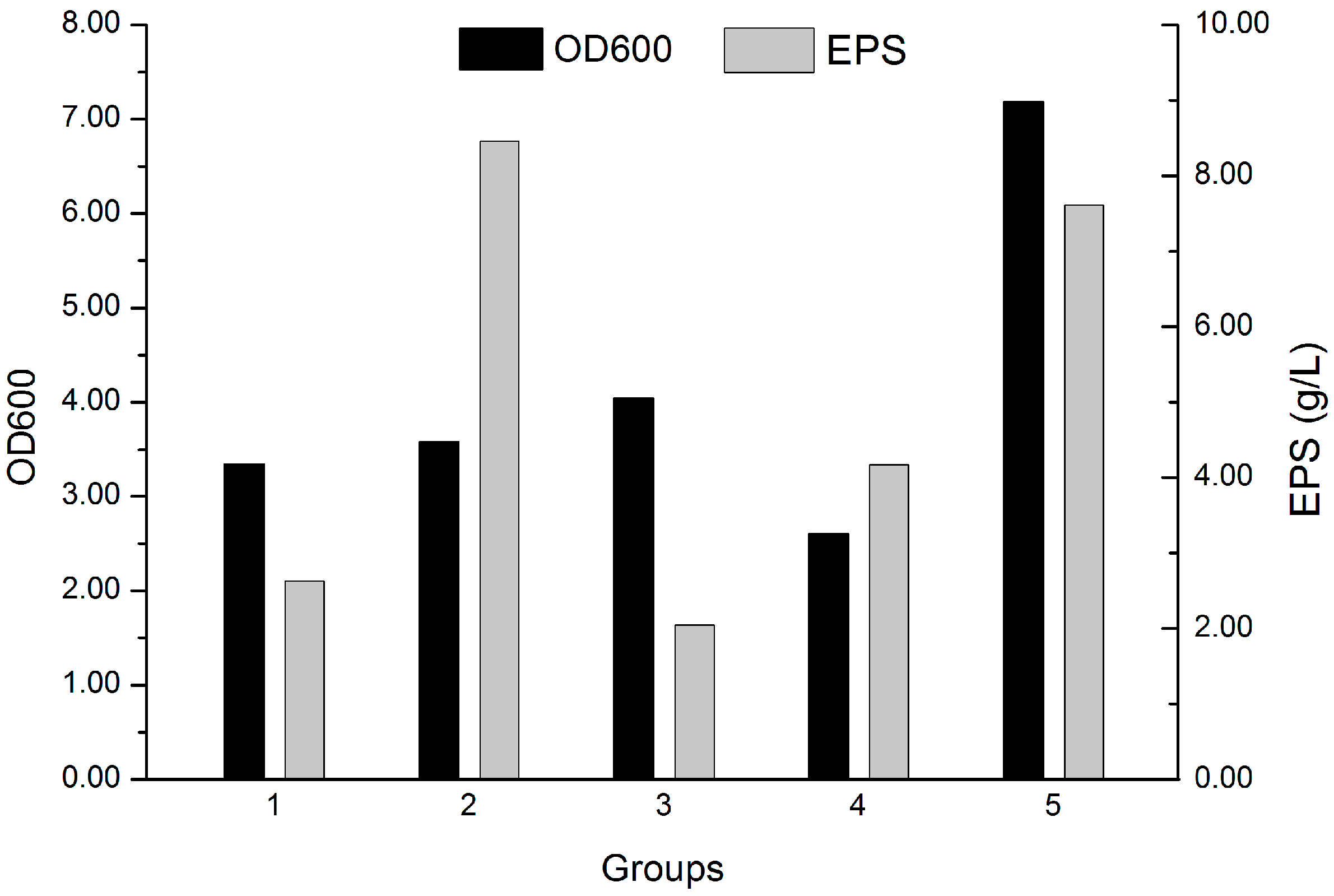
2.2. Screening of the Fermentation Culture Media
2.3. Optimization of Culture Media Compositions for EPS Production
2.3.1. Screening Stage: Half Fractional Factorial Design
2.3.2. The Steepest Ascent Experiment
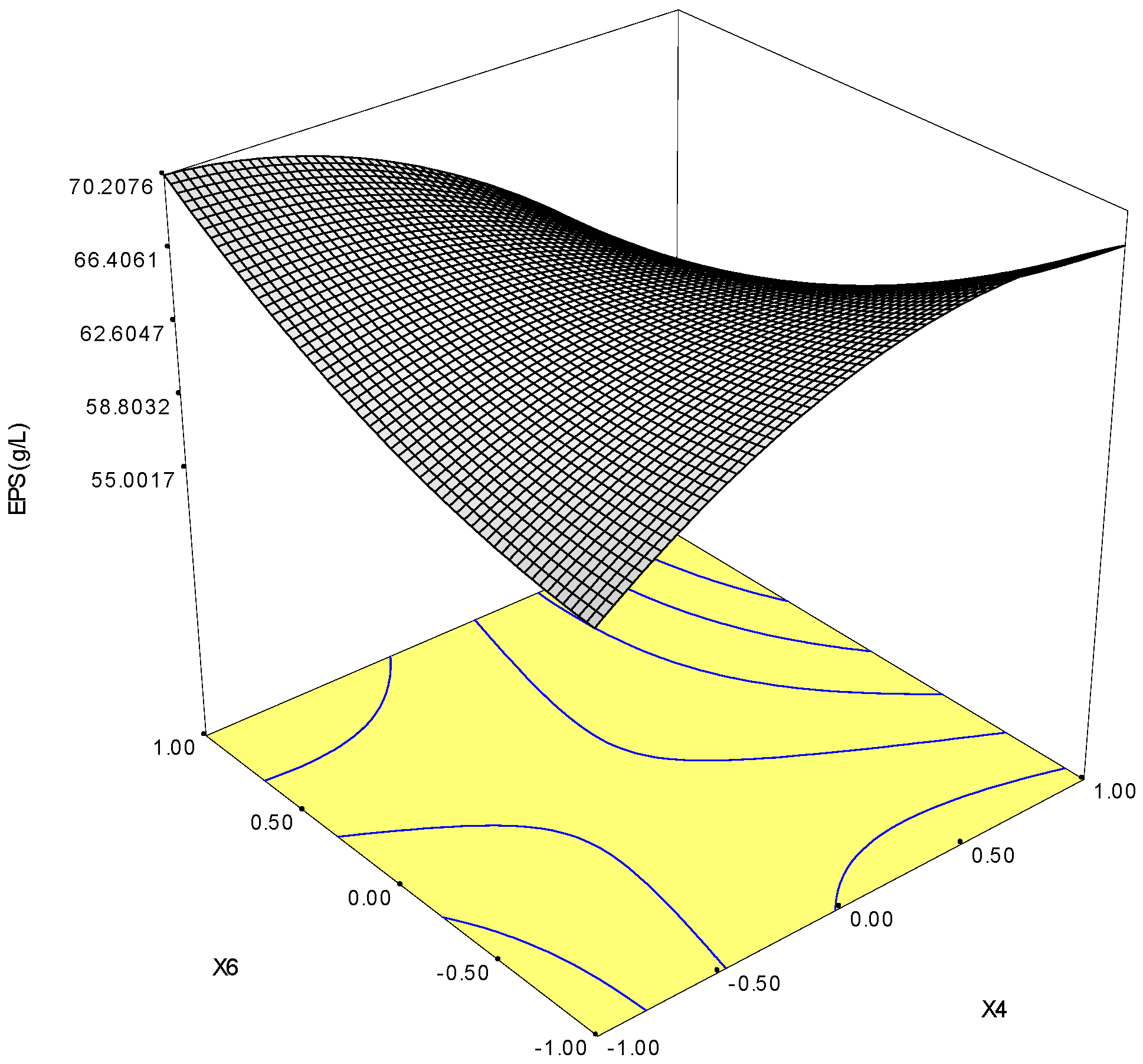
2.3.3. Optimization of KCl and MgSO4 for the Maximal Production of EPSs
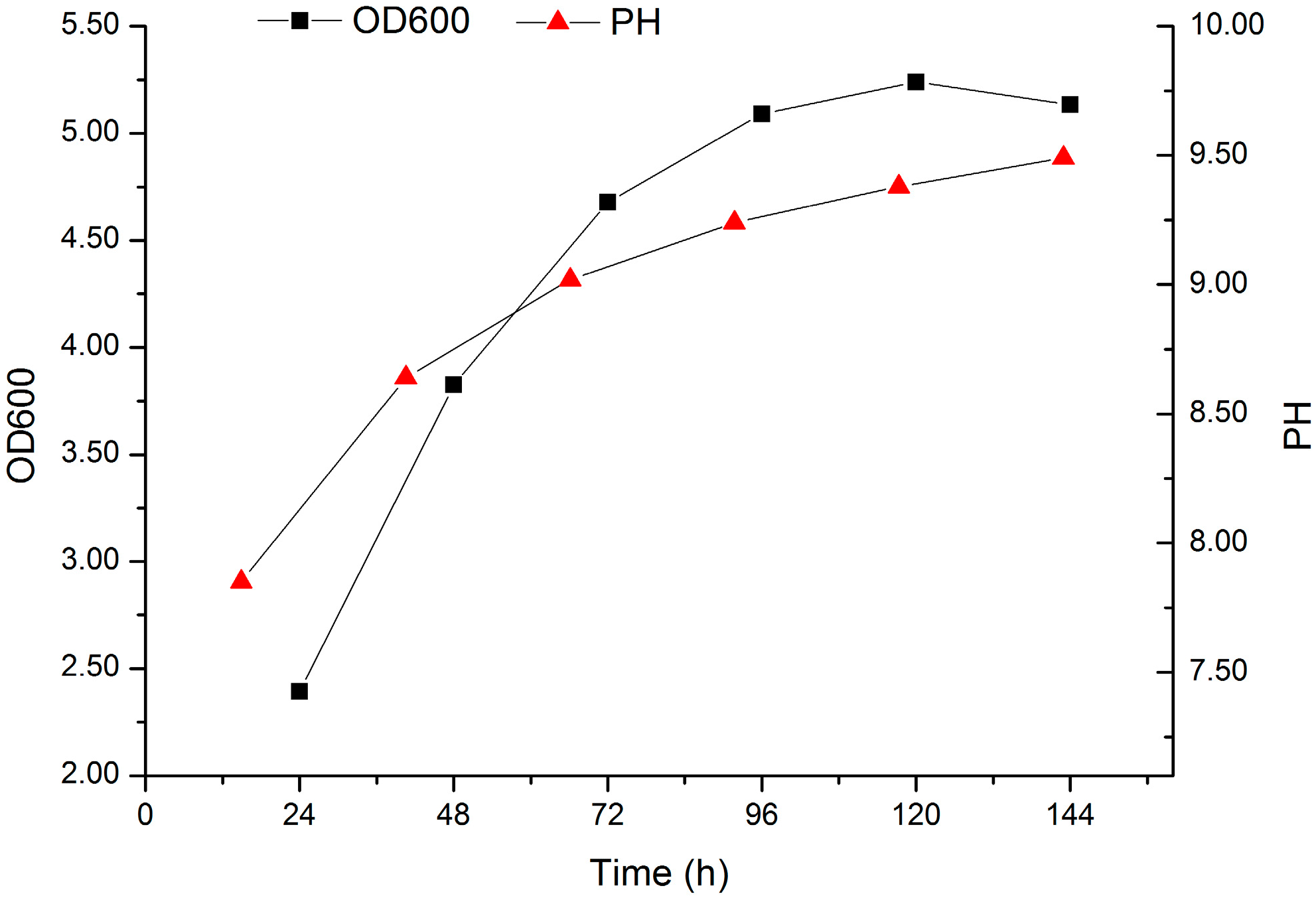
2.4. Validation of the Optimized Culture Conditions
2.5. Effects of KCl and MgSO4 on EPS Production
2.6. Validation of the Essentiality of MgSO4
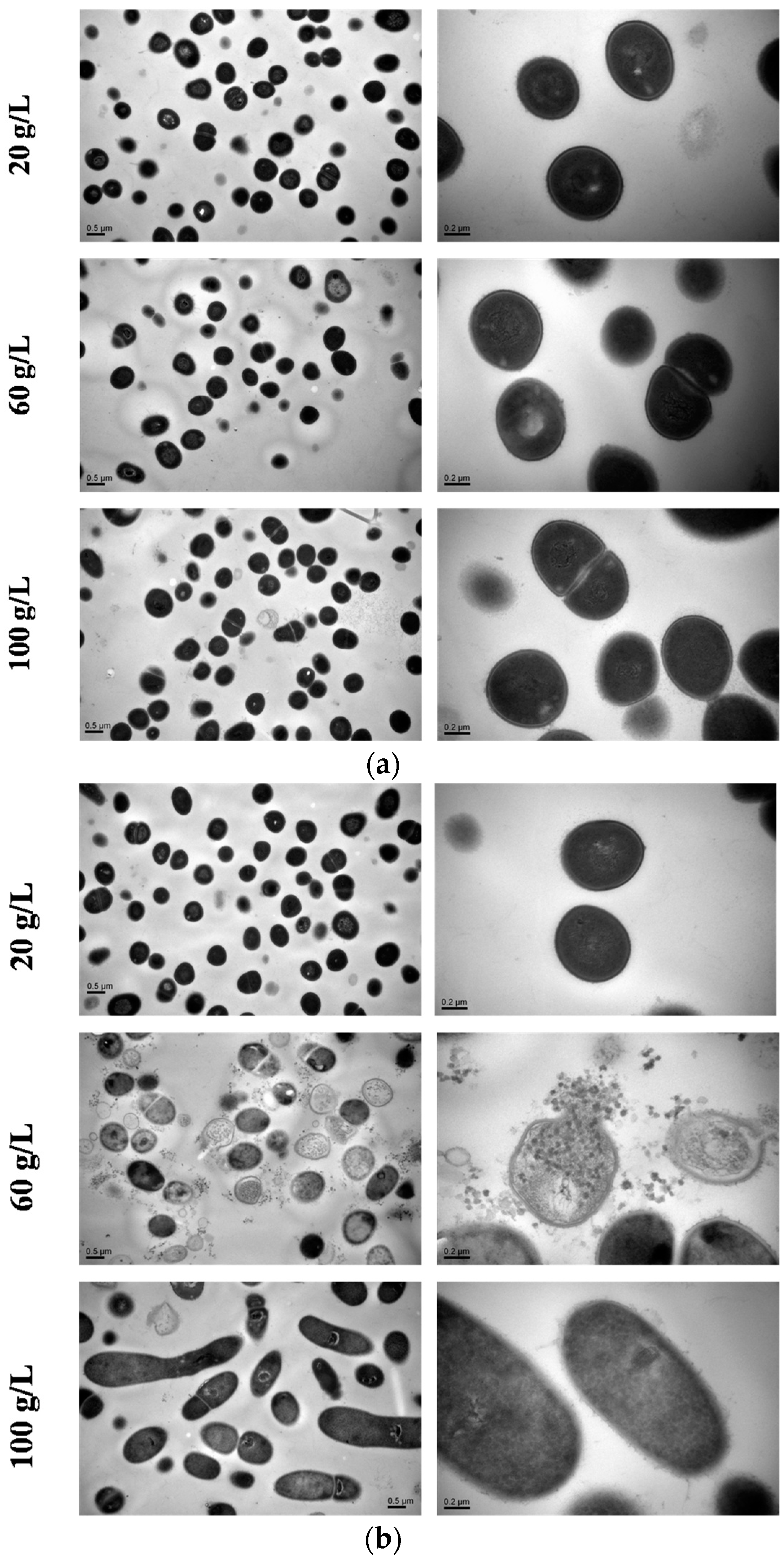
2.7. TEM Observation
2.8. Characterization of EPS
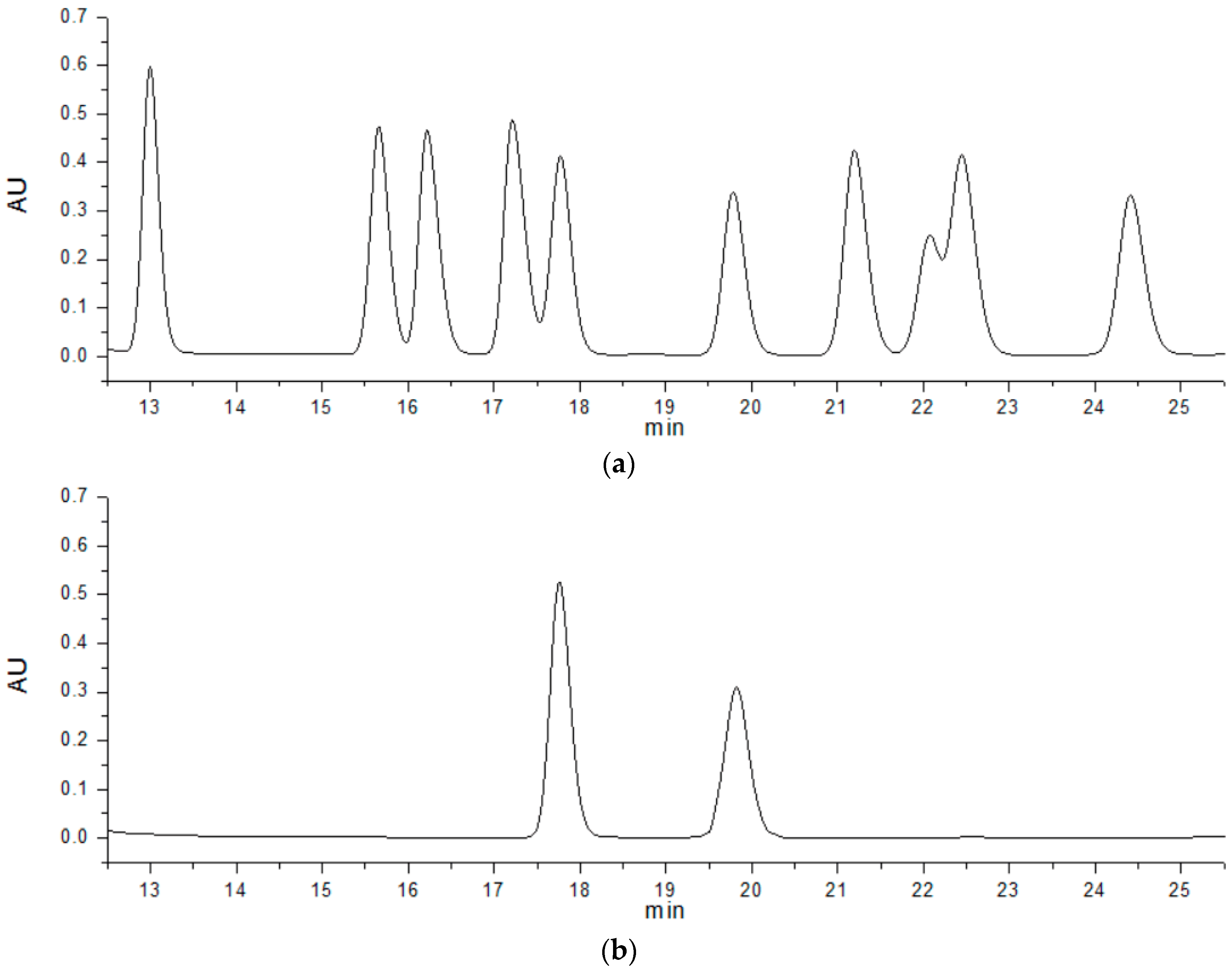
2.8.1. Molecular Mass Determination and Analysis of Monosaccharide Composition
2.8.2. Determination of Total Sugar, Uronic Sugar and Protein
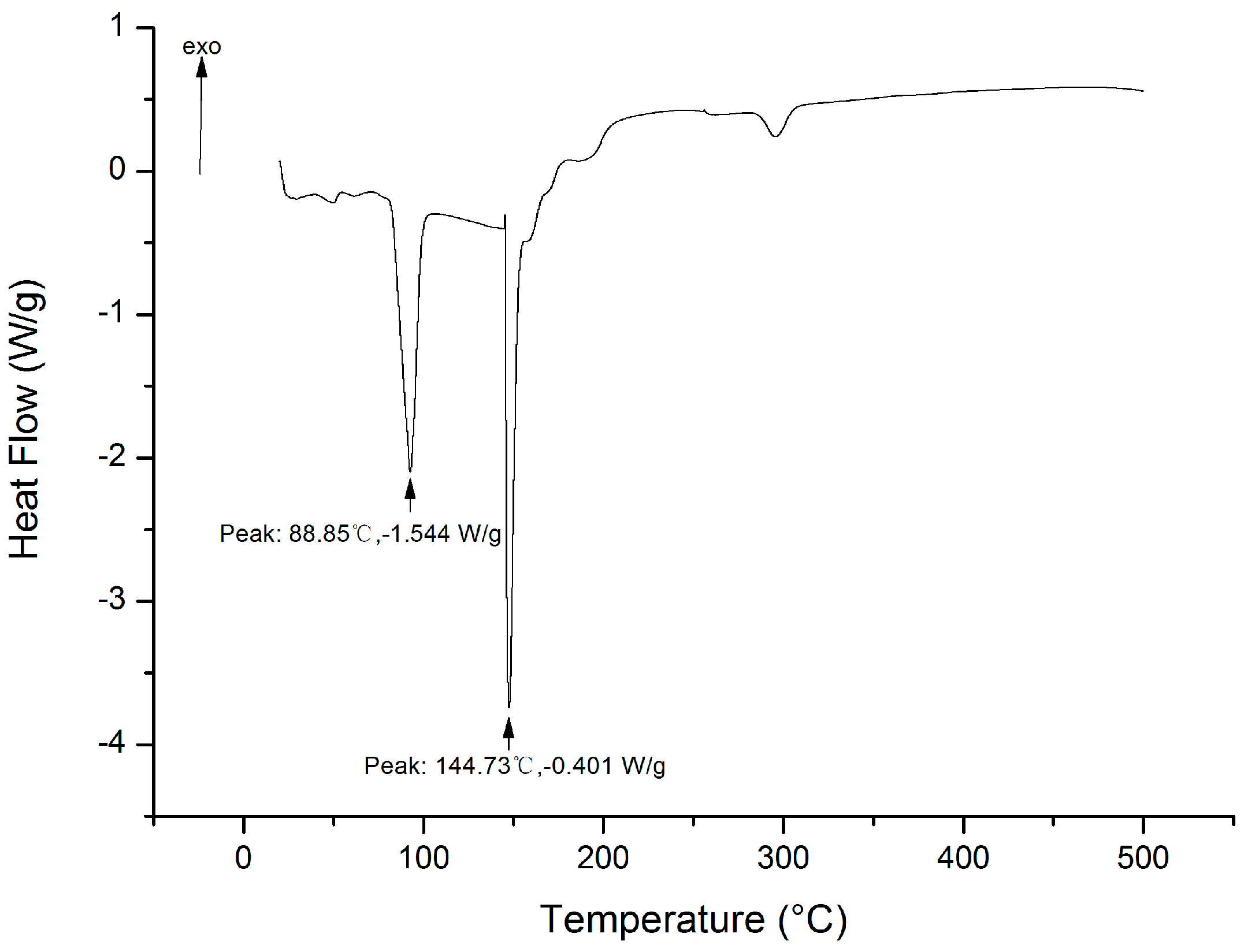
2.8.3. DSC Analysis
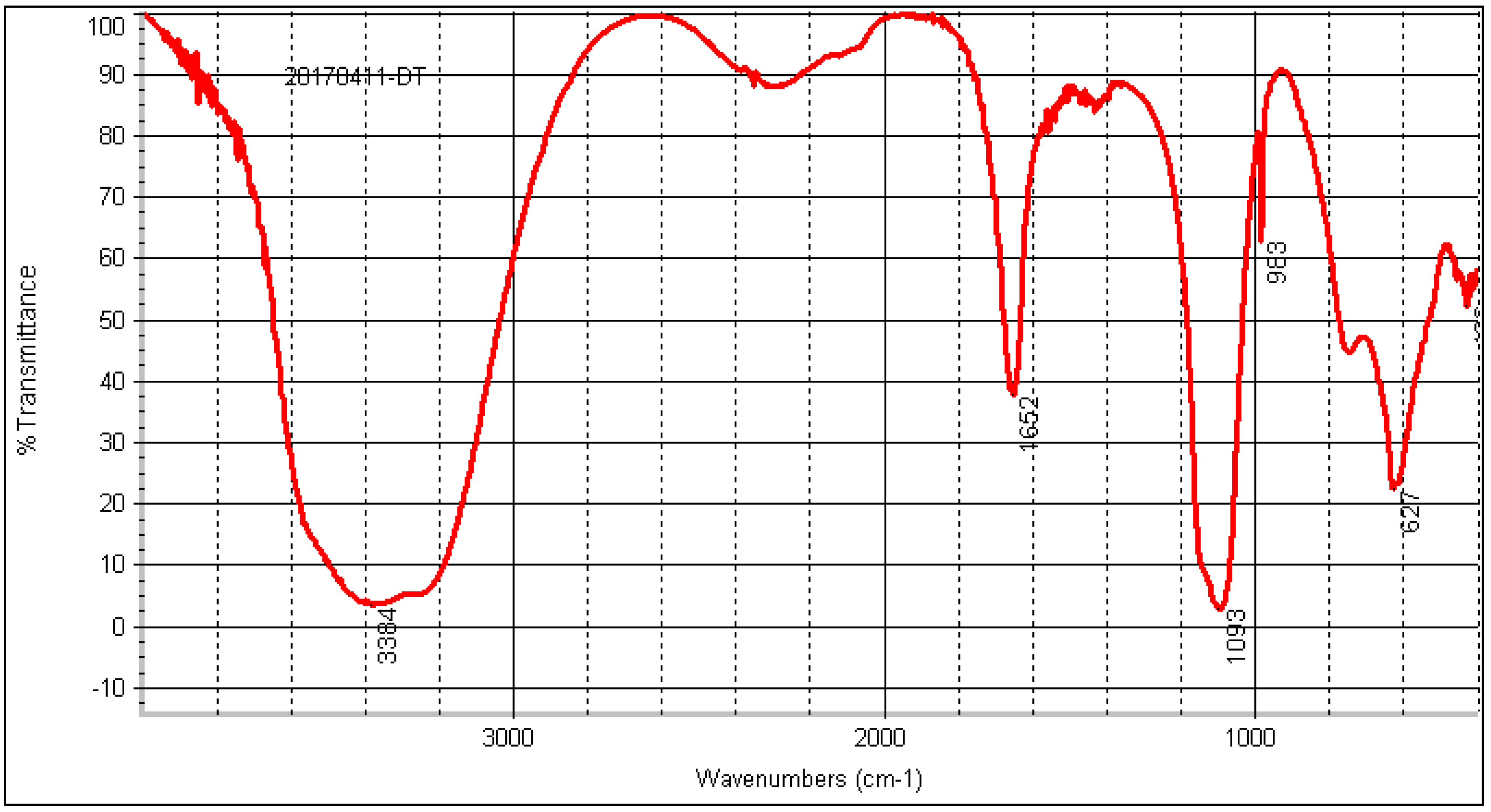
2.8.4. FT-IR Analysis
3. Discussion
4. Materials and Methods
4.1. Microorganism and Culture Media
4.2. Culture Conditions
4.3. Determination of Salt Tolerance
4.4. Screening of the Fermentation Culture Compositions
4.5. Effects of KCl and MgSO4 on EPS Production
4.6. Validation of the Essentiality of MgSO4
4.7. Transmission Electron Microscopy (TEM)
- Double fixation: The specimen was first fixed with 2.5% glutaraldehyde in phosphate buffer (0.1M, pH 7.0) for more than 4 h; washed three times in the phosphate buffer (0.1 M, pH 7.0) for 15 min at each step; then postfixed with 1% OsO4 in phosphate buffer (0.1 M, pH 7.0) for 1–2 h and washed three times in the phosphate buffer (0.1 M, pH 7.0) for 15 min at each step.
- Dehydration: The specimen was first dehydrated by a graded series of ethanol (30%, 50%, 70%, 80%, 90%, 95%, and 100%) for about 15 to 20 min at each step, then transferred to absolute acetone for 20 min.
- Infiltration: The specimen was placed in 1:1 mixture of absolute acetone and final Spurr resin mixture for 1 h at room temperature, then transferred to 1:3 mixture of absolute acetone and the final resin mixture for 3 h and to final Spurr resin mixture for overnight.
- Embedding and ultrathin sectioning: Specimen was placed in an Eppendorf containing Spurr resin and heated at 70 °C for more than 9 h. The specimen was sectioned in LEICA EM UC7 ultratome and sections were stained by uranyl acetate and alkaline lead citrate for 5 to 10 min respectively and observed in a Hitachi Model H-7650 TEM.
4.8. Analytical Methods
4.8.1. Determination of Cell Dry Weight and Cell Biomass
4.8.2. Recovery of Native Crude EPSs and Quantification
4.9. Characterization of EPS
4.9.1. Purification of EPS
4.9.2. Molecular Mass Determination and Analysis of Monosaccharide Composition
4.9.3. Determination of Total Sugar, Uronic Sugar, and Protein
4.9.4. DSC Measurements
4.9.5. FT-IR Spectroscopy Analysis
4.10. Experimental Design
4.10.1. Fractional Factorial Designs (FFD)
4.10.2. Central Composite Design (CCD)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Castillo-Carvajal, L.C.; Sanz-Martin, J.L.; Barragan-Huerta, B.E. Biodegradation of organic pollutants in saline wastewater by halophilic microorganisms: A review. Environ. Scie. Pollut. Res. Int. 2014, 21, 9578–9588. [Google Scholar] [CrossRef] [PubMed]
- Jindal, N.; Singh, D.P.; Khattar, J.I.S. Kinetics and physico-chemical characterization of exopolysaccharides produced by the cyanobacterium Oscillatoria formosa. World J. Microbiol. Biotechnol. 2011, 27, 2139–2146. [Google Scholar] [CrossRef]
- Sardari, R.R.R.; Kulcinskaja, E.; Ron, E.Y.C.; Björnsdóttir, S.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó.; Karlsson, E.N. Evaluation of the production of exopolysaccharides by two strains of the thermophilic bacterium Rhodothermus marinus. Carbohydr. Polym. 2017, 156, 1–8. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; De Vin, F.; Vaningelgem, F.; Degeest, B. Recent developments in the biosynthesis and applications of heteropolysaccharides from lactic acid bacteria. Int. Dairy J. 2001, 11, 687–707. [Google Scholar] [CrossRef]
- Laws, A.; Gu, Y.; Marshall, V. Biosynthesis, characterisation, and design of bacterial exopolysaccharides from lactic acid bacteria. Biotechnol. Adv. 2001, 19, 597–625. [Google Scholar] [CrossRef]
- De Vuyst, L.; Vanderveken, F.; Van de Ven, S.; Degeest, B. Production by and isolation of exopolysaccharides from Streptococcus thermophilus grown in a milk medium and evidence for their growth-associated biosynthesis. J. Appl. Microbiol. 1998, 84, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Mata, J.A.; Béjar, V.; Llamas, I.; Arias, S.; Bressollier, P.; Tallon, R.; Urdaci, M.C.; Quesada, E. Exopolysaccharides produced by the recently described halophilic bacteria Halomonas ventosae and Halomonas anticariensis. Res. Microbiol. 2006, 157, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and characterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, H.P.; Ray, R.M.; Patel, R.M. Exopolymer production by Bacillus species. Carbohydr. Polym. 1997, 34, 323–327. [Google Scholar] [CrossRef]
- Nikinmaa, M.; Alam, S.A.; Raulio, M.; Katina, K.; Kajala, I.; Nordlund, E.; Sozer, N. Bioprocessing of bran with exopolysaccharide producing microorganisms as a tool to improve expansion and textural properties of extruded cereal foams with high dietary fibre content. LWT Food Sci. Technol. 2017, 77, 170–177. [Google Scholar] [CrossRef]
- Yamada, T.; Ogamo, A.; Saito, T.; Watanabe, J.; Uchiyama, H.; Nakagawa, Y. Preparation and anti-HIV activity of low-molecular-weight carrageenans and their sulfated derivatives. Carbohydr. Polym. 1997, 32, 51–55. [Google Scholar] [CrossRef]
- Sutherland, I.W. Novel and established applications of microbial polysaccharides. Trends Biotechnol. 1998, 16, 41–46. [Google Scholar] [CrossRef]
- DeAngelis, P.L. Glycosaminoglycan polysaccharide biosynthesis and production: Today and tomorrow. Appl. Microbiol. Biotechnol. 2012, 94, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Sathiyanarayanan, G.; Seghal Kiran, G.; Selvin, J. Synthesis of silver nanoparticles by polysaccharide bioflocculant produced from marine Bacillus subtilis MSBN17. Colloids Surf. B Biointerfaces 2013, 102, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, O.A.; Hao, X.; Xie, P.; Hatab, S.; Lin, Y.; Wei, G. Biosorption of Copper (II) from Aqueous Solution Using Non-Living Mesorhizobium amorphae Strain CCNWGS0123. Microbes Environ. 2012, 27, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ahmed, Z.; Feng, W.; Li, C.; Song, S. Physicochemical properties of exopolysaccharide produced by Lactobacillus kefiranofaciens ZW3 isolated from Tibet kefir. Int. J. Biol. Macromol. 2008, 43, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.A.; Hasan, F.; Hameed, A.; Ahmed, S. Biological degradation of plastics: A comprehensive review. Biotechnol. Adv. 2008, 26, 246–265. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Gurevich, P.; Azachi, M.; Henis, Y. Microbial degradation of pollutants at high salt concentrations. Biodegradation 1992, 3, 387–398. [Google Scholar] [CrossRef]
- Zhuang, X.L.; Han, Z.; Bai, Z.H.; Zhuang, G.Q.; Shim, H.J. Progress in decontamination by halophilic microorganisms in saline wastewater and soil. Environ. Pollut. 2010, 158, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Joyce, A.P.; Leung, S.S. Use of response surface methods and path of steepest ascent to optimize ligand-binding assay sensitivity. J. Immunol. Methods 2013, 392, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Podell, S.; Emerson, J.B.; Jones, C.M.; Ugalde, J.A.; Welch, S.; Heidelberg, K.B.; Banfield, J.F.; Allen, E.E. Seasonal fluctuations in ionic concentrations drive microbial succession in a hypersaline lake community. ISME J. 2014, 8, 979–990. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Life at high salt concentrations, intracellular KCl concentrations, and acidic proteomes. Front. Microbiol. 2013, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-S. Renewable resource-based composites of recycled natural fibers and maleated polylactide bioplastic: Characterization and biodegradability. Polym. Degrad. Stab. 2009, 94, 1076–1084. [Google Scholar] [CrossRef]
- Lin, L.; Xie, J.; Liu, S.; Shen, M.; Tang, W.; Xie, M. Polysaccharide from Mesona chinensis: Extraction optimization, physicochemical characterizations and antioxidant activities. Int. J. Biol. Macromol. 2017, 99, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.-L.; Tang, Z.-H.; Zhang, X.-F.; Zhong, Y.-H.; Yao, S.-Z.; Wang, L.-S.; Lin, C.-W.; Luo, X. Chemical properties and antioxidant activity of a water-soluble polysaccharide from Dendrobium officinale. Int. J. Biol. Macromol. 2016, 89, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Galinski, E.A.; Grant, W.D.; Oren, A.; Ventosa, A. Halophiles 2010: Life in saline environments. Appl. Environ. Microbiol. 2010, 76, 6971–6981. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Heldal, M.; Norland, S.; Galinski, E.A. Intracellular ion and organic solute concentrations of the extremely halophilic bacterium Salinibacter ruber. Extrem. Life Extrem. Cond. 2002, 6, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Epstein, W. The roles and regulation of potassium in bacteria. Prog. Nucleic Acid Res. Mol. Biol. 2003, 75, 293–320. [Google Scholar] [PubMed]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Pastor, J.M.; Bernal, V.; Salvador, M.; Argandona, M.; Vargas, C.; Csonka, L.; Sevilla, A.; Iborra, J.L.; Nieto, J.J.; Canovas, M. Role of Central Metabolism in the Osmoadaptation of the Halophilic Bacterium Chromohalobacter salexigens. J. Biol. Chem. 2013, 288, 17769–17781. [Google Scholar] [CrossRef] [PubMed]
- Doan, V.-T.; Guzman, H.; Mai, T.-H.; Hatti-Kaul, R. Ectoine Production by Halomonas boliviensis: Optimization Using Response Surface Methodology. Mar. Biotechnol. 2010, 12, 586–593. [Google Scholar]
- Tao, P.; Li, H.; Yu, Y.; Gu, J.; Liu, Y. Ectoine and 5-hydroxyectoine accumulation in the halophile Virgibacillus halodenitrificans PDB-F2 in response to salt stress. Appl. Microbiol. Biotechnol. 2016, 100, 6779–6789. [Google Scholar] [CrossRef] [PubMed]
- Amjres, H.; Béjar, V.; Quesada, E.; Carranza, D.; Abrini, J.; Sinquin, C.; Ratiskol, J.; Colliec-Jouault, S.; Llamas, I. Characterization of haloglycan, an exopolysaccharide produced by Halomonas stenophila HK30. Int. J. Biol. Macromol. 2015, 72, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Palaniraj, A.; Jayaraman, V. Production, recovery and applications of xanthan gum by Xanthomonas campestris. J. Food Eng. 2011, 106, 1–12. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, Y.; Phillips, G.O.; Yang, G. Utilization of bacterial cellulose in food. Food Hydrocoll. 2014, 35, 539–545. [Google Scholar] [CrossRef]
- Yu, L.; Xu, X.; Zhou, J.; Lv, G.; Chen, J. Chain conformation and rheological behavior of exopolysaccharide from Bacillus mucilaginosus SM-01. Food Hydrocoll. 2017, 65, 165–174. [Google Scholar] [CrossRef]
- Chang, C.; Zhang, L. Cellulose-based hydrogels: Present status and application prospects. Carbohydr. Polym. 2011, 84, 40–53. [Google Scholar] [CrossRef]
- Rasulov, B.; Rozi, P.; Pattaeva, M.; Yili, A.; Aisa, H. Exopolysaccharide-Based Bioflocculant Matrix of Azotobacter chroococcum XU1 for Synthesis of AgCl Nanoparticles and Its Application as a Novel Biocidal Nanobiomaterial. Materials 2016, 9, 528. [Google Scholar] [CrossRef]
- Chen, T.; Xu, P.; Zong, S.; Wang, Y.; Su, N.; Ye, M. Purification, structural features, antioxidant and moisture-preserving activities of an exopolysaccharide from Lachnum YM262. Bioorg. Med. Chem. Lett. 2017, 27, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Morris, E.R.; Nishinari, K.; Rinaudo, M. Gelation of gellan—A review. Food Hydrocoll. 2012, 28, 373–411. [Google Scholar] [CrossRef]
- Kaur, V.; Bera, M.B.; Panesar, P.S.; Kumar, H.; Kennedy, J.F. Welan gum: Microbial production, characterization, and applications. Int. J. Biol. Macromol. 2014, 65, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- Cerning, J. Exocellular polysaccharides produced by lactic-acid bacteria. FEMS Microbiol. Lett. 1990, 87, 113–130. [Google Scholar] [CrossRef]
- Looijesteijn, P.J.; Boels, I.C.; Kleerebezem, M.; Hugenholtz, J. Regulation of exopolysaccharide production by Lactococcus lactis subsp. cremoris by the sugar source. Appl. Environ. Microbiol. 1999, 65, 5003–5008. [Google Scholar] [PubMed]
- He, P.; Geng, L.; Wang, Z.; Mao, D.; Wang, J.; Xu, C. Fermentation optimization, characterization and bioactivity of exopolysaccharides from Funalia trogii. Carbohydr. Polym. 2012, 89, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Zohra, R.R.; Aman, A.; Ansari, A.; Haider, M.S.; Qader, S.A.U. Purification, characterization and end product analysis of dextran degrading endodextranase from Bacillus licheniformis KIBGE-IB25. Int. J. Biol. Macromol. 2015, 78, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Bednar, B.; Hennessey, J.P. Molecular size analysis of capsular polysaccharide preparations from Streptococcus pneumoniae. Carbohydr. Res. 1993, 243, 115–130. [Google Scholar] [CrossRef]
- Wei, C.-Y.; Li, W.-Q.; Shao, S.-S.; He, L.; Cheng, J.; Han, S.; Liu, Y. Structure and chain conformation of a neutral intracellular heteropolysaccharide from mycelium of Paecilomyces cicadae. Carbohydr. Polym. 2016, 136, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Luo, D. Identification of structure and antioxidant activity of a fraction of polysaccharide purified from Dioscorea nipponica Makino. Carbohydr. Polym. 2008, 71, 544–549. [Google Scholar] [CrossRef]
- Xu, R.; Shen, Q.; Ding, X.; Gao, W.; Li, P. Chemical characterization and antioxidant activity of an exopolysaccharide fraction isolated from Bifidobacterium animalis RH. Eur. Food Res. Technol. 2011, 232, 231–240. [Google Scholar] [CrossRef]
- Li, J.-H.; Li, S.; Zhi, Z.-J.; Yan, L.-F.; Ye, X.-Q.; Ding, T.; Yan, L.; Linhardt, R.J.; Chen, S.-G. Depolymerization of Fucosylated Chondroitin Sulfate with a Modified Fenton-System and Anticoagulant Activity of the Resulting Fragments. Mar. Drugs 2016, 14, E170. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lee, I.W.; Li, J.; Chen, X.; Park, H.J. Fabrication of electrospun antioxidant nanofibers by rutin-pluronic solid dispersions for enhanced solubility. J. Appl. Polym. Sci. 2017, 134. [Google Scholar] [CrossRef]
- Li, C.; Bai, J.H.; Cai, Z.L.; Fan, O.Y. Optimization of a cultural medium for bacteriocin production by Lactococcus lactis using response surface methodology. J. Biotechnol. 2002, 93, 27–34. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
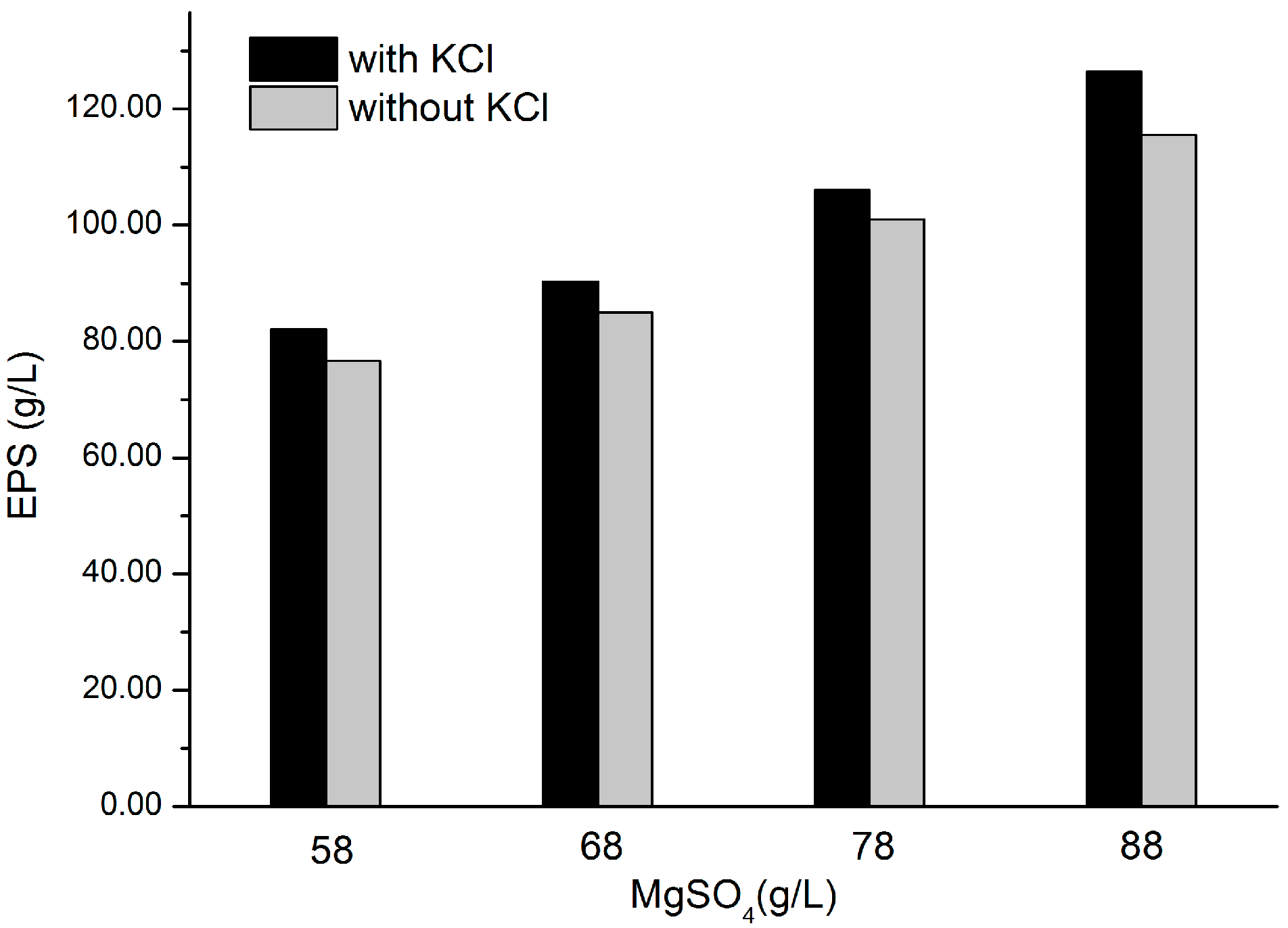{kind=link}
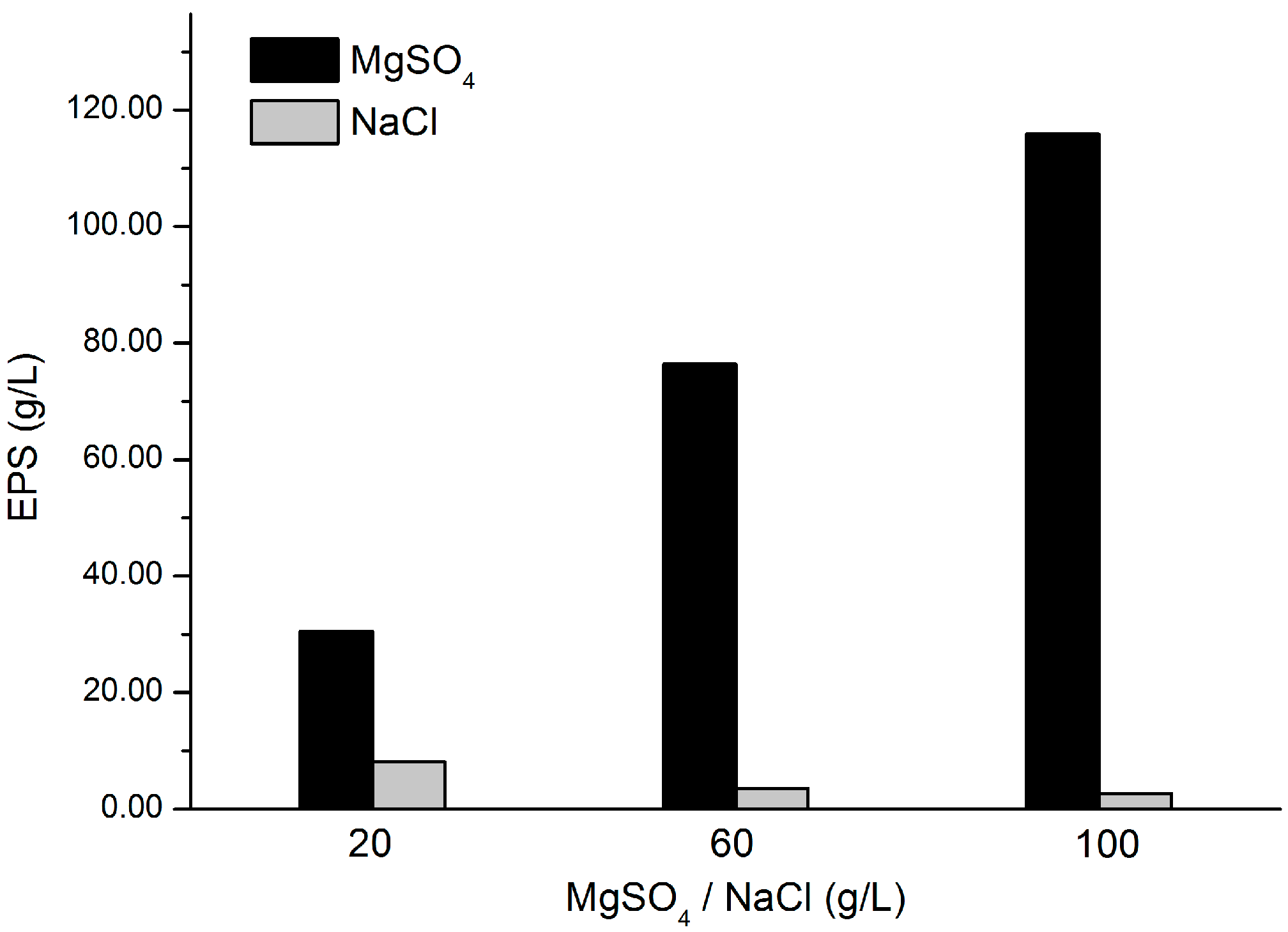{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Formula |
|---|---|
| M1 | 10 g/L peptone, 3 g/L beef extract, 3 g/L sodium citrate, 10 g/L MgSO4, 2 g/L KCl, 0.05 g/L FeSO4·7H2O, 5 g/L NaCl; pH 7.0–7.2. |
| M2 | 70g/L glucose, 10 g/L (NH4)2SO4, 1 g/L yeast extract, 3 g/L KH2PO4, 9 g/L K2HPO4, 0.4 g/L MgSO4·7H2O, 0.01 g/L MnSO4·H2O, 30 g/L NaCl; pH 7.0–7.2. |
| M3 | 5 g/L beef extract, 10 g/L peptone, 5 g/L KCl, 2.5 g/L MgSO4·7H2O; pH 7.0–7.2. |
| M4 | 20 g/L soluble starch, 0.5 g/L NaCl, 1 g/L KNO3, 0.5 g/L KH2PO4·3H2O, 2.5 g/L MgSO4·7H2O, 0.01 g/L FeSO4·7H2O; pH 7.4–7.6. |
| M5 | 5 g/L casein hydrolysate, 10 g/L sodium citrate, 3 g/L yeast extract, 2 g/L KCl, 5 g/L peptone, 10g/L MgSO4; pH 7.0–7.2. |
| Independent Variables | Variable Name | Levels a | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| X1 (g/L) | Casein hydrolysate | 2 | 5 | 8 |
| X2 (g/L) | Sodium citrate | 5 | 10 | 15 |
| X3 (g/L) | Yeast extract | 1 | 3 | 5 |
| X4 (g/L) | KCl | 1 | 2 | 3 |
| X5 (g/L) | Peptone | 2 | 5 | 8 |
| X6 (g/L) | MgSO4 | 20 | 30 | 40 |
| Run | x1 | x2 | x3 | x4 | x5 | x6 | Y (EPS, g/L) |
|---|---|---|---|---|---|---|---|
| 1 | 1 | −1 | 1 | −1 | −1 | 1 | 35.97 |
| 2 | 1 | −1 | −1 | 1 | 1 | 1 | 40.20 |
| 3 | 0 | 0 | 0 | 0 | 0 | 0 | 27.36 |
| 4 | −1 | −1 | −1 | −1 | −1 | −1 | 15.16 |
| 5 | −1 | 1 | 1 | −1 | −1 | −1 | 16.62 |
| 6 | −1 | 1 | 1 | 1 | −1 | 1 | 38.21 |
| 7 | 1 | 1 | −1 | 1 | −1 | −1 | 20.40 |
| 8 | −1 | −1 | 1 | 1 | 1 | −1 | 17.87 |
| 9 | 1 | −1 | −1 | −1 | 1 | −1 | 12.34 |
| 10 | 1 | 1 | −1 | −1 | −1 | 1 | 36.88 |
| 11 | −1 | −1 | −1 | 1 | −1 | 1 | 38.87 |
| 12 | −1 | −1 | 1 | −1 | 1 | 1 | 36.18 |
| 13 | 0 | 0 | 0 | 0 | 0 | 0 | 26.21 |
| 14 | −1 | 1 | −1 | 1 | 1 | −1 | 27.25 |
| 15 | 0 | 0 | 0 | 0 | 0 | 0 | 27.42 |
| 16 | −1 | 1 | −1 | −1 | 1 | 1 | 26.54 |
| 17 | 1 | 1 | 1 | −1 | 1 | −1 | 20.60 |
| 18 | 1 | −1 | 1 | 1 | −1 | −1 | 19.28 |
| 19 | 0 | 0 | 0 | 0 | 0 | 0 | 30.41 |
| 20 | 1 | 1 | 1 | 1 | 1 | 1 | 44.77 |
| Source | Coefficient Estimate | Mean Square | F Value | Prob > F |
|---|---|---|---|---|
| Model | 27.9463 | 126.9735 | 11.9082 | 0.0064 |
| X1 | 0.8588 | 11.7992 | 1.1066 | 0.3410 |
| X2 | 0.9625 | 14.8225 | 1.3901 | 0.2914 |
| X3 | 0.7413 | 8.7912 | 0.8245 | 0.4055 |
| X4 | 2.9100 | 135.4896 | 12.7069 | 0.0161 |
| X5 | 0.2725 | 1.1881 | 0.1114 | 0.7521 |
| X6 | 9.2563 | 1370.8506 | 128.5655 | <0.0001 |
| X1 × X2 | 0.8950 | 12.8164 | 1.2020 | 0.3229 |
| X1 × X3 | 0.6088 | 5.9292 | 0.5561 | 0.4894 |
| X1 × X4 | −0.5525 | 4.8841 | 0.4581 | 0.5286 |
| X1 × X5 | 0.4000 | 2.5600 | 0.2401 | 0.6449 |
| X1 × X6 | 1.3938 | 31.0806 | 2.9149 | 0.1485 |
| X2 × X4 | 0.8388 | 11.2560 | 1.0556 | 0.3513 |
| X2 × X6 | −1.5650 | 39.1876 | 3.6752 | 0.1134 |
| Curvature | 0.4234 | 0.0397 | 0.8499 | |
| Residual | 10.6627 | |||
| Lack of Fit | 22.2676 | 7.6101 | 0.0668 | |
| Pure error | 2.9261 | |||
| Cor total |
| Run | X4 (KCl) | X6 (MgSO4) | EPSs (g/L) |
|---|---|---|---|
| 1 | 2.0 | 30 | 36.27 |
| 2 | 2.5 | 35 | 40.70 |
| 3 | 3.0 | 40 | 51.96 |
| 4 | 3.5 | 45 | 58.05 |
| 5 | 4.0 | 50 | 63.13 |
| 6 | 4.5 | 55 | 58.03 |
| Run | x4 | x6 | Observed EPS a (g/L) | Predicted EPS (g/L) |
|---|---|---|---|---|
| 1 | −1 | −1 | 52.56 | 52.47 |
| 2 | 1 | 1 | 48.01 | 50.39 |
| 3 | 0 | 1.414 | 75.91 | 71.77 |
| 4 | 0 | −1.414 | 76.81 | 75.14 |
| 5 | 1 | −1 | 62.94 | 65.56 |
| 6 | 0 | 0 | 67.34 | 66.47 |
| 7 | 0 | 0 | 45.29 | 43.30 |
| 8 | −1.414 | 0 | 70.71 | 74.45 |
| 9 | 1.414 | 0 | 61.6 | 63.05 |
| 10 | 0 | 0 | 65.38 | 63.05 |
| 11 | 0 | 0 | 63.87 | 63.05 |
| 12 | −1 | 1 | 60.53 | 63.05 |
| 13 | 0 | 0 | 63.86 | 63.05 |
| Source | Sum of Squares | DF | Mean Square | F Value | Prob > F |
|---|---|---|---|---|---|
| Model | 1030.53 | 5 | 206.11 | 21.75 | 0.0004 |
| x4 | 0.83 | 1 | 0.83 | 0.09 | 0.7762 |
| x6 | 970.19 | 1 | 970.19 | 102.37 | <0.0001 |
| x42 | 15.32 | 1 | 15.32 | 1.62 | 0.2442 |
| x62 | 30.27 | 1 | 30.27 | 3.19 | 0.1171 |
| x4 × x6 | 7.43 | 1 | 7.43 | 0.78 | 0.4055 |
| Residual | 66.34 | 7 | 9.48 | ||
| Lack of fit | 51.13 | 3 | 17.04 | 4.48 | 0.0907 |
| Pure error | 15.21 | 4 | 3.80 | ||
| Cor total | 1096.88 | 12 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, D.; Jiao, Y.; Wu, J.; Liu, Z.; Chen, Q. Optimization of EPS Production and Characterization by a Halophilic Bacterium, Kocuria rosea ZJUQH from Chaka Salt Lake with Response Surface Methodology. Molecules 2017, 22, 814. https://doi.org/10.3390/molecules22050814
Gu D, Jiao Y, Wu J, Liu Z, Chen Q. Optimization of EPS Production and Characterization by a Halophilic Bacterium, Kocuria rosea ZJUQH from Chaka Salt Lake with Response Surface Methodology. Molecules. 2017; 22(5):814. https://doi.org/10.3390/molecules22050814
Chicago/Turabian StyleGu, Di, Yingchun Jiao, Jianan Wu, Zhengjie Liu, and Qihe Chen. 2017. "Optimization of EPS Production and Characterization by a Halophilic Bacterium, Kocuria rosea ZJUQH from Chaka Salt Lake with Response Surface Methodology" Molecules 22, no. 5: 814. https://doi.org/10.3390/molecules22050814
APA StyleGu, D., Jiao, Y., Wu, J., Liu, Z., & Chen, Q. (2017). Optimization of EPS Production and Characterization by a Halophilic Bacterium, Kocuria rosea ZJUQH from Chaka Salt Lake with Response Surface Methodology. Molecules, 22(5), 814. https://doi.org/10.3390/molecules22050814