
Anti-Inflammatory Activity of a Polymeric Proanthocyanidin from Serjania schiedeana


 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
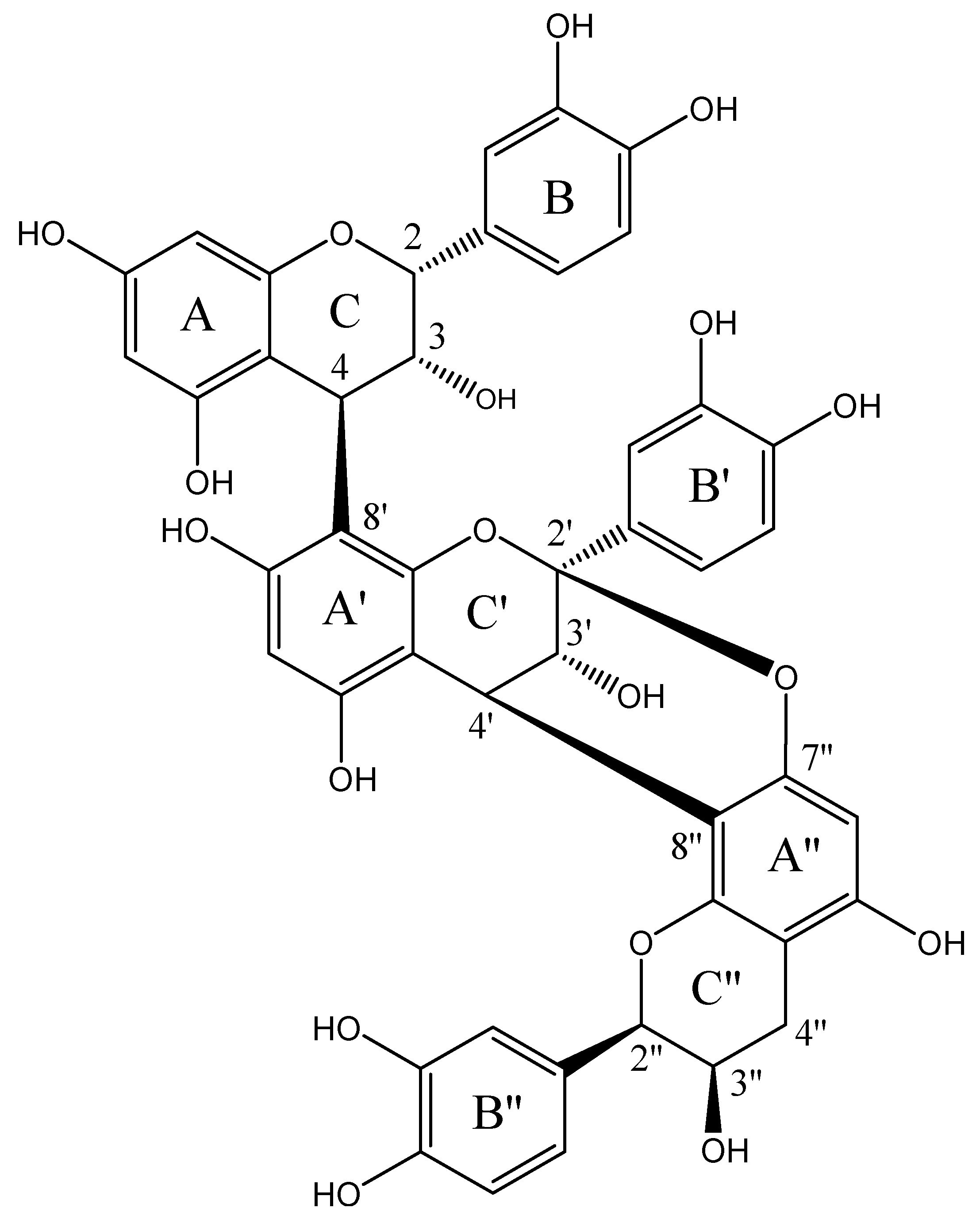
2.1. Structural Elucidation of Epicatechin Trimeric Proanthocyanidin (ETP)
2.2. Anti-Inflammatory Activity of Serjania schiedeana in TPA-Induced Ear Edema
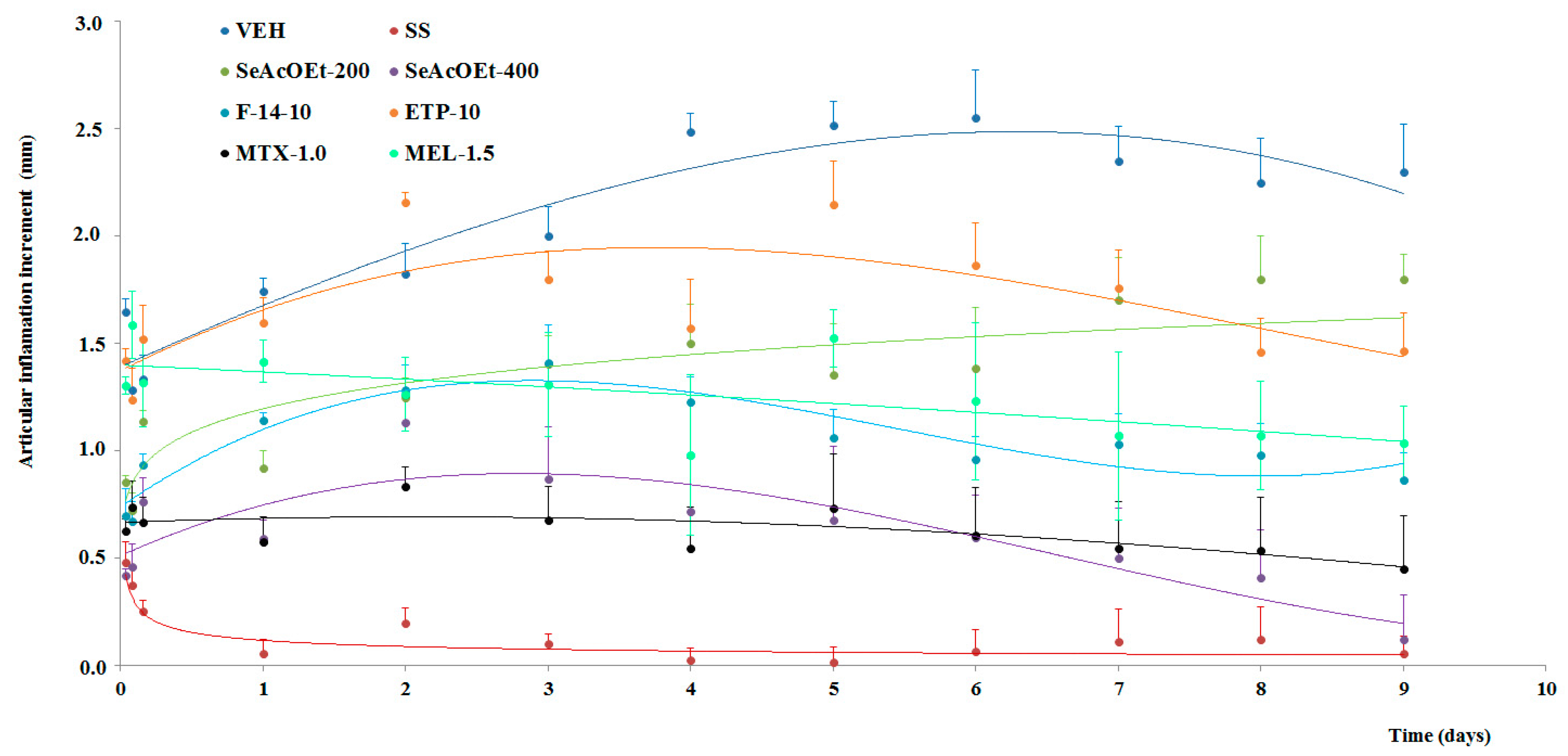
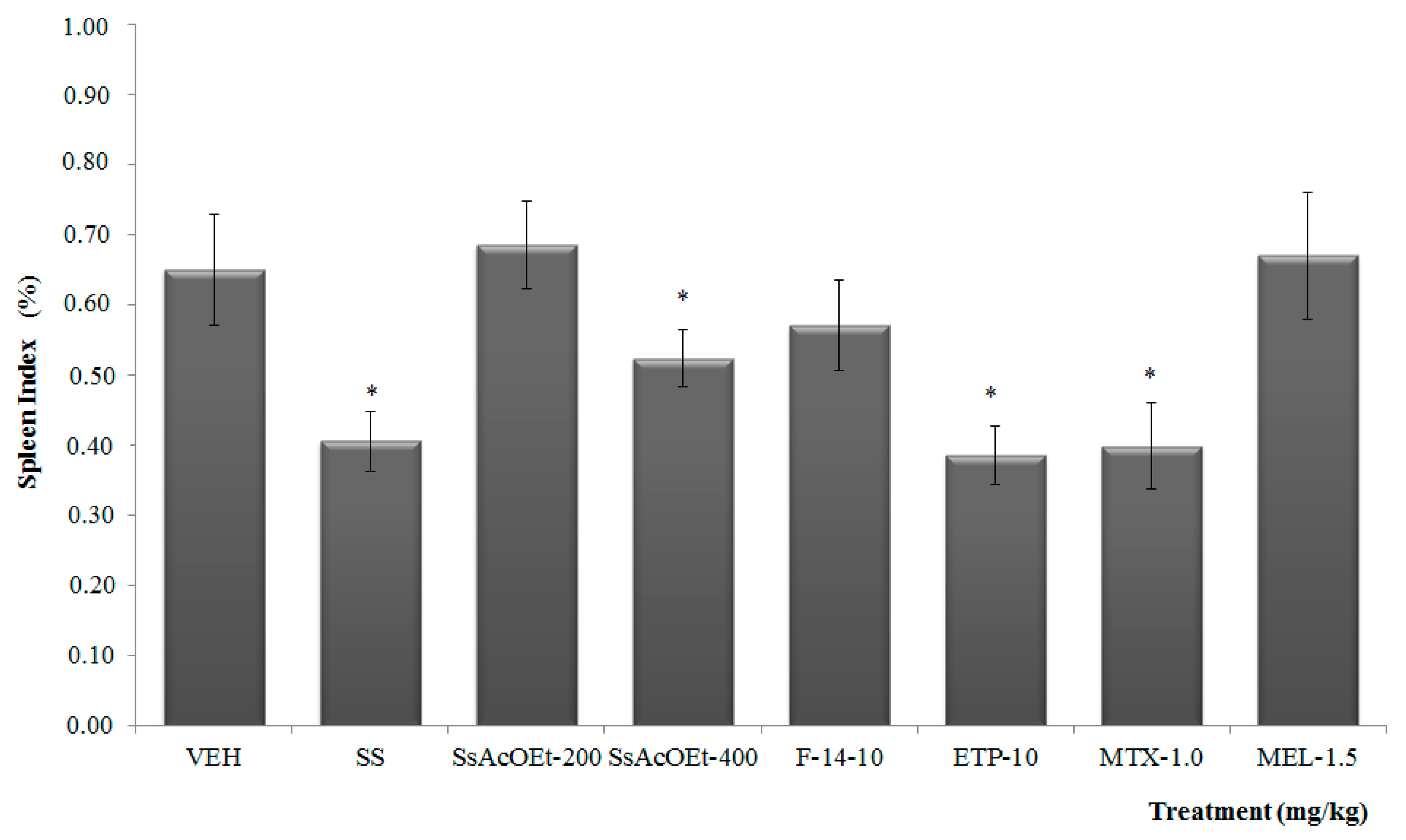
2.3. Anti-Arthritic Activity
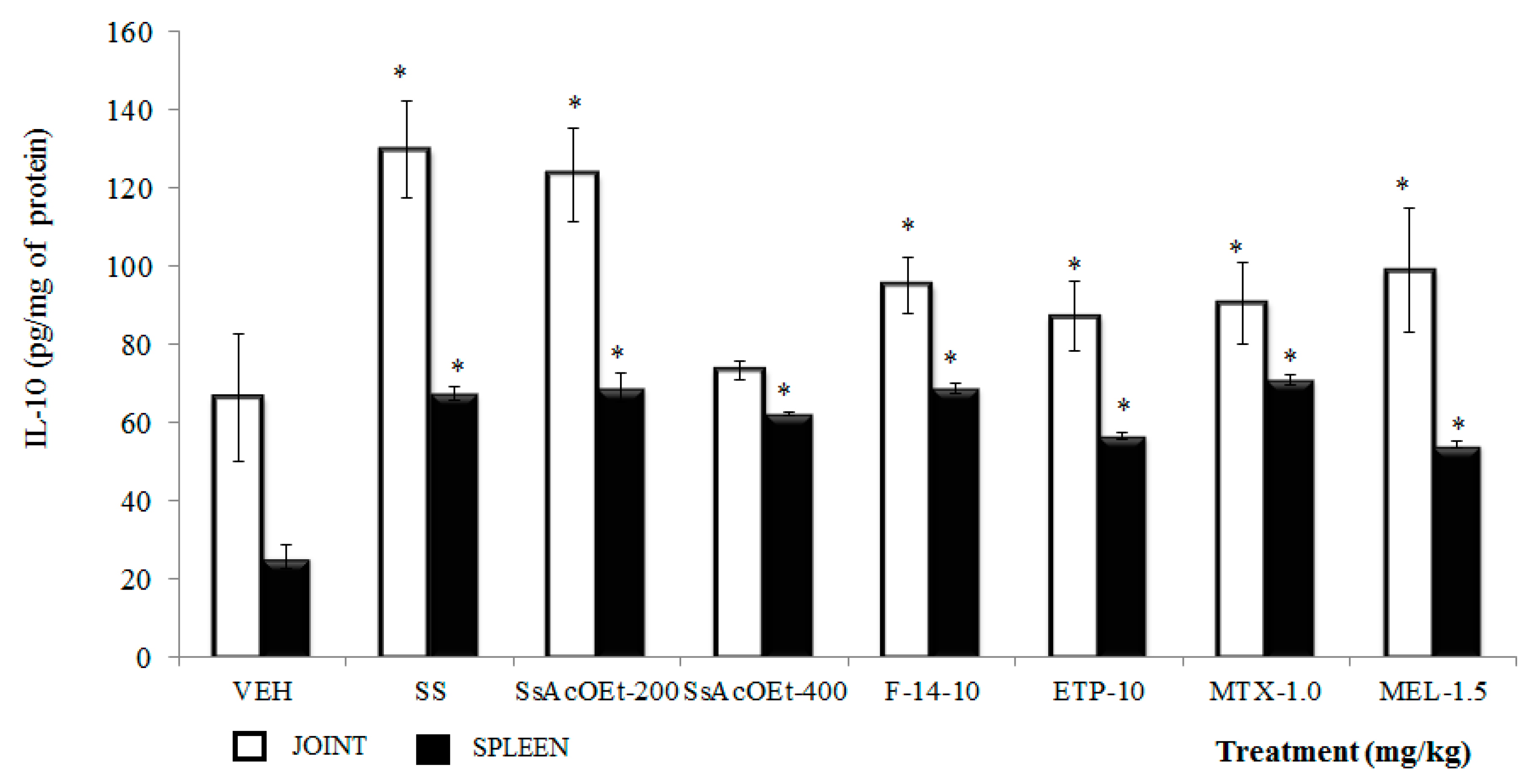
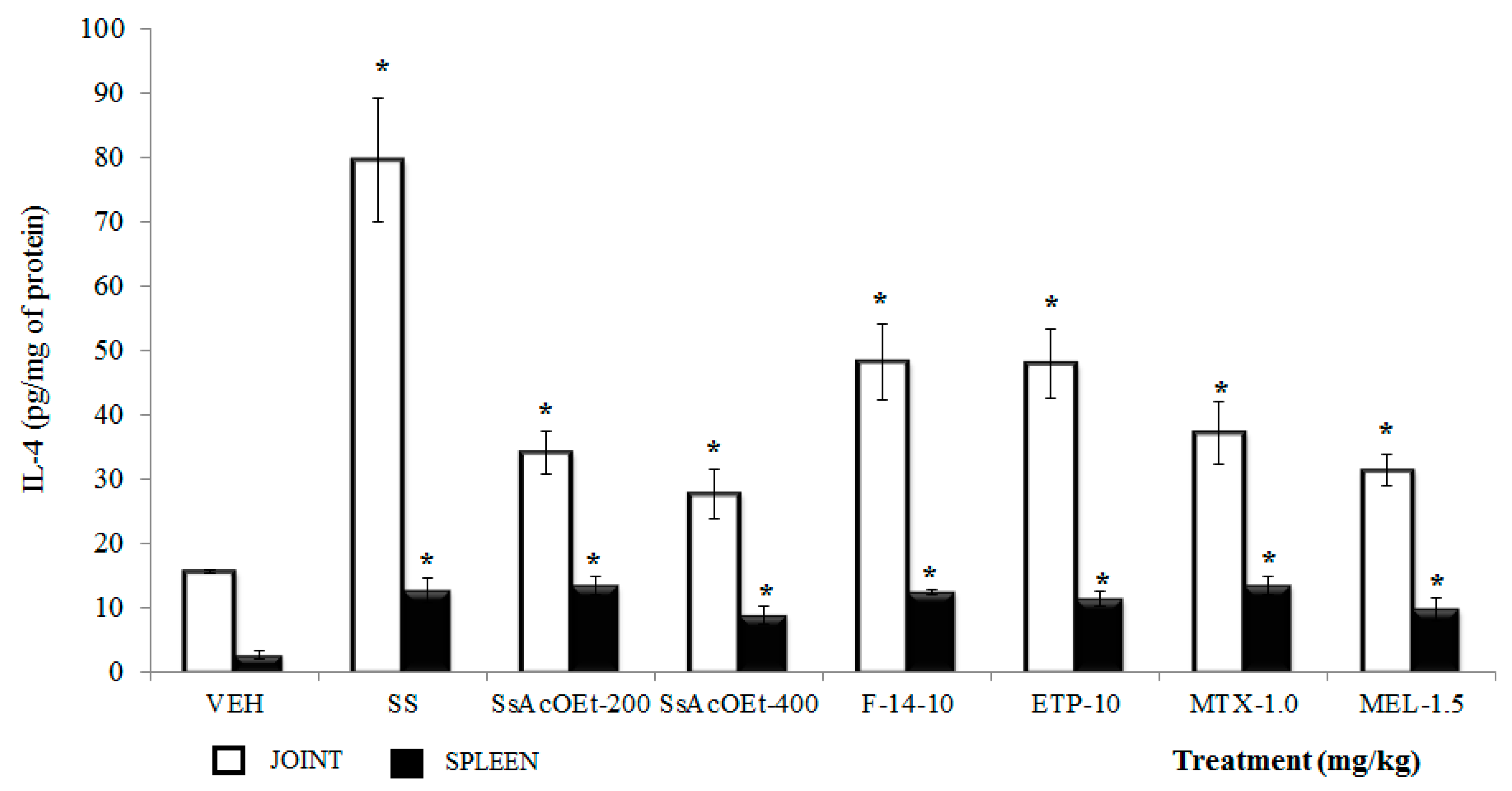
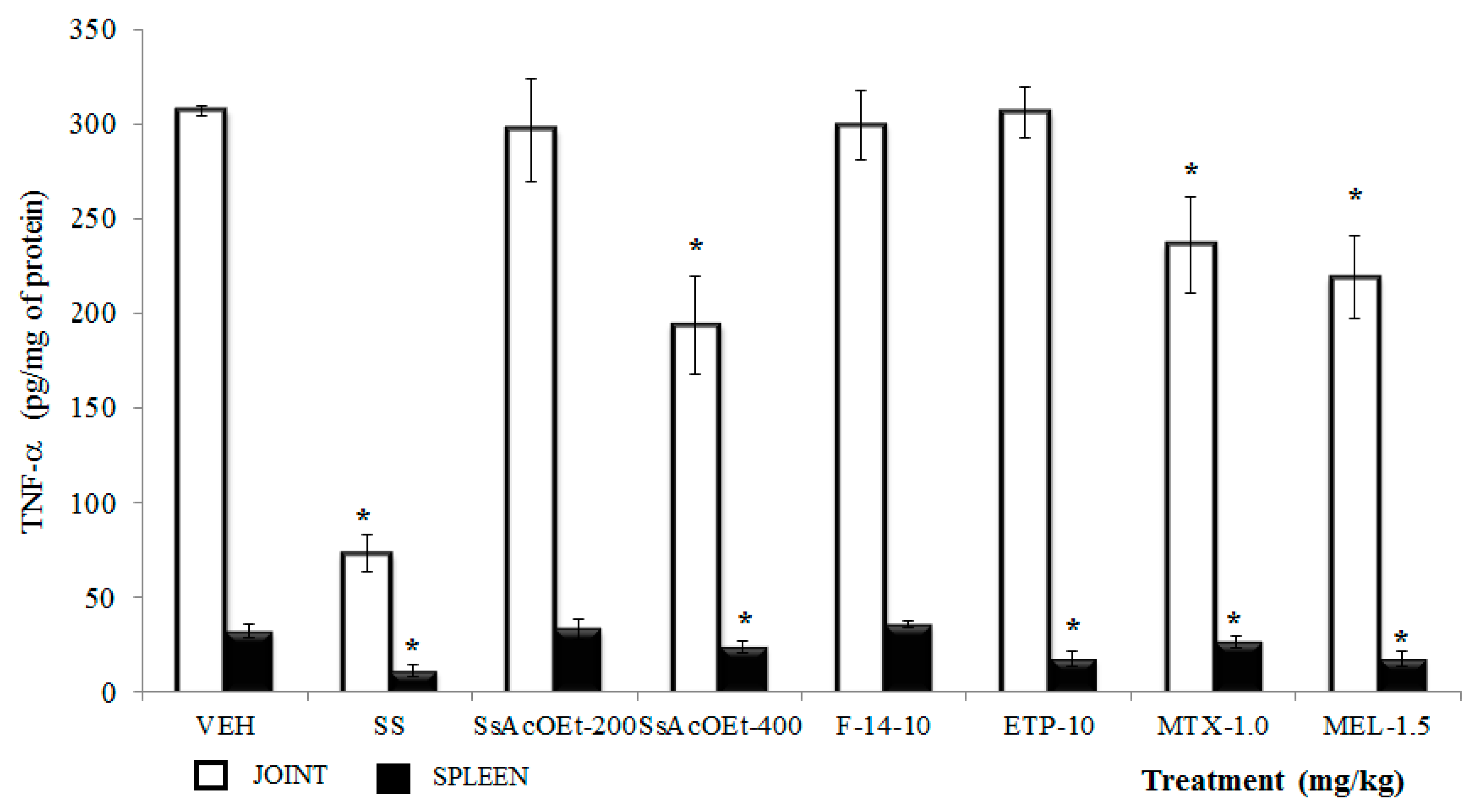
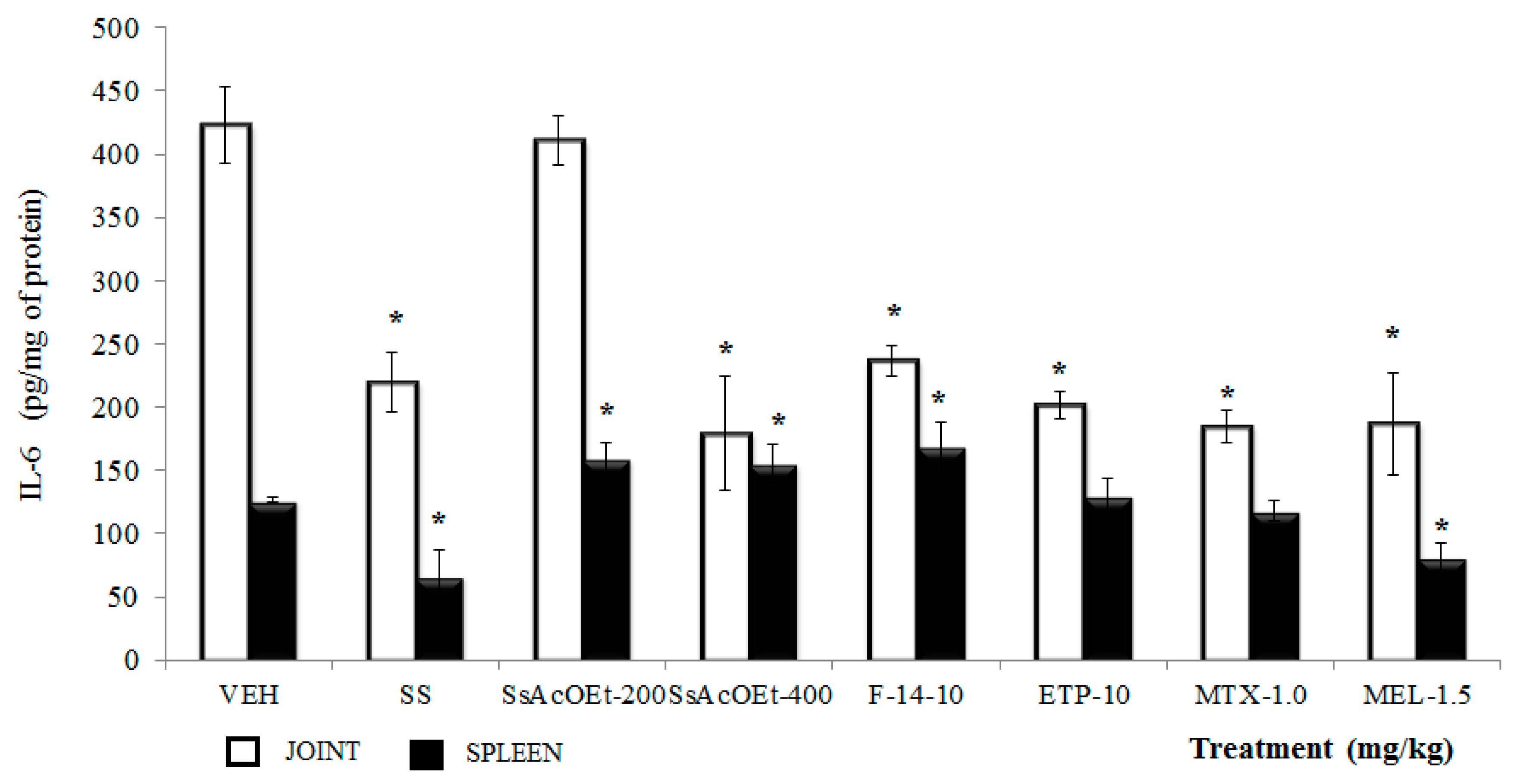
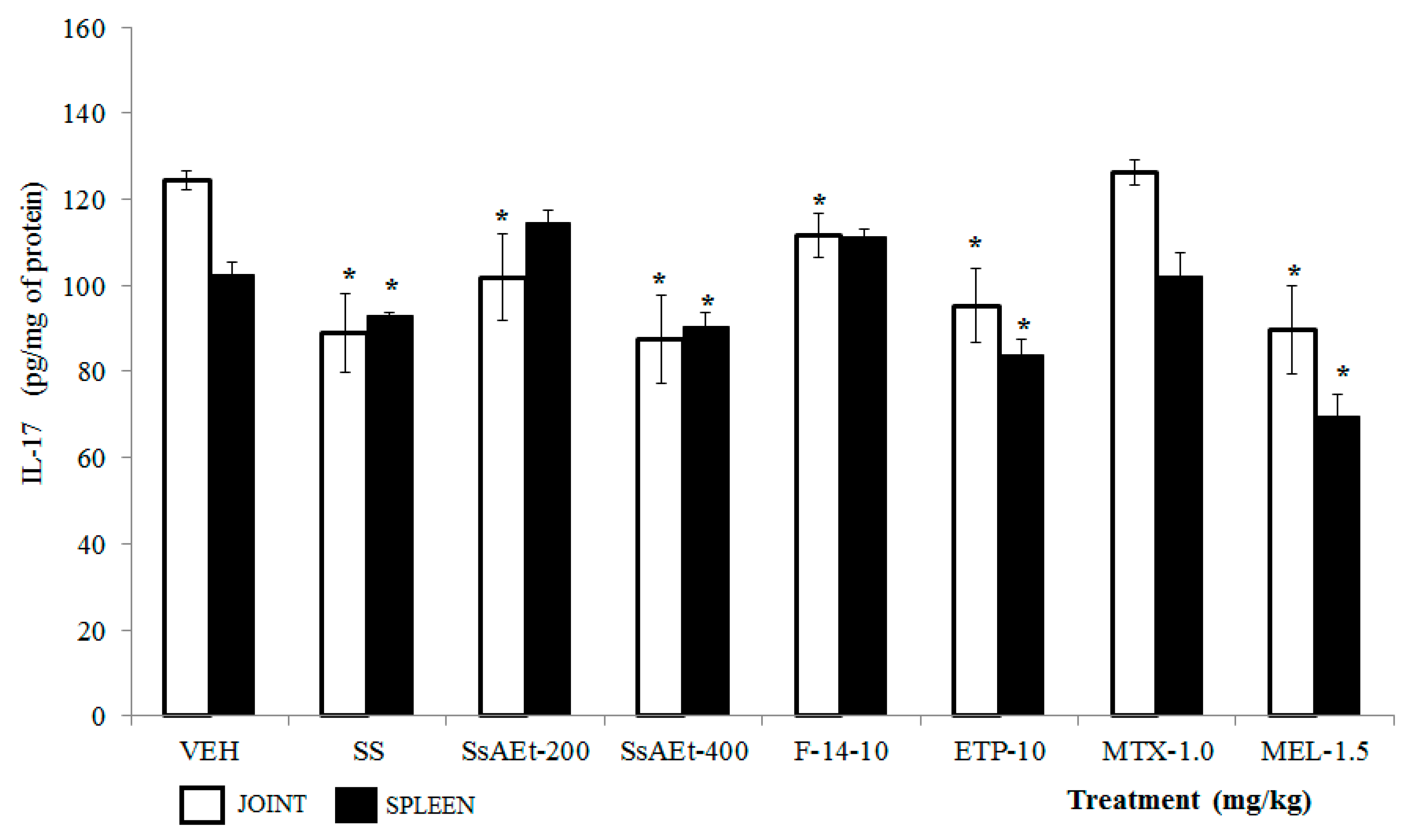
2.4. Effect of S. schiedeana on Local Cytokines
3. Discussion
4. Materials and Methods
4.1. General Information
4.2. Plant Material
4.3. Preparation of Extracts
4.4. Epicatechin Trimeric Proanthocyanidin (ETP) Isolation
4.5. Pharmacological Activity
4.5.1. Animals
4.5.2. Model of Acute Inflammation in Mice with TPA
4.5.3. Arthritis Model Induced by Kaolin/Carrageenan (K/C)
- Group 1: healthy mice (SS)
- Group 2: SsEtOAc at 200 mg/kg (SsEtOAc-200)
- Group 3: SsEtOAc at 400 mg/Kg (SsEtOAc-400)
- Group 4: F-14 at 10 mg/kg (F-14-10)
- Group 5: ETP at 10 mg/kg (ETP-10)
- Group 6: Methotrexate at 1.0 mg/kg (MTX-1.0)
- Group 7: Meloxicam at 1.5 mg/kg (MEL-1.5)
- Group 8: Negative control group (NO treatment, only K/C-VEH-).
4.5.4. Spleen Index
4.5.5. Homogenization of Joint and Spleen Tissues
4.5.6. Quantification of Cytokines IL-1β, IL-6, IL-17, TNF-α, IL-4, and IL-10 by ELISA
4.5.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Humphreys, J.; Hyrich, K.; Symmons, D. What is the impact of biologic therapies on common co-morbidities in patients with rheumatoid arthritis? Arthritis Res. Ther. 2016, 18, 282. [Google Scholar] [CrossRef] [PubMed]
- Chronic Diseases and Health Promotion; World Health Organization: Geneva, Switzerland. Available online: http://www.who.int/chp/topics/rheumatic/en/ (accessed on 29 March 2017).
- Venkateshka, S.H.; Berman, B.M.; Moudgil, K.D. Herbal medicinal products target biochemical and molecular mediators of inflammatory autoimmune arthritis. Rev. Bioorg. Med. Chem. 2011, 19, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.S.; Cronstein, B.N. Methotrexate-how does it really work? Nat. Rev. Rheumatol. 2010, 6, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Díaz-González, F.; Sánchez-Madrid, F. NSAIDs: Learning new tricks from old drugs. Eur. J. Immunol. 2015, 45, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Olsen, N.J.; Murray, L.M. Antiproliferative effects of methotrexate on peripheral blood mononuclear cells. Arthritis Rheum. 1989, 32, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Visser, K.; Katchamart, W.; Loza, E.; Martínez-López, J.A.; Salliot, C.; Trudeau, J.; Bombardier, C.; Carmona, L.; Van der Heijde, D.; Bijlsma, J.W.; et al. Multinational evidence-based recommendations for the use of Methotrexate in rheumatic disorders with a focus on rheumatoid arthritis: Integrating systematic literature research and expert opinion of a broad international panel of Rheumatologists in the 3E Initiative. Ann. Rheum. Dis. 2009, 68, 1086–1089. [Google Scholar] [PubMed]
- Smolen, J.S.; Landewé, R.; Breedveld, F.C.; Dougados, M.; Emery, P.; Gaujoux-Viala, C.; Gorter, S.; Knevel, R.; Nam, J.; Schoels, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs. Ann. Rheum. Dis. 2010, 69, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Fernández de Caleya, P.B.; Espejo-Serna, M.A.; López-Ferrari, A.R. Herbario de la Real Expedición Botánica de Nueva España (1787–1803); Consejo Superior de Investigaciones Científicas (CSIC) Real Jardín Botánico: Madrid, Spain, 2010. [Google Scholar]
- Gomig, F.; Pietrovski, E.F.; Guedes, A.; Dalmarco, E.M.; Calderari, M.T.; Guimarães, C.L.; Pinheiro, R.M.; Cabrini, D.A.; Otuki, M.F. Topical anti-inflammatory activity of Serjania erecta Radlk (Sapindaceae) extracts. J. Ethnopharmacol. 2008, 118, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ye, R.; Yang, Y.; Ma, C. Structures, antioxidant and intestinal disaccharidase inhibitory activities of A-type proanthocyanidins from peanut skin. J. Agric. Food Chem. 2013. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Nonaka, G.I.; Nishioka, I. Tannins and Related Compounds. XXXVIII. Isolation and characterization of flavan-3-ol glucosides and procyanidin oligomers from Cassia bark (Cinnamomum cassia Blume). Chem. Pharm. Bull. 1986, 34, 633–642. [Google Scholar] [CrossRef]
- Morimoto, S.; Nonaka, G.I.; Nishioka, I. Tannins and related Compounds. LX. Isolation and characterization of proanthocyanidins with a doubly-linked unit from Vaccinium vitis-idaea L. Chem. Pharm. Bull. 1988, 36, 33–38. [Google Scholar] [CrossRef]
- Krueger, C.G.; Chesmore, N.; Chen, X.; Parker, J.; Khoo, C.; Marais, J.P.J.; Shanmuganayagam, D.; Crump, P.; Reed, J.D. Critical reevaluation of the 4-(dimethyamino) cinnamaldehyde assay: Cranberry proanthocyanidin standard is superior to proanthocyanidins in cranberry products. J. Funct. Foods 2016, 22, 13–19. [Google Scholar] [CrossRef]
- Feliciano, R.P.; Meudt, J.J.; Shanmuganayagam, D.; Krueger, C.G.; Reed, J.D. Ratio of “A-type” to”B-type” Proanthocyanidin Interflavan Bonds Affects Extra-intestinal Pathogenic Escherichia coli Invasion of Gut Ephithelial Cells. J. Agric. Food Chem. 2014, 62, 3919–3925. [Google Scholar] [CrossRef] [PubMed]
- Neugebauer, V.; Haans, J.S.; Adwanikar, H.; Fu, Y.; Ji, G. Techniques for assessing knee joint pain in arthritis. Mol. Pain 2007, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Simpson, B.; Claudie, D.; Smith, N.; Wang, J.; McKinnon, R.; Semple, S. Evaluation of the anti-inflammatory properties of Dodonaea polyandra, a Kaanju traditional medicine. J. Ethnopharmacol. 2010, 132, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Simpson, B.S.; Claudie, D.J.; Gerber, J.P.; Pyke, S.M.; Wang, J.; McKinnon, R.A.; Semple, S.J. In vivo activity of benzoyl ester clerodane diterpenoid derivatives from Dodonaea polyandra. J. Nat. Prod. 2011, 74, 650–700. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Sánchez, D.O.; Herrera-Ruiz, M.; Pérez, S.; Jiménez-Ferrer, E.; Zamilpa, A. Anti-inflammatory activity of hautriwaic acid isolated from Dodonaea viscosa leaves. Molecules 2012, 17, 4292–4299. [Google Scholar] [CrossRef] [PubMed]
- Salinas-Sánchez, D.O.; Herrera-Ruiz, M.; Pérez, S.; Jiménez-Ferrer, E.; Zamilpa, A.; Tortoriello, J.; González-Cortázar, M. Effect of hautriwaic acid isolated from Dodonaea viscosa in a model of kaolin/carrageenan-induced mono-arthritis. Plant Med. 2015, 21, 1240–1247. [Google Scholar]
- Huang, M.T.; Liu, Y.; Ramji, D.; Lo, C.Y.; Ghai, G.; Dushenkov, S.; Ho, C.T. Inhibitory effects of black tea theaflavin derivatives on 12-O-tetradecanoylphorbol-13-acetate-induced inflammation and arachidonic acid metabolism in mouse ears. Mol. Nutr. Food Res. 2006, 50, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Katsukawa, M.; Nakano, A.; Hiraki, E.; Nishimura, K.; Jisaka, M.; Yokota, K.; Ueda, N. Arachidonate 12-lipoxygenases with reference to their selective inhibitors. Biochem. Biophys. Res. Commun. 2005, 338, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Shaohua, X.; Houshan, L.; Jianhao, L.; Zhankun, C.; Dongfang, J. Regulation of TNFα and IL1-β in rheumatoid arthritis synovial fibroblasts by leukotriene B4. Rheumatol. Int. 2010, 30, 1183–1189. [Google Scholar]
- Vervoordeldonk, M.J.B.M.; Tak, P.P. Cytokines in rheumatoid arthritis. Curr. Rheumatol. Rep. 2002, 4, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Maini, R.N.; Elliott, M.J.; Brennan, F.M.; Williams, R.O.; Chu, C.Q.; Paleolog, E.; Charles, P.J.; Taylor, P.C.; Feldmann, M. Monoclonal anti-TNF alpha antibody as a probe of pathogenesis and therapy of rheumatoid disease. Immunol. Rev. 1995, 144, 195–223. [Google Scholar] [CrossRef] [PubMed]
- Isomäki, P.; Punnonen, J. Pro- and anti-inflammatory cytokines in rheumatoid arthritis. Ann. Med. (Helsinki) 1997, 29, 499–507. [Google Scholar] [CrossRef]
- Smolen, J.S.; Redlich, K.; Zwerina, J.; Aletaha, D.; Steine, G.; Schett, G. Pro-inflammatory cytokines in rheumatoid arthritis. Clin. Rev. Allergy Immunol. 2005, 28, 239–248. [Google Scholar] [CrossRef]
- Kimura, A.; Kishimoto, T. IL-6: Regulator of Treg/Th17 balance. Eur. J. Immunol. 2010, 40, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.C.; Symons, J.A.; Dickens, E.; Duff, G.W. In situ hybridization of IL-6 in rheumatoid arthritis. Clin. Exp. Immunol. 1992, 87, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Matsuda, T.; Turner, M.; Miyasaka, N.; Buchan, G.; Tang, B.; Satoa, K.; Shimid, M.; Maid, R.; Feldmann, M.; Kishimoto, T. Excessive production of interleukin 6/B cell stimulatory factor-2 in rheumatoid arthritis. Eur. J. Immunol. 1988, 18, 1797–1801. [Google Scholar] [CrossRef] [PubMed]
- Katayama, M.; Ohmura, K.; Yukawa, N.; Terao, C.; Hashimoto, M. Neutrophils are essential as a source of IL-17 in the effector phase of arthritis. PLoS ONE 2013, 8, e62231. [Google Scholar] [CrossRef]
- Ye, L.; Wen, Z.; Li, Y.; Chen, B.; Ting, Y.; Liu, L.; Zhang, J.; Ma, Y.; Xiao, S.; Ding, L.; Li, L.; Huang, Z. Interleukin-10 attenuation of collagen-induced arthritis is associated with suppression of interleukin-17 and retinoid-related orphan receptor γt production in macrophages and repression of classically activated macrophages. Arthritis Res. Ther. 2014, 16, R96. [Google Scholar] [PubMed]
- Venkatesha, S.H.; Dudics, S.; Acharya, B.; Moudgil, K.D. Cytokine-modulating strategies and newer cytokine targets for arthritis therapy. Int. J. Mol. Sci. 2015, 16, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.K.; O’Donnell, K.; Lawlor, K.E.; Wicks, I.P. Severe inflammatory arthritis and lymphadenopathy in the absence of TNF. J. Clin. Investig. 2001, 107, 1519–1527. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Umar, S.; Riegsecker, S.; Chourasia, M.; Ahmed, S. Regulation of transforming growth factor β-activated kinase activation by epigallocatechin-3-gallate in rheumatoid arthritis synovial fibroblasts: Suppression of K(63)-linked autoubiquitination of tumor necrosis factor receptor-associated factor 6. Arthritis Rheumatol. 2016, 68, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Payá, M.; Ferrándiz, M.; Sánz, M.J.; Bustos, G.; Blasco, R.; Ríos, J.L.; Alcaráz, M.J. Study of the antioedema activity of some seaweed and sponge extracts from the Mediterranean coast in mice. Phytother. Res. 1993, 7, 159–162. [Google Scholar] [CrossRef]
Sample Availability: Samples of compound: epicatechin-(4β → 8)–epicatechin–(4β → 8, 2β → O → 7) epicatechin, it is not available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ring | No. | 13C | 1H | Ring | No. | 13C | 1H | Ring | No | 13C | 1H |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | 2 | 79.01 | 5.7, s, m | C′ | 2′ | 105.08 | 3.28 (d, 3.47) 4.15 (d, 3.47) | C″ | 2″ | 80.44 | 4.39 (s) |
| 3 | 72.72 | 4.13, d, m | 3′ | 67.33 | 3.28 (d, 3.47) | 3″ | 67.67 | 3.87–3.85 (m) | |||
| 4 | 38.92 | 4.6, br, s | 4′ | 29.01 | 4.15, (d, 3.47) | 4″ | 30 | a. 2.84 (dd, 4.6, 17.3) b. 2.84 (dd, 4.6, 17.3) | |||
| A | 5 | 156.90 | A′ | 5′ | 151.2 | A″ | 5″ | 156.16 | |||
| 6 | 98.9 | 6.02 (d, 2.3) | 6′ | 96.21 | 5.8 (s) | 6″ | 96.61 | 6.10 (s) | |||
| 7 | 157.95 | 7′ | 155.9 | 7″ | 155.94 | ||||||
| 8 | 96.7 | 5.98 (d, 2.3) | 8′ | 106.87 | 8″ | 108.9 | |||||
| 9 | 154.29 | 9′ | 151.92 | 9″ | 151.92 | ||||||
| 10 | 106.55 | 10′ | 100.09 | 10″ | 100.19 | ||||||
| B | 11 | 131 | B′ | 11′ | 119.56 | B″ | 11″ | 133.3 | |||
| 12 | 116.86 | 7.32 d (1.9) | 12′ | 115.88 | 6.10 d (1.5) | 12″ | 115.85 | 7.03 (d, 1.93) | |||
| 13 | 145.89 | 13′ | 145.46 | 13″ | 146.74 | ||||||
| 14 | 146.42 | 14′ | 145.89 | 14″ | 146.03 | ||||||
| 15 | 120.01 | 6.83 (d, 8.0) | 15′ | 116.14 | 6.83 (d, 8.0) | 15″ | 116.27 | 6.75 (d, 8.0) | |||
| 16 | 121.49 | 7.2 (dd, 1.9, 8) | 16′ | 115.61 | 7.2 (dd, 1.9, 8) | 16″ | 119.56 | 6.73 (dd, 1.93, 8.04) |
| Treatment | Doses (mg/Ear) | Edema (mg) Mean ± SEM | Edema Inhibition (%) | Pharmacologic Parameters |
|---|---|---|---|---|
| SsEtOAc | 2.0 | 1.02 ± 0.71 * | 90.09 | |
| F-6 | 1.0 | 2.94 ± 0.34 * | 71.68 | |
| F-12 | 2.24 ± 0.70 * | 78.42 | ||
| F-13 | 3.40 ± 1.0 * | 67.24 | ||
| F-14 | 1.04 ± 0.22 * | 89.95 | ||
| ETP | 0.125 | 7.9 ± 1.01 | 23.5 | ED50 = 0.25 mg/ear Emax = 52.9% |
| 0.25 | 7.6 ± 0.73 | 26.46 | ||
| 0.5 | 7.5 ± 0.89 | 27.74 | ||
| 1.0 | 5.5 ± 1.39 * | 47.01 | ||
| 2.0 | 2.8 ± 0.93 * | 72.22 | ||
| IND | 1.0 | 1.2 ± 0.4 * | 88.25 | |
| VEH | ---- | 10.3 ±1.46 | ---- |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salinas-Sánchez, D.O.; Jiménez-Ferrer, E.; Sánchez-Sánchez, V.; Zamilpa, A.; González-Cortazar, M.; Tortoriello, J.; Herrera-Ruiz, M. Anti-Inflammatory Activity of a Polymeric Proanthocyanidin from Serjania schiedeana. Molecules 2017, 22, 863. https://doi.org/10.3390/molecules22060863
Salinas-Sánchez DO, Jiménez-Ferrer E, Sánchez-Sánchez V, Zamilpa A, González-Cortazar M, Tortoriello J, Herrera-Ruiz M. Anti-Inflammatory Activity of a Polymeric Proanthocyanidin from Serjania schiedeana. Molecules. 2017; 22(6):863. https://doi.org/10.3390/molecules22060863
Chicago/Turabian StyleSalinas-Sánchez, David Osvaldo, Enrique Jiménez-Ferrer, Veronica Sánchez-Sánchez, Alejandro Zamilpa, Manasés González-Cortazar, Jaime Tortoriello, and Maribel Herrera-Ruiz. 2017. "Anti-Inflammatory Activity of a Polymeric Proanthocyanidin from Serjania schiedeana" Molecules 22, no. 6: 863. https://doi.org/10.3390/molecules22060863
APA StyleSalinas-Sánchez, D. O., Jiménez-Ferrer, E., Sánchez-Sánchez, V., Zamilpa, A., González-Cortazar, M., Tortoriello, J., & Herrera-Ruiz, M. (2017). Anti-Inflammatory Activity of a Polymeric Proanthocyanidin from Serjania schiedeana. Molecules, 22(6), 863. https://doi.org/10.3390/molecules22060863