
Production of Putative Diterpene Carboxylic Acid Intermediates of Triptolide in Yeast

 ,
,
Abstract
:1. Introduction
2. Results
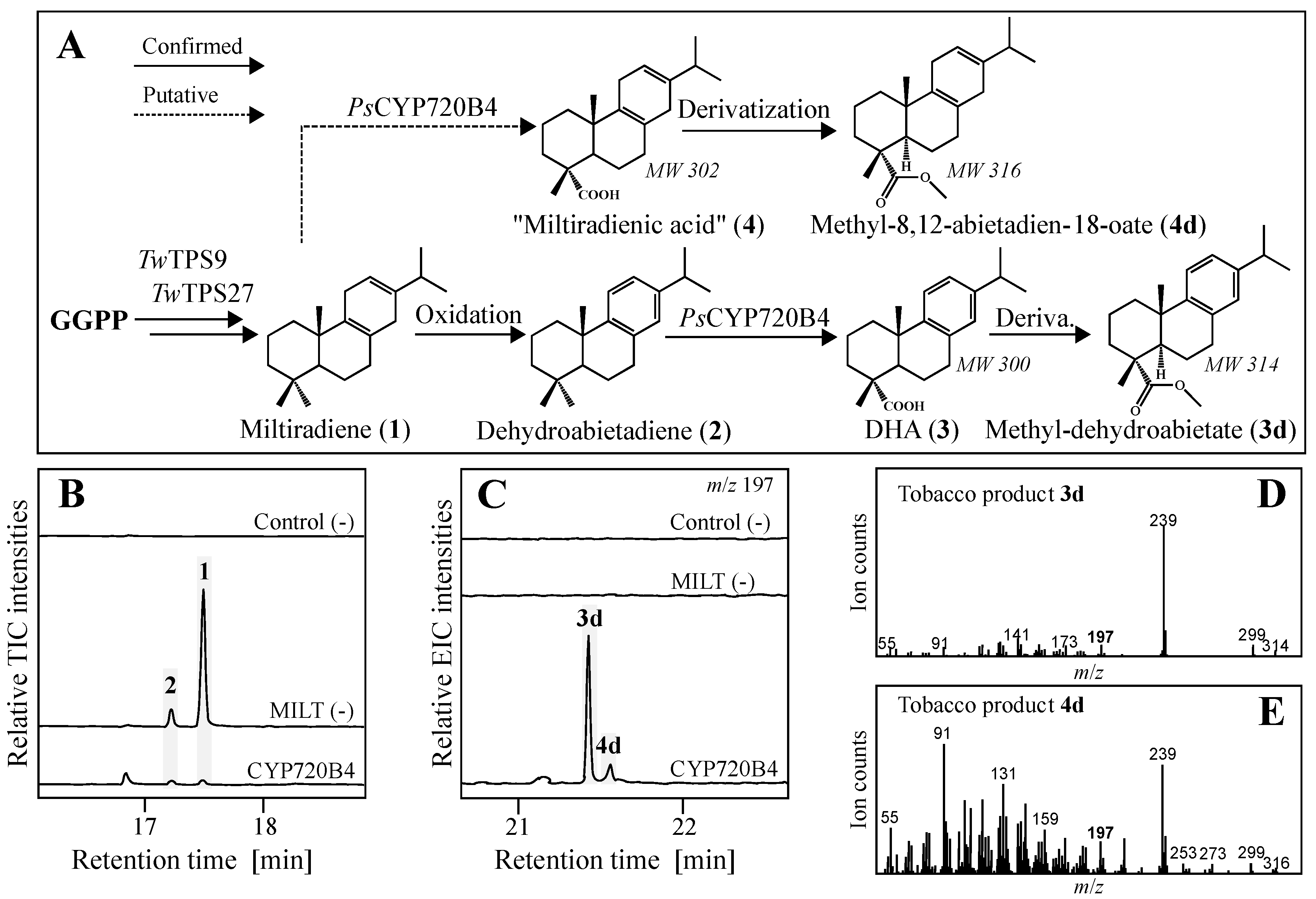
2.1. Identification of a Biosynthetic Pathway to Dehydroabietic Acid by Transient Expression in Tobacco
2.2. Production of Diterpene Backbones in Yeast
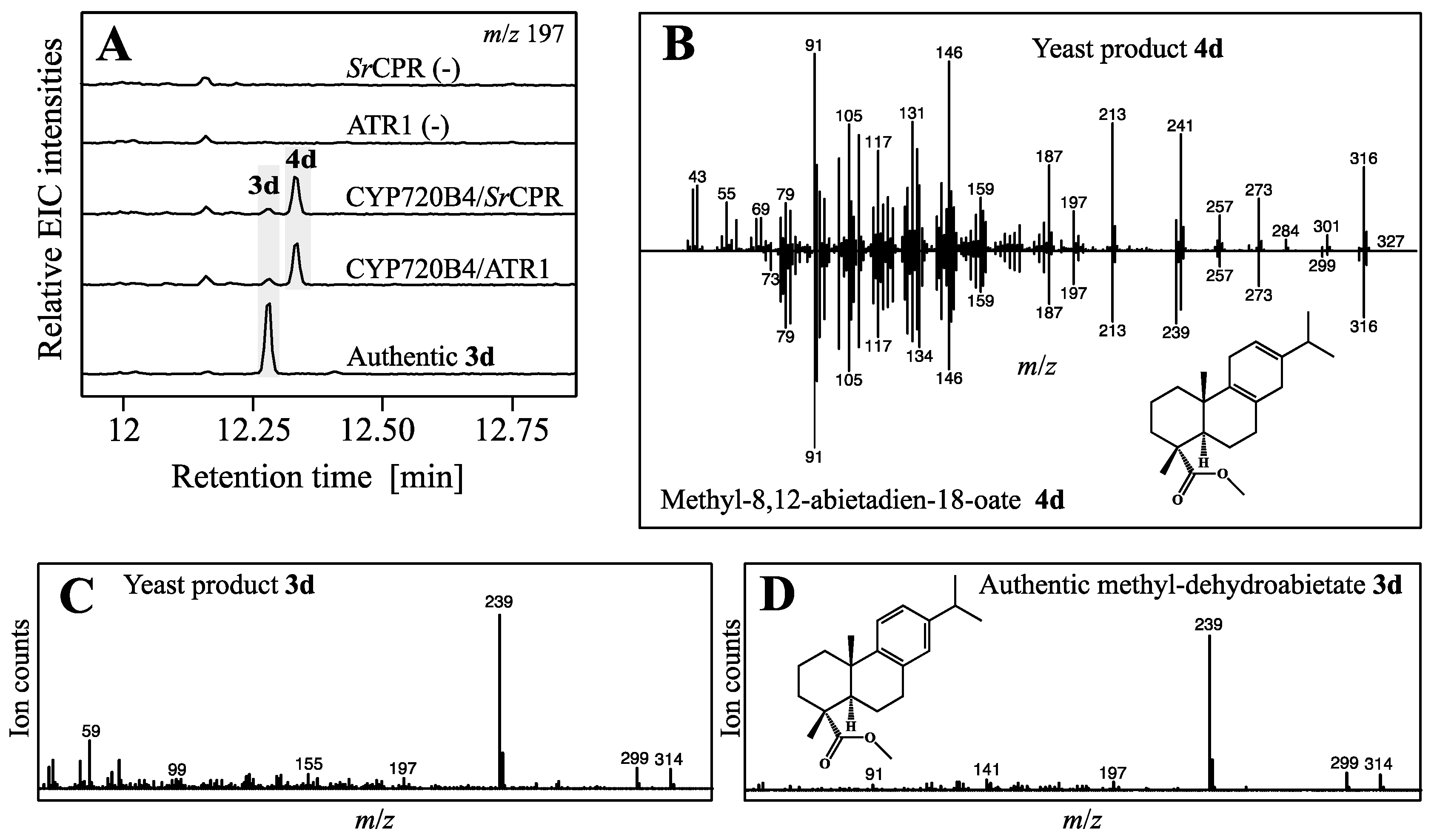
2.3. Yeast In Vivo Production of Dehydroabietic Acid and Novel Acid Form of Miltiradiene
3. Discussion
4. Materials and Methods
4.1. Transient in Planta Expression in Tobacco
4.2. Cloning and Construction of Yeast Plasmids
4.3. Yeast Diterpene Production Procedure
4.4. Yeast Extraction Procedures
4.5. Analytical method
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Peters, R.J. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Wagh, V.; Patil, P.; Surana, S.; Wagh, K. Forskolin: Upcoming antiglaucoma molecule. J. Postgrad. Med. 2012, 58, 199. [Google Scholar] [CrossRef] [PubMed]
- Barrero, A.F.; Alvarez-Manzaneda, E.J.; Altarejos, J.; Salido, S.; Ramos, J.M. Synthesis of Ambrox® from (−)-sclareol and (+)-cis-abienol. Tetrahedron 1993, 49, 10405–10412. [Google Scholar] [CrossRef]
- Zhou, Z.-L.; Yang, Y.-X.; Ding, J.; Li, Y.-C.; Miao, Z.-H. Triptolide: Structural modifications, structure-activity relationships, bioactivities, clinical development and mechanisms. Nat. Prod. Rep. 2012, 29, 457. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Salnikov, A.V.; Bauer, N.; Aleksandrowicz, E.; Labsch, S.; Nwaeburu, C.; Mattern, J.; Gladkich, J.; Schemmer, P.; Werner, J.; et al. Triptolide reverses hypoxia-induced epithelial-mesenchymal transition and stem-like features in pancreatic cancer by NF-κB downregulation. Int. J. Cancer 2014, 134, 2489–2503. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Chen, J.; Guo, Z.; Xu, X.-M.; Wang, L.; Pei, X.-F.; Yang, J.; Underhill, C.B.; Zhang, L. Triptolide inhibits the growth and metastasis of solid tumors. Mol. Cancer Ther. 2003, 2, 65–72. [Google Scholar] [PubMed]
- Xu, F.; Li, Y.; Li, S.; Ma, Y.; Zhao, N.; Liu, Y.; Qian, N.; Zhao, H.; Li, Y. Complete Freund’s adjuvant-induced acute inflammatory pain could be attenuated by triptolide via inhibiting spinal glia activation in rats. J. Surg. Res. 2014, 188, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; LeBlanc, K.J.; Li, L. Triptolide preserves cognitive function and reduces neuropathology in a mouse model of Alzheimer’s disease. PLoS ONE 2014, 9, e108845. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.M.; Raskin, I. Determination of triptolide in root extracts of Tripterygium wilfordii by solid-phase extraction and reverse-phase high-performance liquid chromatography. J. Chromatogr. A 2005, 1070, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Tang, H.; Feng, H.; Li, Y. Metal-mediate reactions based formal synthesis of triptonide and triptolide. Tetrahedron Lett. 2014, 55, 7118–7120. [Google Scholar] [CrossRef]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.D.; Tai, A.; Main, A.; Eng, D.; et al. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528–532. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Callari, R.; Hamberger, B.; Wubshet, S.G.; Nielsen, M.T.; Andersen-Ranberg, J.; Hallström, B.M.; Cozzi, F.; Heider, H.; Lindberg Møller, B.; et al. Oxidation and cyclization of casbene in the biosynthesis of Euphorbia factors from mature seeds of Euphorbia lathyris L. Proc. Natl. Acad. Sci. USA 2016. [Google Scholar] [CrossRef] [PubMed]
- Pateraki, I.; Andersen-ranberg, J.; Jensen, N.B.; Wubshet, S.G.; Heskes, A.M.; Forman, V.; Hallström, B.; Hamberger, B.; Motawia, M.S.; Olsen, C.E.; et al. Total biosynthesis of the cyclic AMP booster forskolin from Coleus forskohlii. Elife 2017, 6, e23001. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Zerbe, P.; Hamberger, B.; Yuen, M.M.S.; Chiang, A.; Sandhu, H.K.; Madilao, L.L.; Nguyen, A.; Hamberger, B.; Bach, S.S.; Bohlmann, J. Gene discovery of modular diterpene metabolism in nonmodel systems. Plant Physiol. 2013, 162, 1073–1091. [Google Scholar] [CrossRef] [PubMed]
- Hamberger, B.; Bak, S. Plant P450s as versatile drivers for evolution of species-specific chemical diversity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120426. [Google Scholar] [CrossRef] [PubMed]
- Pateraki, I.; Andersen-Ranberg, J.; Hamberger, B.; Heskes, A.M.; Martens, H.J.; Zerbe, P.; Bach, S.S.; Møller, B.L.; Bohlmann, J.; Hamberger, B. Manoyl oxide (13R), the biosynthetic precursor of forskolin, is synthesized in specialized root cork cells in Coleus forskohlii. Plant Physiol. 2014, 164, 1222–1236. [Google Scholar] [CrossRef] [PubMed]
- Ro, D.-K.; Arimura, G.-I.; Lau, S.Y.W.; Piers, E.; Bohlmann, J. Loblolly pine abietadienol/abietadienal oxidase PtAO (CYP720B1) is a multifunctional, multisubstrate cytochrome P450 monooxygenase. Proc. Natl. Acad. Sci. USA 2005, 102, 8060–8065. [Google Scholar] [CrossRef] [PubMed]
- Zi, J.; Peters, R.J. Characterization of CYP76AH4 clarifies phenolic diterpenoid biosynthesis in the Lamiaceae. Org. Biomol. Chem. 2013, 11, 7650–7652. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.L.; Heskes, A.M.; Hamberger, B.; Olsen, C.E.; Hallström, B.M.; Andersen-Ranberg, J.; Hamberger, B. The terpene synthase gene family in Tripterygium wilfordii harbors a labdane-type diterpene synthase among the monoterpene synthase TPS-b subfamily. Plant J. 2017, 89, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Brinker, A.M.; Ma, J.; Lipsky, P.E.; Raskin, I. Medicinal chemistry and pharmacology of genus Tripterygium (Celastraceae). Phytochemistry 2007, 68, 732–766. [Google Scholar] [CrossRef] [PubMed]
- Kutney, J.P.; Han, K. Studies with plant-cell cultures of the Chinese herbal plant, Tripterygium wilfordii. Isolation and characterization of diterpenes. Recl. Trav. Chim. Pays-Bas 1996, 93, 77–93. [Google Scholar] [CrossRef]
- Takaishi, Y.; Warhshi, N.; Tateishi, H.; Kawazoe, K.; Miyagi, K.; Li, K.; Duan, H. Phenolic diterpenes from Tripterygium wilfordii var. regelii. Phytochemistry 1997, 45, 979–984. [Google Scholar] [CrossRef]
- van Tamelen, E.E.; Leiden, T.M. Total Synthesis of I-Triptonide and I-Triptolide. J. Am. Chem. Soc. 1982, 104, 1785–1786. [Google Scholar] [CrossRef]
- Zhou, B.; Li, X.; Tang, H.; Miao, Z.; Feng, H.; Li, Y. Total synthesis of novel D-ring-modified triptolide analogs: Structure-cytotoxic activity relationship studies on the D-ring of triptolide. Org. Biomol. Chem. 2011, 9, 3176–3179. [Google Scholar] [CrossRef] [PubMed]
- Hamberger, B.; Ohnishi, T.; Hamberger, B.; Séguin, A.; Bohlmann, J. Evolution of Diterpene Metabolism: Sitka Spruce CYP720B4 Catalyzes Multiple Oxidations in Resin Acid Biosynthesis of Conifer Defense against Insects. Plant Physiol. 2011, 157, 1677–1695. [Google Scholar] [CrossRef] [PubMed]
- Andersen-Ranberg, J.; Kongstad, K.T.; Nielsen, M.T.; Jensen, N.B.; Pateraki, I.; Bach, S.S.; Hamberger, B.; Zerbe, P.; Staerk, D.; Bohlmann, J.; et al. Expanding the landscape of diterpene structural diversity through stereochemically controlled combinatorial biosynthesis. Angew. Chem. Int. Ed. 2016, 55, 2142–2146. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Liu, Y.; Huang, L.; Zhang, X. Production of miltiradiene by metabolically engineered Saccharomyces cerevisiae. Biotechnol. Bioeng. 2012, 109, 2845–2853. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.J.; Gao, W.; Rong, Q.; Jin, G.; Chu, H.; Liu, W.; Yang, W.; Zhu, Z.; Li, G.; Zhu, G.; et al. Modular pathway engineering of diterpenoid synthases and the mevalonic acid pathway for miltiradiene production. J. Am. Chem. Soc. 2012, 134, 3234–3241. [Google Scholar] [CrossRef] [PubMed]
- Ignea, C.; Ioannou, E.; Georgantea, P.; Loupassaki, S.; Trikka, F.A.; Kanellis, A.K.; Makris, A.M.; Roussis, V.; Kampranis, S.C. Reconstructing the chemical diversity of labdane-type diterpene biosynthesis in yeast. Metab. Eng. 2015, 28, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Gonz, F.J.; Almendros, G.; Martin, F. An evaluation of the differences in the composition of humic acids in soils under oak and pine forest by GC-MS after mild degradation. Plant Soil 1987, 103, 83–88. [Google Scholar]
- Lanza, A.M.; Curran, K.A.; Rey, L.G.; Alper, H.S. A condition-specific codon optimization approach for improved heterologous gene expression in Saccharomyces cerevisiae. BMC Syst. Biol. 2014, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.J.; Flory, J.E.; Jetter, R.; Ravn, M.M.; Lee, H.J.; Coates, R.M.; Croteau, R.B. Abietadiene synthase from grand fir (Abies grandis): Characterization and mechanism of action of the “pseudomature” recombinant enzyme. Biochemistry 2000, 39, 15592–15602. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.C.; Wildung, M.R.; Jin, A.Q.; Dalal, D.; Oliver, J.S.; Coates, R.M.; Croteau, R. Heterologous expression and characterization of a “Pseudomature” form of taxadiene synthase involved in paclitaxel (Taxol) biosynthesis and evaluation of a potential intermediate and inhibitors of the multistep diterpene cyclization reaction. Arch. Biochem. Biophys. 2000, 379, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Brückner, K.; Božić, D.; Manzano, D.; Papaefthimiou, D.; Pateraki, I.; Scheler, U.; Ferrer, A.; de Vos, R.C.H.; Kanellis, A.K.; Tissier, A. Characterization of two genes for the biosynthesis of abietane-type diterpenes in rosemary (Rosmarinus officinalis) glandular trichomes. Phytochemistry 2014, 101, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, B.; Astudillo, L.; Rodríguez, J.A.; Yáñez, T.; Theoduloz, C.; Schmeda-Hirschmann, G. Gastroprotective and cytotoxic effect of dehydroabietic acid derivatives. Pharmacol. Res. 2005, 52, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Hirai, S.; Goto, T.; Kuroyanagi, K.; Lee, J.Y.; Uemura, T.; Ezaki, Y.; Takahashi, N.; Kawada, T. Dehydroabietic acid, a phytochemical, acts as ligand for PPARs in macrophages and adipocytes to regulate inflammation. Biochem. Biophys. Res. Commun. 2008, 369, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.C.; Wang, M.; Pan, Y.M.; Yao, G.Y.; Wang, H.S.; Tian, X.Y.; Qin, J.K.; Zhang, Y. Synthesis and antitumor activities of novel thiourea α- aminophosphonates from dehydroabietic acid. Eur. J. Med. Chem. 2013, 69, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Nour-Eldin, H.H.; Hansen, B.G.; Nørholm, M.H.H.; Jensen, J.K.; Halkier, B.A. Advancing uracil-excision based cloning towards an ideal technique for cloning PCR fragments. Nucleic Acids Res. 2006, 34, e122. [Google Scholar] [CrossRef] [PubMed]
- Bach, S.S.; Bassard, J.-É.; Ranberg, J.A.; Møldrup, M.E.; Simonsen, H.T.; Hamberger, B. Plant Isoprenoids. In Plant Isoprenoids, Methods in Molecular Biology; Rodríguez, M., Ed.; Springer: New York, NY, USA, 2014; Volume 1153, pp. 285–299. [Google Scholar]
- Partow, S.; Siewers, V.; Bjørn, S.; Nielsen, J.; Maury, J. Characterization of different promoters for designing a new expression vector in Saccharomyces cerevisiae. Yeast 2010, 27, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.D.; Woods, R.A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Methods Enzymol. 2002, 350, 87–96. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Genotype |
|---|---|
| EYS2010 | MATα, MAL2-8C, SUC2, his3Δ1, leu2-3_112, ura3-52, YCT1::PGPD1-tHMGR1-TCYC1 |
| YS1 | [CEN/ARS/pPGK1-CO_TwTPS9-tCYC1/URA3], [CEN/ARS/pTEF1-CO_TwTPS27-tADH1/HIS3], [CEN/ARS/pCYC1-GGPPS7-tADH2/LEU2] |
| YS2 | [CEN/ARS/pPGK1-CO_Δ46TwTPS9-tCYC1/URA3], [CEN/ARS/pTEF1-COΔ41TwTPS27-tADH1/HIS3], [CEN/ARS/pCYC1-GGPPS7-tADH2/LEU2] |
| YS3 | [CEN/ARS/pPGK1-WT_TwTPS9-tCYC1/URA3],[CEN/ARS/pTEF1-WT_TwTPS27-tADH1/HIS3], [CEN/ARS/pCYC1-GGPPS7-tADH2/LEU2] |
| YS4 | [CEN/ARS/pPGK1-WT_Δ46TwTPS9-tCYC1/URA3], [CEN/ARS/pTEF1-WTΔ41TwTPS27-tADH1/HIS3], [CEN/ARS/pCYC1-GGPPS7-tADH2/LEU2] |
| YS2-1 | [CEN/ARS/pPGK1-CO_Δ46TwTPS9-tCYC1/ pTEF1-COΔ41TwTPS27-tADH11//URA3], [CEN/ARS/pPGK1-CO_PsCYP720B4-tCYC1/pTEF1-ATR1-tADH11/HIS3], [CEN/ARS/pCYC1-GGPPS7-tCYC1/LEU2] |
| YS2-2 | [CEN/ARS/pPGK1-CO_Δ46TwTPS9-tCYC1/ pTEF1-COΔ41TwTPS27-tADH11//URA3], [CEN/ARS/pPGK1-CO_PsCYP720B4-tCYC1/pTEF1-ATR1-tADH11/HIS3], [CEN/ARS/pCYC1-GGPPS7-tCYC1/LEU2] |
| ATR1 (−) | [CEN/ARS/pPGK1-CO_Δ46TwTPS9-tCYC1/pTEF1-COΔ41TwTPS27-tADH11//URA3], [CEN/ARS/pTEF1-ATR1-tADH11/HIS3], [CEN/ARS/pCYC1-GGPPS7-tCYC1/LEU2] |
| SrCPR (−) | [CEN/ARS/pPGK1-CO_Δ46TwTPS9-tCYC1/ pTEF1-COΔ41TwTPS27-tADH11//URA3], [CEN/ARS/pTEF1-SrCPR-tADH11/HIS3], [CEN/ARS/pCYC1-GGPPS7-tCYC1/LEU2] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forman, V.; Callari, R.; Folly, C.; Heider, H.; Hamberger, B. Production of Putative Diterpene Carboxylic Acid Intermediates of Triptolide in Yeast. Molecules 2017, 22, 981. https://doi.org/10.3390/molecules22060981
Forman V, Callari R, Folly C, Heider H, Hamberger B. Production of Putative Diterpene Carboxylic Acid Intermediates of Triptolide in Yeast. Molecules. 2017; 22(6):981. https://doi.org/10.3390/molecules22060981
Chicago/Turabian StyleForman, Victor, Roberta Callari, Christophe Folly, Harald Heider, and Björn Hamberger. 2017. "Production of Putative Diterpene Carboxylic Acid Intermediates of Triptolide in Yeast" Molecules 22, no. 6: 981. https://doi.org/10.3390/molecules22060981
APA StyleForman, V., Callari, R., Folly, C., Heider, H., & Hamberger, B. (2017). Production of Putative Diterpene Carboxylic Acid Intermediates of Triptolide in Yeast. Molecules, 22(6), 981. https://doi.org/10.3390/molecules22060981