Cytotoxic Polyketides with an Oxygen-Bridged Cyclooctadiene Core Skeleton from the Mangrove Endophytic Fungus Phomosis sp. A818


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
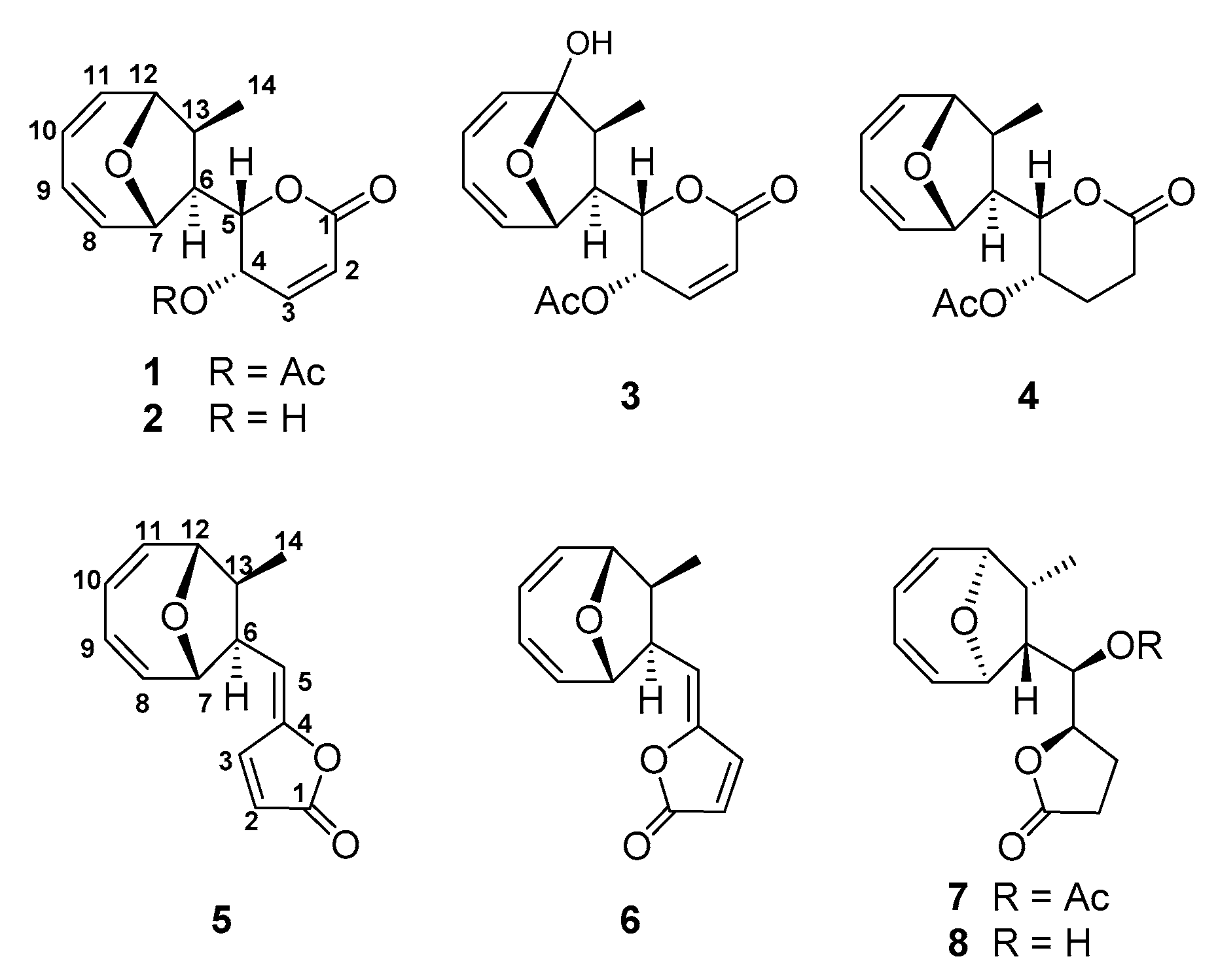
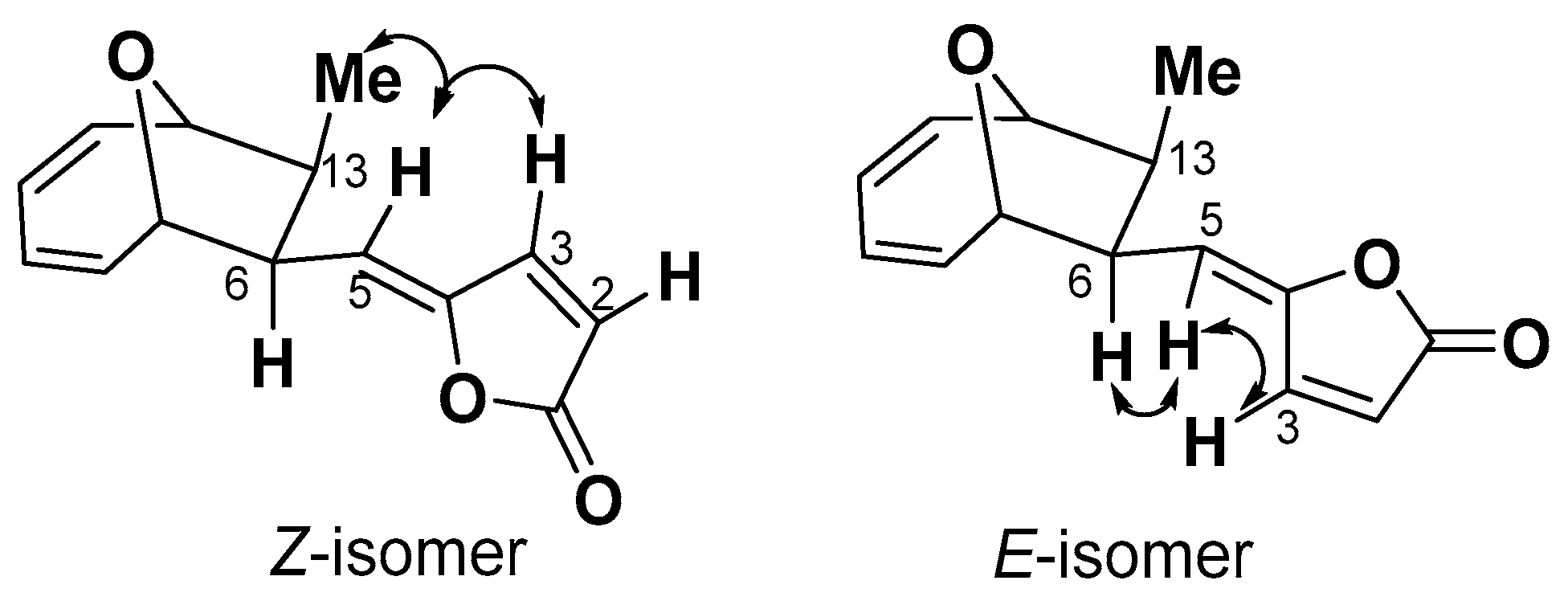
2.1. Structure Elucidation
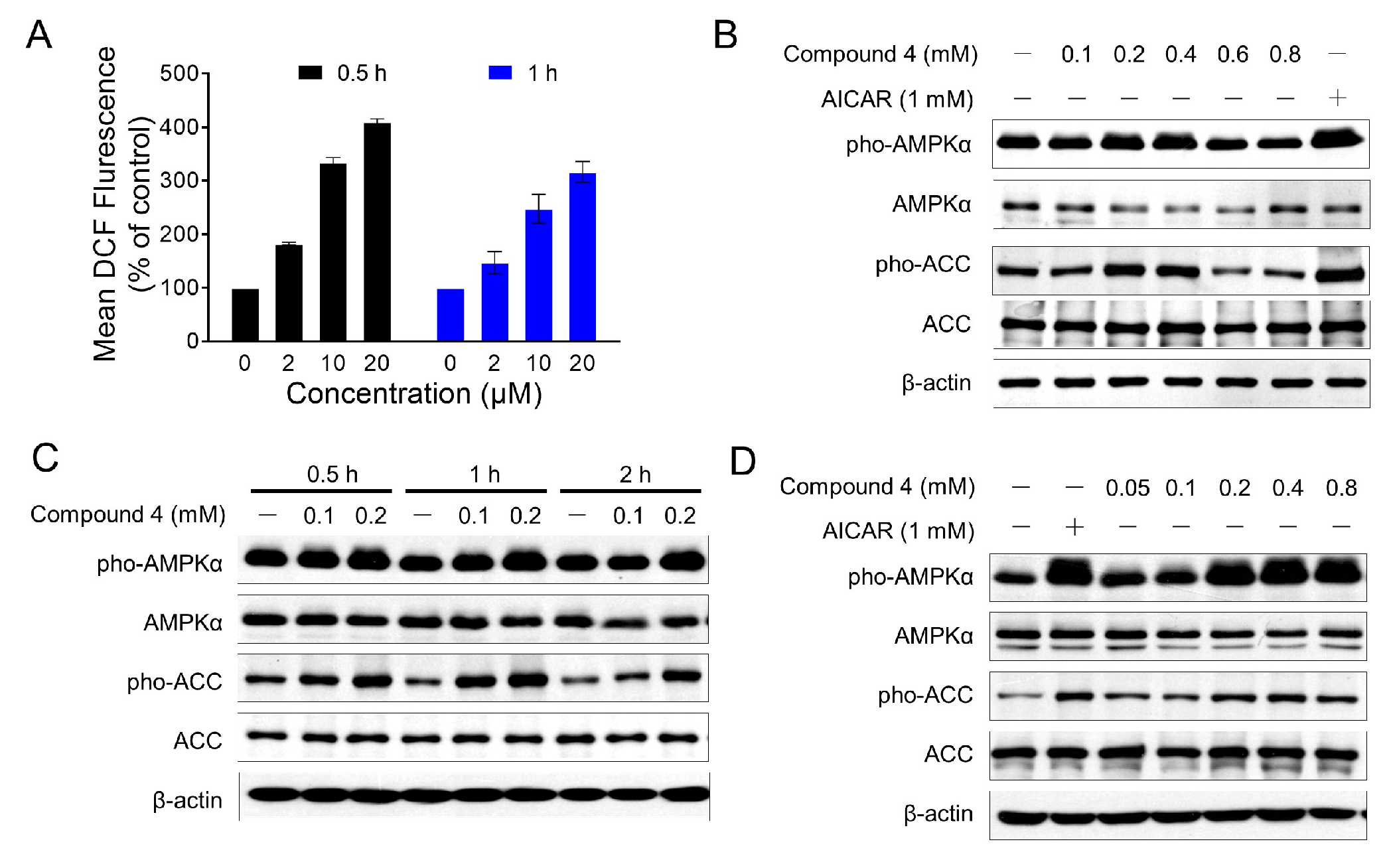
2.2. Cytotoxicity Study
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Microbial Strains
4.3. Culture Conditions and Extraction
4.4. Structure Isolation and Purification
4.5. Cell Culture and Cytotoxicity Assays
4.6. Western Blot Analyses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, H.; Song, Y.; Tan, R. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Ali, M.S.; Hussain, S.; Jabbar, A.; Ashraf, M.; Lee, Y.S. Marine natural products of fungal origin. Nat. Prod. Rep. 2007, 24, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Huang, Y.; Fang, M.; Wang, J.; Zheng, Z.; Su, W. Cytotoxic and antimicrobial metabolites from marine lignicolous fungi, Diaporthe sp. FEMS Microbiol. Lett. 2005, 251, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Lin, Y.C.; She, Z.G.; Du, D.S.; Chan, W.L.; Zheng, Z.H. Paeciloxanthone, a new cytotoxic xanthone from the marine mangrove fungus Paecilomyces sp. (Tree1–7). J. Asian Nat. Prod. Res. 2008, 10, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Wang, J.; Huang, Y.; Zheng, Z.; Song, S.; Zhang, Y.; Su, W. Metabolites from mangrove endophytic fungus Dothiorella sp. Acta Oceanol. Sin. 2004, 23, 541–547. [Google Scholar]
- Zhan, Y.; Du, X.; Chen, H.; Liu, J.; Zhao, B.; Huang, D.; Li, G.; Xu, Q.; Zhang, M.; Weimer, B.C.; et al. Cytosporone B is an agonist for nuclear orphan receptor Nur77. Nat. Chem. Biol. 2008, 4, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yi, Y.; Chen, J.; Zhang, W.; Huang, Y.; Zheng, Z.; Song, S.; Shen, Y. Probing Target and Designing Nicotinoid Derivatives for Antitumor Leading Compound MED. Chem. J. Chin. Univ.-Chin. 2010, 31, 1184–1189. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, L.; Yang, L.; Huang, Y.; Li, S.; Shen, Y. Phomopsidone A, a novel depsidone metabolite from the mangrove endophytic fungus Phomopsis sp. A123. Fitoterapia 2014, 96, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; McPhail, A.T.; Krainer, E.; Katz, B.; Pearce, C.; Boros, C.; Caceres, B.; Smith, D.; Houck, D.R. Mycoepoxydiene represents a novel class of fungal metabolites. Tetrahedron Lett. 1999, 40, 1479–1482. [Google Scholar] [CrossRef]
- Takao, K.; Watanabe, G.; Yasui, H.; Tadano, K. Total synthesis of (+/−)-mycoepoxydiene, a novel fungal metabolite having an oxygen-bridged cyclooctadiene skeleton. Org. Lett. 2002, 4, 2941–2943. [Google Scholar] [CrossRef] [PubMed]
- Takao, K.; Yasui, H.; Yamamoto, S.; Sasaki, D.; Kawasaki, S.; Watanabe, G.; Tadano, K. Asymmetric total syntheses of (+)-mycoepoxydiene and related natural product (−)-1893A: Application of one-pot ring-opening/cross/ring-closing metathesis to construct their 9-oxabicyclo[4.2.1]nona-2,4-diene skeleton. J. Org. Chem. 2004, 69, 8789–8795. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, B.; Yi, Y.; Zhang, W.; Wu, X.; Zhang, L.; Shen, Y. Mycoepoxydiene, a fungal polyketide inhibits MCF-7 cells through simultaneously targeting p53 and NF-κB pathways. Biochem. Pharmacol. 2012, 84, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, B.; Zhang, W.; Wu, X.; Wang, R.; Huang, Y.; Chen, D.; Park, K.; Weimer, B.C.; Shen, Y. Mycoepoxydiene, a fungal polyketide, induces cell cycle arrest at the G2/M phase and apoptosis in HeLa cells. Bioorg. Med. Chem. Lett. 2010, 20, 7054–7058. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Chen, Q.; Liu, K.; Mo, P.L.; Zhu, J.W.; Zhuang, M.Q.; Shen, Y.M.; Yu, C.D. Mycoepoxydiene inhibits antigen-stimulated activation of mast cells and suppresses IgE-mediated anaphylaxis in mice. Int. Immunopharmacol. 2013, 17, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, Q.; Xia, X.; Mo, P.; Shen, Y.; Yu, C. Mycoepoxydiene suppresses RANKL-induced osteoclast differentiation and reduces ovariectomy-induced bone loss in mice. Appl. Microbiol. Biotechnol. 2013, 97, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, M.; Huang, Y.; Chea, S.K.; Zheng, Z.; Qian, X.; Shen, Y. New and highly efficient methodology for screening high-yield strains of cytotoxic deacetylmycoepoxydiene (DAM). Lett. Appl. Microbiol. 2011, 52, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, W.; Xu, W.; Shen, Y.; Du, L. Optimization of genome shuffling for high-yield production of the antitumor deacetylmycoepoxydiene in an endophytic fungus of mangrove plants. Appl. Microbiol. Biotechnol. 2016, 100, 7491–7498. [Google Scholar] [CrossRef] [PubMed]
- Prachya, S.; Wiyakrutta, S.; Sriubolmas, N.; Ngamrojanavanich, N.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Cytotoxic mycoepoxydiene derivatives from an endophytic fungus Phomopsis sp. isolated from Hydnocarpus anthelminthicus. Planta Med. 2007, 73, 1418–1420. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yang, L.; Huang, Y.; Zheng, Z.; Song, S.; Su, W. Method for Preparing Deacetyl mycoepoxydiene from Metabolite of Phomosis A-1–2-3 and Its Application in Antitumor Drugs and Other Bioactive Precursor. Chinese Patent CN1903858 A 20070131, 31 January 2007. [Google Scholar]
- Kornsakulkarn, J.; Somyong, W.; Supothina, S.; Boonyuen, N.; Thongpanchang, C. Bioactive oxygen-bridged cyclooctadienes from endophytic fungus Phomopsis sp. BCC 45011. Tetrahedron 2015, 71, 9112–9116. [Google Scholar] [CrossRef]
- Chen, G.; Lin, Y.; Wen, L.; Vrijmoedc, L.L.P.; Gareth, J.E.B. Two new metabolites of a marine endophytic fungus (No. 1893) from an estuarine mangrove on the South China Sea coast. Tetrahedron 2003, 59, 4907–4909. [Google Scholar] [CrossRef]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]
- Huang, Y.; Wang, J.; Li, G.; Zheng, Z.; Su, W. Antitumor and antifungal activities in endophytic fungi isolated from pharmaceutical plants Taxus mairei, Cephalataxus fortunei and Torreya grandis. FEMS Immunol. Med. Microbiol. 2001, 31, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.; Lin, M.; Liu, S.; Chiang, W.; Hsieh, W.; Cheng, Y.; Hsu, K.; Liu, Y. Arecoline-mediated inhibition of AMP-activated protein kinase through reactive oxygen species is required for apoptosis induction. Oral Oncol. 2011, 47, 345–351. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds 1 and 2 are available from the authors. |



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Zhao, B.; Du, L.; Shen, Y. Cytotoxic Polyketides with an Oxygen-Bridged Cyclooctadiene Core Skeleton from the Mangrove Endophytic Fungus Phomosis sp. A818. Molecules 2017, 22, 1547. https://doi.org/10.3390/molecules22091547
Zhang W, Zhao B, Du L, Shen Y. Cytotoxic Polyketides with an Oxygen-Bridged Cyclooctadiene Core Skeleton from the Mangrove Endophytic Fungus Phomosis sp. A818. Molecules. 2017; 22(9):1547. https://doi.org/10.3390/molecules22091547
Chicago/Turabian StyleZhang, Wei, Baobing Zhao, Liangcheng Du, and Yuemao Shen. 2017. "Cytotoxic Polyketides with an Oxygen-Bridged Cyclooctadiene Core Skeleton from the Mangrove Endophytic Fungus Phomosis sp. A818" Molecules 22, no. 9: 1547. https://doi.org/10.3390/molecules22091547
APA StyleZhang, W., Zhao, B., Du, L., & Shen, Y. (2017). Cytotoxic Polyketides with an Oxygen-Bridged Cyclooctadiene Core Skeleton from the Mangrove Endophytic Fungus Phomosis sp. A818. Molecules, 22(9), 1547. https://doi.org/10.3390/molecules22091547