The Impact of Lipid Types and Liposomal Formulations on Osteoblast Adiposity and Mineralization



Abstract
:1. Introduction
2. Results
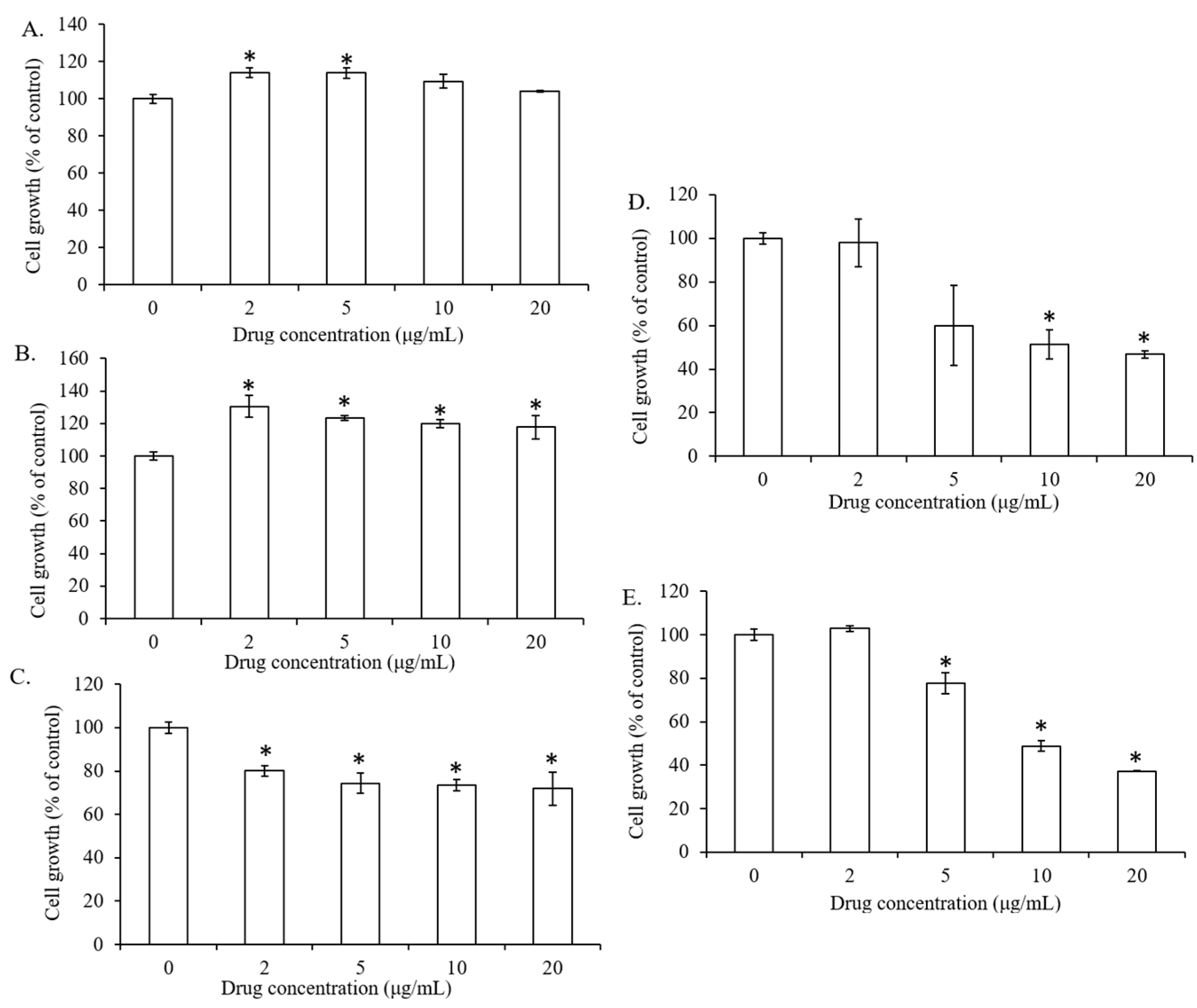
2.1. The Effect of Lipid Types on Cell Viability of 7F2 Osteoblasts
2.2. The Influence of Lipid Types on the Lipid Droplet Formation of 7F2 Osteoblasts
2.3. The Impact of Lipid Types on Osteogenesis in 7F2 Osteoblasts
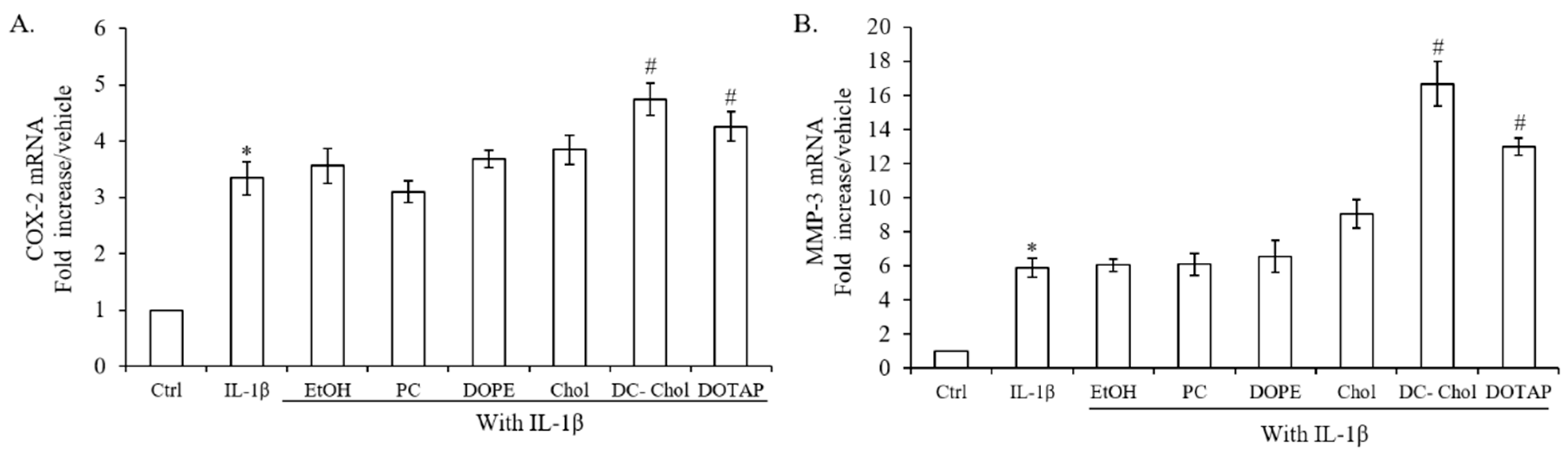
2.4. The Impact of Lipid Types on the Inflammatory Response of IL-1β-stimulated 7F2 Osteoblasts
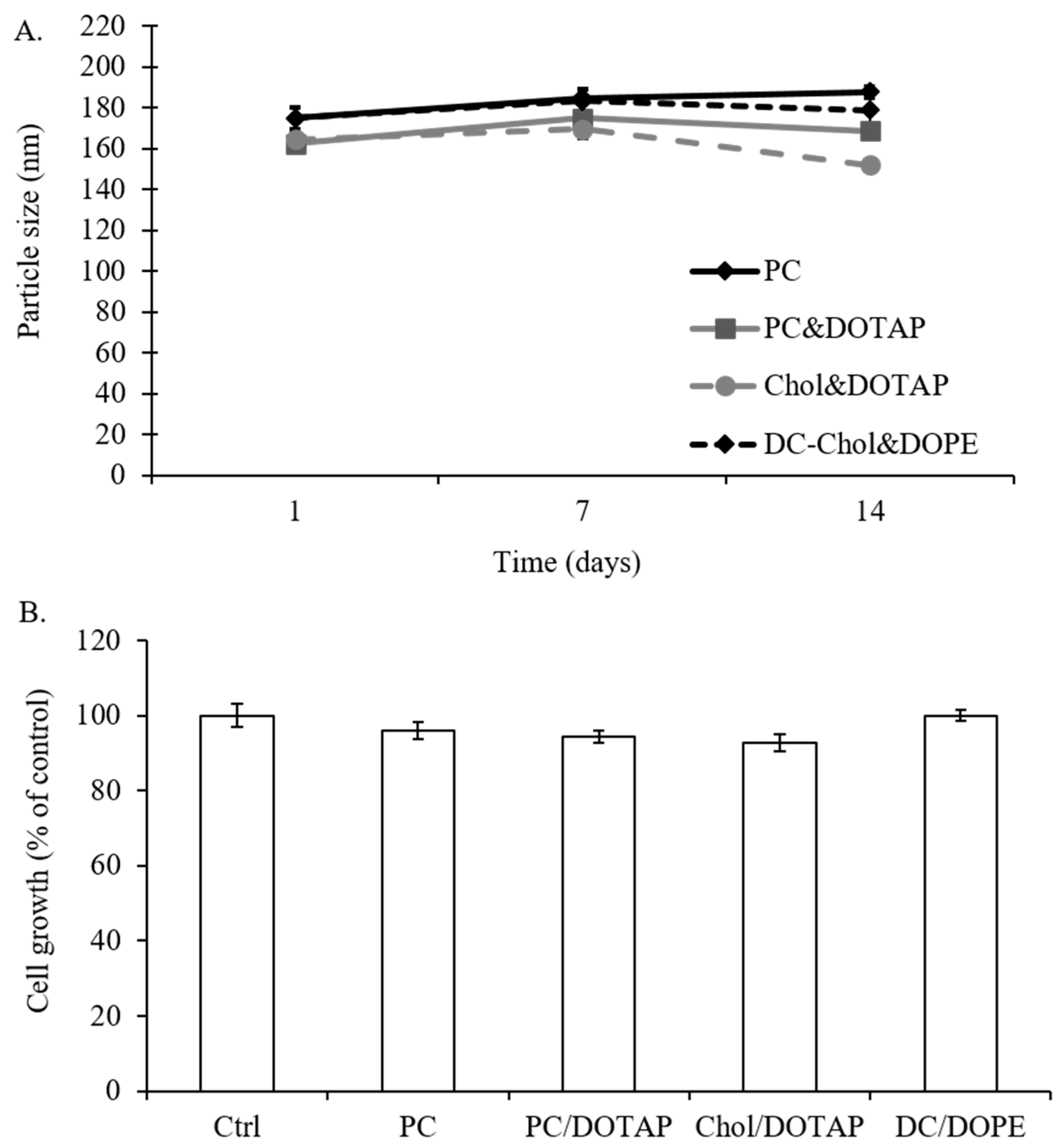
2.5. The Effect of Liposomal Formulations on Particle Size and Stability and Cell Viability of 7F2 Osteoblasts
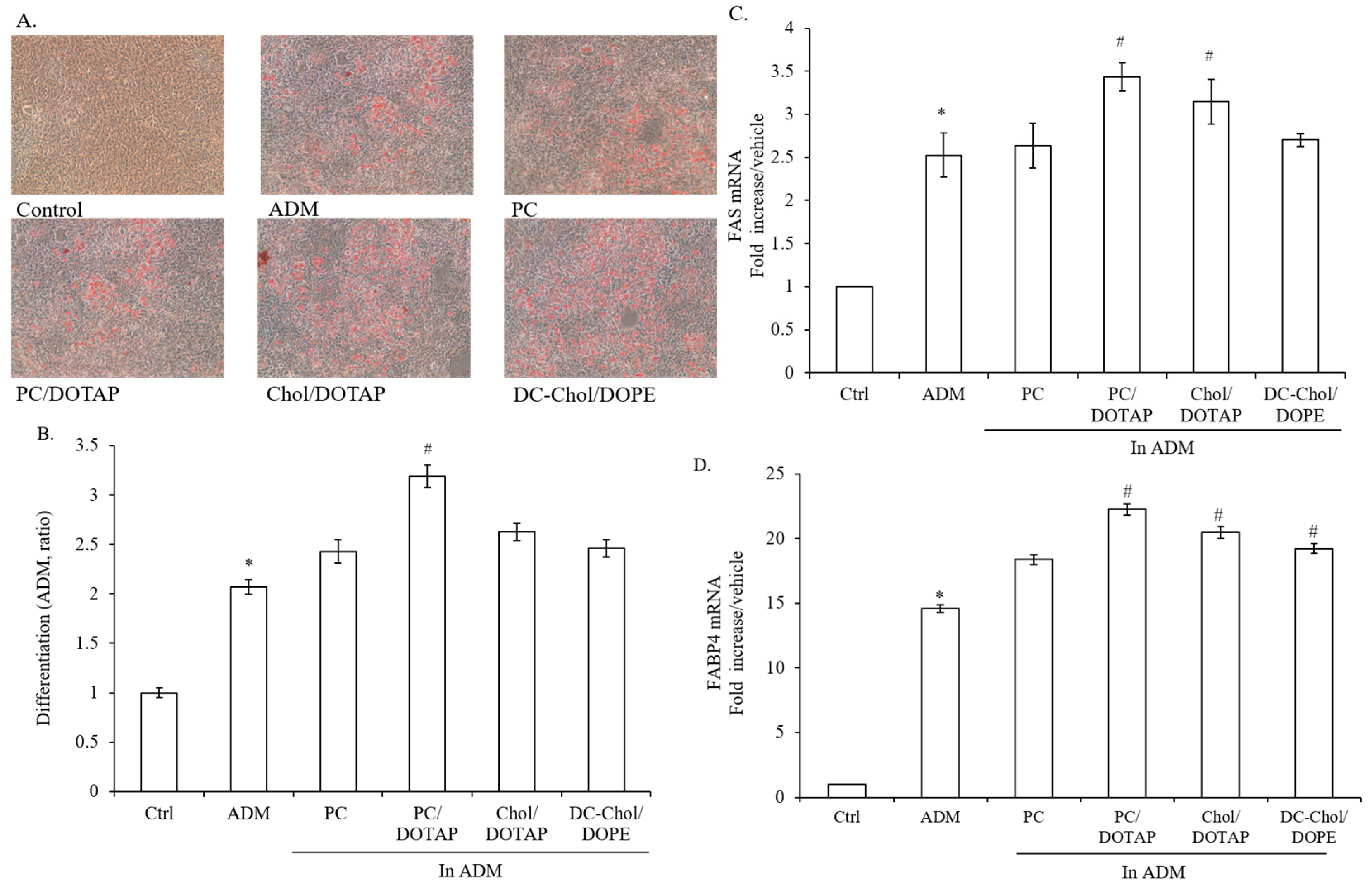
2.6. The Impact of Liposomal Formulations on the Lipid Droplet Formation in 7F2 Osteoblasts
2.7. The Impact of Liposomal Formulations on Osteogenesis in 7F2 Osteoblasts
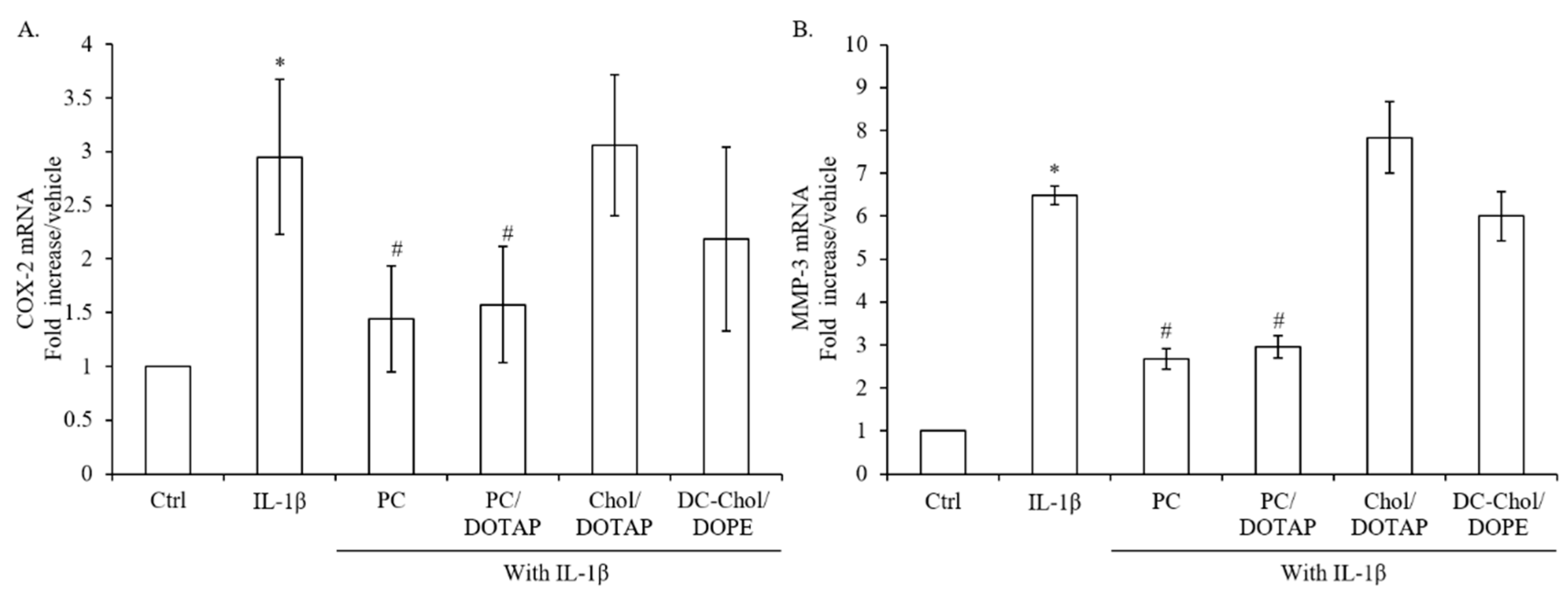
2.8. The Inflammatory Responses of IL-1β-stimulated Osteoblasts after Treatment with Various Liposomal Formulations
3. Discussion
4. Materials and Methods
4.1. Materials and Cell Culture
4.2. MTT Cell Growth Assay
4.3. Liposomal Formulation
4.4. Particle Characterization
4.5. Oil Red O Staining
4.6. Alkaline Phosphatase (ALP) Activity Analysis
4.7. Alizarin red S Staining for Osteoblast Mineralization
4.8. Quantitative Real-Time PCR
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- James, A.W. Review of signaling pathways governing MSC osteogenic and adipogenic differentiation. Scientifica (Cairo) 2013, 2013, 684736. [Google Scholar] [CrossRef] [PubMed]
- Dragojevič, J.; Logar, D.B.; Komadina, R.; Marc, J. Osteoblastogenesis and adipogenesis are higher in osteoarthritic than in osteoporotic bone tissue. Arch. Med. Res. 2011, 42, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Van de Vyver, M.; Andrag, E.; Cockburn, I.L.; Ferris, W.F. Thiazolidinedione-induced lipid droplet formation during osteogenic differentiation. J. Endocrinol. 2014, 223, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Hoshiba, T.; Kawazoe, N.; Chen, G. The balance of osteogenic and adipogenic differentiation in human mesenchymal stem cells by matrices that mimic stepwise tissue development. Biomaterials 2012, 33, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.H.; Reimer, R.A.; Seerattan, R.A.; Leonard, T.R.; Herzog, W. Using diet-induced obesity to understand a metabolic subtype of osteoarthritis in rats. Osteoarthr. Cartil. 2015, 23, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.C.; Su, Y.H.; Lin, Y.J.; Chen, P.J.; Shi, C.S.; Chen, C.N.; Chang, H.I. Evaluation of the protective effects of curcuminoid (curcumin and bisdemethoxycurcumin)-loaded liposomes against bone turnover in a cell-based model of osteoarthritis. Drug Des. Dev. Ther. 2015, 9, 2285–2300. [Google Scholar] [CrossRef]
- Kc, R.; Voigt, R.; Li, X.; Forsyth, C.B.; Ellman, M.B.; Summa, K.C.; Turek, F.W.; Keshavarzian, A.; Kim, J.S.; Im, H.J. Induction of osteoarthritis-like pathologic changes by chronic alcohol consumption in an experimental mouse model. Arthritis Rheumatol. 2015, 67, 1678–1680. [Google Scholar] [CrossRef] [PubMed]
- Belcaro, G.; Cesarone, M.R.; Dugall, M.; Pellegrini, L.; Ledda, A.; Grossi, M.G.; Togni, S.; Appendino, G. Efficacy and safety of Meriva®, a curcumin- hosphatidylcholine complex, during extended administration in osteoarthritis patients. Altern. Med. Rev. 2010, 15, 337–344. [Google Scholar] [PubMed]
- Lv, H.; Zhang, S.; Wang, B.; Cui, S.; Yan, J. Toxicity of cationic lipids and cationic polymers in gene delivery. J. Control. Release 2006, 114, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Tseng, L.P.; Liang, H.J.; Chung, T.W.; Huang, Y.Y.; Liu, D.Z. Liposomes incorporated with cholesterol for drug release triggered by magnetic field. J. Med. Biol. Eng. 2007, 27, 29–34. [Google Scholar]
- Goyal, K.; Huang, L. Gene therapy using Dc-Chol liposomes. J. Liposome Res. 1995, 5, 49–60. [Google Scholar] [CrossRef]
- Galović Rengel, R.; Barisić, K.; Pavelić, Z.; Zanić Grubisić, T.; Cepelak, I.; Filipović-Grcić, J. High efficiency entrapment of superoxide dismutase into mucoadhesive chitosan-coated liposomes. Eur. J. Pharm. Sci. 2002, 15, 441–448. [Google Scholar] [CrossRef]
- Chang, H.I.; Yeh, M.K. Clinical development of liposome-based drugs: Formulation, characterization, and therapeutic efficacy. Int. J. Nanomed. 2012, 7, 49–60. [Google Scholar] [CrossRef]
- Komatsu, H.; Okada, S. Ethanol-induced aggregation and fusion of small phosphatidylcholine liposome: Participation of interdigitated membrane formation in their processes. Biochim. Biophys. Acta 1995, 1235, 270–280. [Google Scholar] [CrossRef]
- Shimanouchi, T.; Ishii, H.; Yoshimoto, N.; Umakoshi, H.; Kuboi, R. Calcein permeation across phosphatidylcholine bilayer membrane: Effects of membrane fluidity, liposome size, and immobilization. Colloids Surf. B Biointerfaces 2009, 73, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Baczynska, D.; Widerak, K.; Ugorski, M.; Langner, M. Surface charge and the association of liposomes with colon carcinoma cells. Zeitschrift für Naturforschung C 2001, 56, 872–877. [Google Scholar] [CrossRef]
- Ciani, L.; Ristori, S.; Salvati, A.; Calamai, L.; Martini, G. DOTAP/DOPE and DC-Chol/DOPE lipoplexes for gene delivery: Zeta potential measurements and electron spin resonance spectra. Biochim. Biophys. Acta 2004, 1664, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Crook, K.; Stevenson, B.J.; Dubouchet, M.; Porteous, D.J. Inclusion of cholesterol in DOTAP transfection complexes increases the delivery of DNA to cells in vitro in the presence of serum. Gene Ther. 1998, 5, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Marchini, C.; Montani, M.; Amici, A.; Amenitsch, H.; Marianecci, C.; Pozzi, D.; Caracciolo, G. Structural stability and increase in size rationalize the efficiency of lipoplexes in serum. Langmuir 2009, 25, 3013–3021. [Google Scholar] [CrossRef] [PubMed]
- Akopian, D.; Kawashima, R.L.; Medh, J.D. Phosphatidylcholine-Mediated Aqueous Diffusion of Cellular Cholesterol Down-Regulates the ABCA1 Transporter in Human Skin Fibroblasts. Int. J. Biochem. Res. Rev. 2015, 5, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kwon, M.S.; Son, J.; Kang, S.W.; Song, Y. Selective effect of phosphatidylcholine on the lysis of adipocytes. PLoS ONE 2017, 12, e0176722. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y.; Sugiyama, A.; Yamamoto, T.; Naito, M.; Noguchi, N.; Yokoyama, S.; Tsujita, M.; Kawabe, Y.; Kobayashi, M.; Izumi, A.; et al. Lipid accumulation in smooth muscle cells under LDL loading is independent of LDL receptor pathway and enhanced by hypoxic conditions. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1712–1719. [Google Scholar] [CrossRef] [PubMed]
- Wobser, H.; Dorn, C.; Weiss, T.S.; Amann, T.; Bollheimer, C.; Büttner, R.; Schölmerich, J.; Hellerbrand, C. Lipid accumulation in hepatocytes induces fibrogenic activation of hepatic stellate cells. Cell Res. 2009, 19, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Moseti, D.; Regassa, A.; Kim, W.-K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [PubMed]
- Shan, T.; Liu, W.; Kuang, S. Fatty acid binding protein 4 expression marks a population of adipocyte progenitors in white and brown adipose tissues. FASEB J. 2013, 27, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Grimaud, E.; Soubigou, L.; Couillaud, S.; Coipeau, P.; Moreau, A.; Passuti, N.; Gouin, F.; Redini, F.; Heymann, D. Receptor activator of nuclear factor kappaB ligand (RANKL)/osteoprotegerin (OPG) ratio is increased in severe osteolysis. Am. J. Pathol. 2003, 163, 2021–2031. [Google Scholar] [CrossRef]
- Kwan Tat, S.; Pelletier, J.P.; Lajeunesse, D.; Fahmi, H.; Lavigne, M.; Martel-Pelletier, J. The differential expression of osteoprotegerin (OPG) and receptor activator of nuclear factor kappaB ligand (RANKL) in human osteoarthritic subchondral bone osteoblasts is an indicator of the metabolic state of these disease cells. Clin. Exp. Rheumatol. 2008, 26, 295–304. [Google Scholar] [PubMed]
- Towle, C.A.; Hung, H.H.; Bonassar, L.J.; Treadwell, B.V.; Mangham, D.C. Detection of interleukin-1 in the cartilage of patients with osteoarthritis: A possible autocrine/paracrine role in pathogenesis. Osteoarthr. Cartil. 1997, 5, 293–300. [Google Scholar] [CrossRef]
- Fernandes, J.C.; Martel-Pelletier, J.; Pelletier, J.P. The role of cytokines in osteoarthritis pathophysiology. Biorheology 2002, 39, 237–246. [Google Scholar] [PubMed]
- Gao, X.; Huang, L. Cationic liposomes and polymres for gene transfer. J. Liposome Res. 1993, 3, 17–30. [Google Scholar] [CrossRef]
- Kearns, M.D.; Patel, Y.N.; Savva, M. Physicochemical characteristics associated with transfection of cationic cholesterol-based gene delivery vectors in the presence of DOPE. Chem. Phys. Lipids 2010, 163, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Pires, P.; Simões, S.; Nir, S.; Gaspar, R.; Düzgünes, N.; Pedroso de Lima, M.C. Interaction of cationic liposomes and their DNA complexes with monocytic leukemia cells. Biochim. Biophys. Acta 1999, 1418, 71–84. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Ferraz, M.P.; Monteiro, F.J.; Simões, S. Cationic liposome-DNA complexes as gene delivery vectors: Development and behaviour towards bone-like cells. Acta Biomater. 2009, 5, 2142–2151. [Google Scholar] [CrossRef] [PubMed]
- Van der Geest, T.; Metselaar, J.M.; Gerrits, D.; van Lent, P.L.; Storm, G.; Laverman, P.; Boerman, O.C. [(18)]F FDG PET/CT imaging to monitor the therapeutic effect of liposome-encapsulated prednisolone in experimental rheumatoid arthritis. J. Control. Release 2015, 209, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.R.; Lazarenko, O.P.; Shankar, K.; Blackburn, M.L.; Badger, T.M.; Ronis, M.J. A role for ethanol-induced oxidative stress in controlling lineage commitment of mesenchymal stromal cells through inhibition of Wnt/beta-catenin signaling. J. Bone Miner. Res. 2010, 25, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Rezq, A.A.; Labib, F.A.; Attia, A.E.M. Effect of some dietary oils and fats on serum lipid profile, calcium absorption and bone mineralization in mice. Pak. J. Nutr. 2010, 9, 643–650. [Google Scholar] [CrossRef]
- Le Lay, S.; Hajduch, E.; Lindsay, M.R.; Le Lièpvre, X.; Thiele, C.; Ferré, P.; Parton, R.G.; Kurzchalia, T.; Simons, K.; Dugail, I. Cholesterol-induced caveolin targeting to lipid droplets in adipocytes: A role for caveolar endocytosis. Traffic 2006, 7, 549–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manickam, E.; Sinclair, A.J.; Cameron-Smith, D. Suppressive actions of eicosapentaenoic acid on lipid droplet formation in 3T3-L1 adipocytes. Lipids Health Dis. 2010, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Czamara, K.; Majzner, K.; Selmi, A.; Baranska, M.; Ozaki, Y.; Kaczor, A. Unsaturated lipid bodies as a hallmark of inflammation studied by Raman 2D and 3D microscopy. Sci. Rep. 2017, 7, 40889. [Google Scholar] [CrossRef] [PubMed]
- Wernstedt Asterholm, I.; Tao, C.; Morley, T.S.; Wang, Q.A.; Delgado-Lopez, F.; Wang, Z.V.; Scherer, P.E. Adipocyte inflammation is essential for healthy adipose tissue expansion and remodeling. Cell Metab. 2014, 20, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Treede, I.; Braun, A.; Sparla, R.; Kühnel, M.; Giese, T.; Turner, J.R.; Anes, E.; Kulaksiz, H.; Füllekrug, J.; Stremmel, W.; et al. Anti-inflammatory effects of phosphatidylcholine. J. Biol. Chem. 2007, 282, 27155–27164. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Braun, A.; Füllekrug, J.; Stremmel, W.; Ehehalt, R. Lipid based therapy for ulcerative colitis-modulation of intestinal mucus membrane phospholipids as a tool to influence inflammation. Int. J. Mol. Sci. 2010, 11, 4149–4164. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.; El-Meligy, A.; Fetaih, H.; Dessouki, A.; Stoica, G.; Barhoumi, R. Effect of curcumin and Meriva on the lung metastasis of murine mammary gland adenocarcinoma. In Vivo 2010, 24, 401–408. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Liposome Type | PC | PC/DOTAP | Chol/DOTAP | DC-Chol/DOPE |
|---|---|---|---|---|
| Particle size (nm) | 174.9 ± 4.7 | 162.4 ± 3.5 | 164.3 ± 4.0 | 174.8 ± 6.0 |
| PDI | 0.056 ± 0.019 | 0.133 ± 0.029 | 0.093 ± 0.025 | 0.122 ± 0.020 |
| Zeta Potential | −5.09 ± 1.3 | +37.4 ± 2.3 | +50.9 ± 2.7 | +34.2 ± 2.5 |
| Target | Forward (5′~3′) | Reverse (5′~3′) |
|---|---|---|
| GAPDH | CATGAGAAGTATGACAACAGCCT | AGTCCTTCCACGATACCAAACT |
| FAS | CCACTGAAGAGCCTGGAAGA | GTAGTCAGCACCCAAGTCCT |
| FABP4 | AGTGAAAACTTCGATGATTACATGAA | GCCTGCCACTTTCCTTGTG |
| OPG | CCTTGCCCTGACCACTCTTAT | CACACACTCGGTTGTGGGT |
| RANKL | CGCTCTGTTCCTGTACTTTCG | GAGTCCTGCAAATCTGCGTT |
| COX-2 | CAGCCAGGCAGCAAATCC | ACATTCCCCACGGTTTTGAC |
| MMp-3 | GGCCTGGAACAGTCTTGGC | TGTCCATCGTTCATCATCGTCA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.-F.; Yeh, C.-C.; Chen, P.-J.; Chang, H.-I. The Impact of Lipid Types and Liposomal Formulations on Osteoblast Adiposity and Mineralization. Molecules 2018, 23, 95. https://doi.org/10.3390/molecules23010095
Chang S-F, Yeh C-C, Chen P-J, Chang H-I. The Impact of Lipid Types and Liposomal Formulations on Osteoblast Adiposity and Mineralization. Molecules. 2018; 23(1):95. https://doi.org/10.3390/molecules23010095
Chicago/Turabian StyleChang, Shun-Fu, Chih-Chang Yeh, Pin-Jyun Chen, and Hsin-I Chang. 2018. "The Impact of Lipid Types and Liposomal Formulations on Osteoblast Adiposity and Mineralization" Molecules 23, no. 1: 95. https://doi.org/10.3390/molecules23010095
APA StyleChang, S.-F., Yeh, C.-C., Chen, P.-J., & Chang, H.-I. (2018). The Impact of Lipid Types and Liposomal Formulations on Osteoblast Adiposity and Mineralization. Molecules, 23(1), 95. https://doi.org/10.3390/molecules23010095