Natural Negative Allosteric Modulators of 5-HT3 Receptors


Abstract
:1. Introduction
2. 5-hydroxytryptamine3 Receptors
3. 5-hydroxytryptamine3 Receptor-Mediated Nausea and Vomiting
4. Natural Negative Allosteric Modulators of 5-hydroxytryptamine3 Receptors
4.1. Terpenes
4.2. Ginger Constituents
4.3. Capsaicin
4.4. Eugenol and Vanilin
4.5. Thujone
4.6. Cannabidiol
4.7. Other Compounds
Author Contributions
Funding
Conflicts of Interest
References
- Janelsins, M.C.; Tejani, M.A.; Kamen, C.; Peoples, A.R.; Mustian, K.M.; Morrow, G.R. Current pharmacotherapy for chemotherapy-induced nausea and vomiting in cancer patients. Expert. Opin. Pharmacother. 2013, 14, 757–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haniadka, R.; Popouri, S.; Palatty, P.L.; Arora, R.; Baliga, M.S. Medicinal plants as antiemetics in the treatment of cancer: A review. Integr. Cancer Ther. 2012, 11, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Navari, R.M. Management of chemotherapy-induced nausea and vomiting: focus on newer agents and new uses for older agents. Drugs 2013, 73, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Hesketh, P.J.; Kris, M.G.; Basch, E.; Bohlke, K.; Barbour, S.Y.; Clark-Snow, R.A.; Danso, M.A.; Dennis, K.; Dupuis, L.L.; Dusetzina, S.B.; et al. Antiemetics: American Society of Clinical Oncology Clinical Practice Guideline Update. J. Clin. Oncol. 2017, 35, 3240–3261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmore, J.; D’Amato, S.; Griffith, N.; Schwartzberg, L. Recent advances in antiemetics: new formulations of 5HT3-receptor antagonists. Cancer Manag. Res. 2018, 10, 1827–1857. [Google Scholar] [CrossRef]
- Rojas, C.; Li, Y.; Zhang, J.; Stathis, M.; Alt, J.; Thomas, A.G. The antiemetic 5-HT3 receptor antagonist Palonosetron inhibits substance P-mediated responses in vitro and in vivo. J. Pharmacol. Exp. Ther. 2010, 335, 362–368. [Google Scholar] [CrossRef]
- Saito, M.; Aogi, K.; Sekine, I.; Yoshizawa, H.; Yanagita, Y.; Sakai, H. Palonosetron plus dexamethasone versus granisetron plus dexamethasone for prevention of nausea and vomiting during chemotherapy: a double-blind, double-dummy, randomised, comparative phase III trial. Lancet. Oncol. 2009, 10, 115–124. [Google Scholar] [CrossRef]
- Curran, M.P.; Robinson, D.M. Aprepitant: a review of its use in the prevention of nausea and vomiting. Drugs 2009, 69, 1853–1878. [Google Scholar] [CrossRef]
- Basch, E.; Prestrud, A.A.; Hesketh, P.J.; Kris, M.G.; Feyer, P.C.; Somerfield, M.R.; Chesney, M.; Clark-Snow, R.A.; Flaherty, A.M.; Freundlich, B.; et al. Antiemetics: American Society of Clinical Oncology clinical practice guideline update. J. Clin. Oncol. 2011, 29, 4189–4198. [Google Scholar] [CrossRef]
- Smith, H.S.; Cox, L.R.; Smith, E.J. 5-HT3 receptor antagonists for the treatment of nausea/vomiting. Ann. Palliat. Med. 2012, 1, 115–120. [Google Scholar]
- Johnston, K.D.; Lu, Z.; Rudd, J.A. Looking beyond 5-HT(3) receptors: a review of the wider role of serotonin in the pharmacology of nausea and vomiting. Eur. J. Pharmacol. 2014, 722, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Walstab, J.; Rappold, G.; Niesler, B. 5-HT(3) receptors: role in disease and target of drugs. Pharmacol. Ther. 2010, 128, 146–169. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Lummis, S.C. Antimalarial drugs inhibit human 5-HT(3) and GABA(A) but not GABA(C) receptors. Br. J. Pharmacol. 2008, 153, 1686–1696. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, B.S.; Lehmann, R.; Thiel, U.; Ziemba, P.M.; Beltran, L.R.; Sherkheli, M.A. Direct action and modulating effect of (+)- and (−)-nicotine on ion channels expressed in trigeminal sensory neurons. Eur. J. Pharmacol. 2014, 728, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Walstab, J.; Wohlfarth, C.; Hovius, R.; Schmitteckert, S.; Roth, R.; Lasitschka, F. Natural compounds boldine and menthol are antagonists of human 5-HT3 receptors: implications for treating gastrointestinal disorders. Neurogastroenterol. Motil. 2014, 26, 810–820. [Google Scholar] [CrossRef] [PubMed]
- De Leon, A. Palonosetron (Aloxi): A second-generation 5-HT(3) receptor antagonist for chemotherapy-induced nausea and vomiting. Proc. (Bayl. Univ. Med. Cent.) 2006, 19, 413–416. [Google Scholar] [CrossRef]
- Sheikhi, M.A.; Ebadi, A.; Talaeizadeh, A.; Rahmani, H. Alternative Methods to Treat Nausea and Vomiting from Cancer Chemotherapy. Chemother. Res. Pract. 2015, 2015. [Google Scholar] [CrossRef]
- Uteshev, V.V. The therapeutic promise of positive allosteric modulation of nicotinic receptors. Eur. J. Pharmacol. 2014, 727, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Davies, P.A. Allosteric modulation of the 5-HT(3) receptor. Curr. Opin. Pharmacol. 2011, 11, 75–80. [Google Scholar] [CrossRef]
- Newman, A.S.; Batis, N.; Grafton, G.; Caputo, F.; Brady, C.A.; Lambert, J.J. 5-Chloroindole: a potent allosteric modulator of the 5-HT(3) receptor. Br. J. Pharmacol. 2013, 169, 1228–1238. [Google Scholar] [CrossRef]
- Williams, D.K.; Wang, J.; Papke, R.L. Positive allosteric modulators as an approach to nicotinic acetylcholine receptor-targeted therapeutics: Advantages and limitations. Biochem. Pharmacol. 2011, 82, 915–930. [Google Scholar] [CrossRef] [Green Version]
- Barnes, N.M.; Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 1999, 38, 1083–1152. [Google Scholar] [CrossRef]
- Boess, F.G.; Lummis, S.C.; Martin, I.L. Molecular properties of 5-hydroxytryptamine3 receptor-type binding sites purified from NG108-15 cells. J. Neurochem. 1992, 59, 1692–1701. [Google Scholar] [CrossRef] [PubMed]
- Green, T.; Stauffer, K.A.; Lummis, S.C. Expression of recombinant homo-oligomeric 5-hydroxytryptamine3 receptors provides new insights into their maturation and structure. J. Biol. Chem. 1995, 270, 6056–6061. [Google Scholar] [CrossRef] [PubMed]
- Lummis, SC.R. 5-HT(3) Receptors. J. Biolog. Chem. 2012, 287, 40239–40245. [Google Scholar] [CrossRef] [PubMed]
- Hassaine, G.; Deluz, C.; Grasso, L.; Wyss, R.; Tol, M.B.; Hovius, R. X-ray structure of the mouse serotonin 5-HT3 receptor. Nature 2014, 512, 276–281. [Google Scholar] [CrossRef]
- Engel, M.; Smidt, M.P.; van Hooft, J.A. The serotonin 5-HT3 receptor: a novel neurodevelopmental target. Front Cell Neurosci. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Yang, J. Ion permeation through 5-hydroxytryptamine-gated channels in neuroblastoma N18 cells. J. Gen. Physiol. 1990, 96, 1177–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maricq, A.V.; Peterson, A.S.; Brake, A.J.; Myers, R.M.; Julius, D. Primary structure and functional expression of the 5HT3 receptor, a serotonin-gated ion channel. Science 1991, 254, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Derkach, V.; Surprenant, A.; North, R.A. 5-HT3 receptors are membrane ion channels. Nature 1989, 339, 706–709. [Google Scholar] [CrossRef]
- Yakel, J.L.; Jackson, M.B. 5-HT3 receptors mediate rapid responses in cultured hippocampus and a clonal cell line. Neuron 1988, 1, 615–621. [Google Scholar] [CrossRef]
- Riering, K.; Rewerts, C.; Zieglgansberger, W. Analgesic effects of 5-HT3 receptor antagonists. Scand. J. Rheumatol. Suppl. 2004, 119, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Faerber, L.; Drechsler, S.; Ladenburger, S.; Gschaidmeier, H.; Fischer, W. The neuronal 5-HT3 receptor network after 20 years of research--evolving concepts in management of pain and inflammation. Eur. J. Pharmacol. 2007, 560, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Machu, T.K. Therapeutics of 5-HT3 receptor antagonists: current uses and future directions. Pharmacol. Ther. 2011, 130, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Navari, R.M. 5-HT3 receptors as important mediators of nausea and vomiting due to chemotherapy. Biochim. Biophys. Acta 2015, 1848, 2738–2746. [Google Scholar] [CrossRef] [PubMed]
- Galligan, J.J. Ligand-gated ion channels in the enteric nervous system. Neurogastroenterol. Motil. 2002, 14, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Lummis, S.C. The 5-HT3 receptor as a therapeutic target. Expert Opin. Ther. Targets 2007, 11, 527–540. [Google Scholar] [CrossRef]
- Rapoport, B.L. Delayed Chemotherapy-Induced Nausea and Vomiting: Pathogenesis, Incidence, and Current Management. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef]
- Andrews, P.L.; Sanger, G.J. Nausea and the quest for the perfect anti-emetic. Eur. J. Pharmacol. 2014, 722, 108–121. [Google Scholar] [CrossRef]
- Eccles, R. Menthol and related cooling compounds. J. Pharm. Pharmacol. 1994, 46, 618–630. [Google Scholar] [CrossRef]
- Patel, T.; Ishiuji, Y.; Yosipovitch, G. Menthol: A refreshing look at this ancient compound. J. Am. Acad. Dermatol. 2007, 57, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Oz, M.; El Nebrisi, E.G.; Yang, K.S.; Howarth, F.C.; Al Kury, L.T. Cellular and Molecular Targets of Menthol Actions. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashoor, A.; Nordman, J.C.; Veltri, D.; Yang, K.H.; Shuba, Y.; Al Kury, L.T. Menthol inhibits 5-HT3 receptor-mediated currents. J. Pharmacol. Exp. Ther. 2013, 347, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Heimes, K.; Hauk, F.; Verspohl, E.J. Mode of action of peppermint oil and (−)-menthol with respect to 5-HT3 receptor subtypes: binding studies, cation uptake by receptor channels and contraction of isolated rat ileum. Phytother. Res. 2011, 25, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Ziemba, P.M.; Schreiner, B.S.; Flegel, C.; Herbrechter, R.; Stark, T.D.; Hofmann, T. Activation and modulation of recombinantly expressed serotonin receptor type 3A by terpenes and pungent substances. Biochem. Biophys. Res. Commun. 2015, 467, 1090–1096. [Google Scholar] [CrossRef]
- Ashoor, A.; Nordman, J.C.; Veltri, D.; Yang, K.H.; Al Kury, L.; Shuba, Y. Menthol binding and inhibition of alpha7-nicotinic acetylcholine receptors. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Hans, M.; Wilhelm, M.; Swandulla, D. Menthol Suppresses Nicotinic Acetylcholine Receptor Functioning in Sensory Neurons via Allosteric Modulation. Chem. Senses 2012, 37, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Ton, H.T.; Smart, A.E.; Aguilar, B.L.; Olson, T.T.; Kellar, K.J.; Ahern, G.P. Menthol Enhances the Desensitization of Human alpha3beta4 Nicotinic Acetylcholine Receptors. Mol. Pharmacol. 2015, 88, 256–264. [Google Scholar] [CrossRef]
- McKemy, D.D.; Neuhausser, W.M.; Julius, D. Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 2002, 416, 52–58. [Google Scholar] [CrossRef]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M. A TRP channel that senses cold stimuli and menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef]
- Hall, A.C.; Turcotte, C.M.; Betts, B.A.; Yeung, W.Y.; Agyeman, A.S.; Burk, L.A. Modulation of human GABAA and glycine receptor currents by menthol and related monoterpenoids. Eur. J. Pharmacol. 2004, 506, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Lau, B.K.; Karim, S.; Goodchild, A.K.; Vaughan, C.W.; Drew, G.M. Menthol enhances phasic and tonic GABAA receptor-mediated currents in midbrain periaqueductal grey neurons. Br. J. Pharmacol. 2014, 171, 2803–2813. [Google Scholar] [CrossRef] [PubMed]
- Tani, M.; Onimaru, H.; Ikeda, K.; Kawakami, K.; Homma, I. Menthol inhibits the respiratory rhythm in brainstem preparations of the newborn rats. Neuroreport. 2010, 21, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.B.; Jiang, P.; Gong, N.; Hu, X.L.; Fei, D.; Xiong, Z.Q. A-type GABA receptor as a central target of TRPM8 agonist menthol. PLoS ONE 2008, 3. [Google Scholar] [CrossRef] [PubMed]
- Turina, A.V.; Nolan, M.V.; Zygadlo, J.A.; Perillo, M.A. Natural terpenes: self-assembly and membrane partitioning. Biophys. Chem. 2006, 122, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Pertz, H.H.; Lehmann, J.; Roth-Ehrang, R.; Elz, S. Effects of ginger constituents on the gastrointestinal tract: role of cholinergic M3 and serotonergic 5-HT3 and 5-HT4 receptors. Planta Med. 2011, 77, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Rehman, M.U.; Amin, I.; Arif, A.; Rasool, S.; Bhat, S.A. A Review on Pharmacological Properties of Zingerone (4-(4-Hydroxy-3-methoxyphenyl)-2-butanone). Sci. World J. 2015, 2015. [Google Scholar] [CrossRef]
- Jin, Z.; Lee, G.; Kim, S.; Park, C.S.; Park, Y.S.; Jin, Y.H. Ginger and its pungent constituents non-competitively inhibit serotonin currents on visceral afferent neurons. Korean J. Physiol. Pharmacol. 2014, 18, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Tramer, M.R.; Carroll, D.; Campbell, F.A.; Reynolds, D.J.; Moore, R.A.; McQuay, H.J. Cannabinoids for control of chemotherapy induced nausea and vomiting: quantitative systematic review. BMJ 2001, 323, 16–21. [Google Scholar] [CrossRef]
- Oz, M.; Lozon, Y.; Sultan, A.; Yang, K.H.; Galadari, S. Effects of monoterpenes on ion channels of excitable cells. Pharmacol. Ther. 2015, 152, 83–97. [Google Scholar] [CrossRef]
- Amato, A.; Serio, R.; Mule, F. Involvement of cholinergic nicotinic receptors in the menthol-induced gastric relaxation. Eur. J. Pharmacol. 2014, 745, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Ruskin, D.N.; Anand, R.; LaHoste, G.J. Chronic menthol attenuates the effect of nicotine on body temperature in adolescent rats. Nicotine Tob. Res. 2008, 10, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- Brody, A.L.; Mukhin, A.G.; La Charite, J.; Ta, K.; Farahi, J.; Sugar, C.A. Up-regulation of nicotinic acetylcholine receptors in menthol cigarette smokers. Int. J. Neuropsychopharmacol. 2013, 16, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Gaudioso, C.; Hao, J.; Martin-Eauclaire, M.F.; Gabriac, M.; Delmas, P. Menthol pain relief through cumulative inactivation of voltage-gated sodium channels. Pain 2012, 153, 473–484. [Google Scholar] [CrossRef]
- Hawthorn, M.; Ferrante, J.; Luchowski, E.; Rutledge, A.; Wei, X.Y.; Triggle, D.J. The actions of peppermint oil and menthol on calcium channel dependent processes in intestinal, neuronal and cardiac preparations. Aliment. Pharmacol. Ther. 1988, 2, 101–118. [Google Scholar] [CrossRef]
- Sidell, N.; Verity, M.A.; Nord, E.P. Menthol blocks dihydropyridine-insensitive Ca2+ channels and induces neurite outgrowth in human neuroblastoma cells. J. Cell Physiol. 1990, 142, 410–419. [Google Scholar] [CrossRef]
- Cheang, W.S.; Lam, M.Y.; Wong, W.T.; Tian, X.Y.; Lau, C.W.; Zhu, Z. Menthol relaxes rat aortae, mesenteric and coronary arteries by inhibiting calcium influx. Eur. J. Pharmacol. 2013, 702, 79–84. [Google Scholar] [CrossRef]
- Wondergem, R.; Bartley, J.W. Menthol increases human glioblastoma intracellular Ca2+, BK channel activity and cell migration. J. Biomed. Sci. 2009, 16. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Hwang, S.W.; Miyamoto, T.; Dubin, A.E.; Patapoutian, A.; Story, G.M. More than cool: promiscuous relationships of menthol and other sensory compounds. Mol. Cell Neurosci. 2006, 32, 335–343. [Google Scholar] [CrossRef]
- Parnas, M.; Peters, M.; Dadon, D.; Lev, S.; Vertkin, I.; Slutsky, I. Carvacrol is a novel inhibitor of Drosophila TRPL and mammalian TRPM7 channels. Cell Calcium 2009, 45, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Iturriaga-Vasquez, P.; Perez, E.G.; Slater, E.Y.; Bermudez, I.; Cassels, B.K. Aporphine metho salts as neuronal nicotinic acetylcholine receptor blockers. Bioorg. Med. Chem. 2007, 15, 3368–3372. [Google Scholar] [CrossRef] [PubMed]
- Stotz, S.C.; Vriens, J.; Martyn, D.; Clardy, J.; Clapham, D.E. Citral sensing by Transient [corrected] receptor potential channels in dorsal root ganglion neurons. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Ferreira-da-Silva, F.W.; da Silva-Alves, K.S.; Alves-Fernandes, T.A.; Coelho-de-Souza, A.N.; Leal-Cardoso, J.H. Effects of 1,8-cineole on Na(+) currents of dissociated superior cervical ganglia neurons. Neurosci. Lett. 2015, 595, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, H.J.; Germann, T.; Gillen, C.; Hatt, H.; Jostock, R. Characterization of the mouse cold-menthol receptor TRPM8 and vanilloid receptor type-1 VR1 using a fluorometric imaging plate reader (FLIPR) assay. Br. J. Pharmacol. 2004, 141, 737–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherkheli, M.A.; Benecke, H.; Doerner, J.F.; Kletke, O.; Vogt-Eisele, A.K.; Gisselmann, G. Monoterpenoids induce agonist-specific desensitization of transient receptor potential vanilloid-3 (TRPV3) ion channels. J. Pharm. Pharm. Sci. 2009, 12, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Takaishi, M.; Fujita, F.; Uchida, K.; Yamamoto, S.; Sawada Shimizu, M.; Hatai Uotsu, C. 1,8-cineole, a TRPM8 agonist, is a novel natural antagonist of human TRPA1. Mol. Pain 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Leal-Cardoso, J.H.; da Silva-Alves, K.S.; Ferreira-da-Silva, F.W.; dos Santos-Nascimento, T.; Joca, H.C.; de Macedo, F.H. Linalool blocks excitability in peripheral nerves and voltage-dependent Na+ current in dissociated dorsal root ganglia neurons. Eur. J. Pharmacol. 2010, 645, 86–93. [Google Scholar] [CrossRef]
- Re, L.; Barocci, S.; Sonnino, S.; Mencarelli, A.; Vivani, C.; Paolucci, G. Linalool modifies the nicotinic receptor-ion channel kinetics at the mouse neuromuscular junction. Pharmacol. Res. 2000, 42, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Riera, C.E.; Menozzi-Smarrito, C.; Affolter, M.; Michlig, S.; Munari, C.; Robert, F. Compounds from Sichuan and Melegueta peppers activate, covalently and non-covalently, TRPA1 and TRPV1 channels. Br. J. Pharmacol. 2009, 157, 1398–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.X.; Tang, X.D.; Wang, F.Y.; Duan, Z.J.; Li, Y.C.; Qiu, J.J. Effect of gingerol on colonic motility via inhibition of calcium channel currents in rats. World J. Gastroenterol. 2015, 21, 13466–13472. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, S.; Ono, K.; Terawaki, K.; Matsumoto, C.; Mizuno, K.; Yamaguchi, K. [6]-gingerol and [6]-shogaol, active ingredients of the traditional Japanese medicine hangeshashinto, relief oral ulcerative mucositis-induced pain via action on Na+ channels. Pharmacol. Res. 2017, 117, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Beltran, L.R.; Dawid, C.; Beltran, M.; Gisselmann, G.; Degenhardt, K.; Mathie, K. The pungent substances piperine, capsaicin, 6-gingerol and polygodial inhibit the human two-pore domain potassium channels TASK-1, TASK-3 and TRESK. Front. Pharmacol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.; Li, H.Y.; Perez-Reyes, E.; Lee, J.H. Effects of eugenol on T-type Ca2+ channel isoforms. J. Pharmacol. Exp. Ther. 2013, 347, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Damiani, C.E.; Moreira, C.M.; Zhang, H.T.; Creazzo, T.L.; Vassallo, D.V. Effects of eugenol, an essential oil, on the mechanical and electrical activities of cardiac muscle. J. Cardiovasc. Pharmacol. 2004, 44, 688–695. [Google Scholar] [CrossRef]
- Ding, J.; Huang, C.; Peng, Z.; Xie, Y.; Deng, S.; Nie, Y.Z. Electrophysiological characterization of methyleugenol: A novel agonist of GABA(A) receptors. ACS Chem. Neurosci. 2014, 5, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Moon, J.Y.; Jung, S.J.; Kang, J.G.; Choi, S.P.; Jang, J.H. Eugenol inhibits the GABAA current in trigeminal ganglion neurons. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Chow, J.C.; Tsai, J.J.; Wu, S.N. Characterizing the effects of Eugenol on neuronal ionic currents and hyperexcitability. Psychopharmacology (Berl) 2012, 221, 575–587. [Google Scholar] [CrossRef]
- Li, H.Y.; Park, C.K.; Jung, S.J.; Choi, S.Y.; Lee, S.J.; Park, K. Eugenol inhibits K+ currents in trigeminal ganglion neurons. J. Dent. Res. 2007, 86, 898–902. [Google Scholar] [CrossRef]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef]
- Yang, B.H.; Piao, Z.G.; Kim, Y.B.; Lee, C.H.; Lee, J.K.; Park, K. Activation of vanilloid receptor 1 (VR1) by eugenol. J. Dent. Res. 2003, 82, 781–785. [Google Scholar] [CrossRef]
- Mahgoub, M.; Keun-Hang, S.Y.; Sydorenko, V.; Ashoor, A.; Kabbani, N.; Al Kury, L. Effects of cannabidiol on the function of alpha7-nicotinic acetylcholine receptors. Eur. J. Pharmacol. 2013, 720, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, J.; Demir, R.; Leuwer, M.; de la Roche, J.; Krampfl, K.; Foadi, N. The nonpsychotropic cannabinoid cannabidiol modulates and directly activates alpha-1 and alpha-1-Beta glycine receptor function. Pharmacology 2009, 83, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Cilio, M.R.; Cross, H.; Fernandez-Ruiz, J.; French, J.; Hill, C. Cannabidiol: pharmacology and potential therapeutic role in epilepsy and other neuropsychiatric disorders. Epilepsia 2014, 55, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Yang, K.S.; Isaev, D.; Nebrisi, E.E.; Syed, N.; Khan, N. Thujone inhibits the function of alpha7-nicotinic acetylcholine receptors and impairs nicotine-induced memory enhancement in one-trial passive avoidance paradigm. Toxicology 2017, 384, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Czyzewska, M.M.; Mozrzymas, J.W. Monoterpene alpha-thujone exerts a differential inhibitory action on GABA(A) receptors implicated in phasic and tonic GABAergic inhibition. Eur. J. Pharmacol. 2013, 702, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Sadraei, H.; Asghari, G.; Jalali, F. Assessment of hydroalcoholic and hexane extracts of Rosa persica Mich. flower on rat ileum spasm. Res. Pharm. Sci. 2016, 11, 160–167. [Google Scholar] [PubMed]
- Jarvis, G.E.; Barbosa, R.; Thompson, A.J. Noncompetitive Inhibition of 5-HT3 Receptors by Citral, Linalool, and Eucalyptol Revealed by Nonlinear Mixed-Effects Modeling. J. Pharmacol. Exp. Ther. 2016, 356, 549–562. [Google Scholar] [CrossRef]
- Hales, T.G.; Lambert, J.J. The actions of propofol on inhibitory amino acid receptors of bovine adrenomedullary chromaffin cells and rodent central neurones. Br. J. Pharmacol. 1991, 104, 619–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barann, M.; Dilger, J.P.; Bonisch, H.; Gothert, M.; Dybek, A.; Urban, B.W. Inhibition of 5-HT3 receptors by propofol: equilibrium and kinetic measurements. Neuropharmacology 2000, 39, 1064–1074. [Google Scholar] [CrossRef]
- Garcia, D.A.; Bujons, J.; Vale, C.; Sunol, C. Allosteric positive interaction of thymol with the GABAA receptor in primary cultures of mouse cortical neurons. Neuropharmacology 2006, 50, 25–35. [Google Scholar] [CrossRef]
- Lansdell, S.J.; Sathyaprakash, C.; Doward, A.; Millar, N.S. Activation of human 5-hydroxytryptamine type 3 receptors via an allosteric transmembrane site. Mol. Pharmacol. 2015, 87, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Leach, M.; Bradley, H. The effectiveness and safety of ginger for pregnancy-induced nausea and vomiting: a systematic review. Women Birth. 2013, 26, e26–e30. [Google Scholar] [CrossRef] [PubMed]
- Haniadka, R.; Saldanha, E.; Sunita, V.; Palatty, P.L.; Fayad, R.; Baliga, M.S. A review of the gastroprotective effects of ginger (Zingiber officinale Roscoe). Food Funct. 2013, 4, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Ernst, E.; Pittler, M.H. Efficacy of ginger for nausea and vomiting: a systematic review of randomized clinical trials. Br. J. Anaesth. 2000, 84, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Marx, W.M.; Teleni, L.; McCarthy, A.L.; Vitetta, L.; McKavanagh, D.; Thomson, D. Ginger (Zingiber officinale) and chemotherapy-induced nausea and vomiting: a systematic literature review. Nutr. Rev. 2013, 71, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aziz, H.; Windeck, T.; Ploch, M.; Verspohl, E.J. Mode of action of gingerols and shogaols on 5-HT3 receptors: binding studies, cation uptake by the receptor channel and contraction of isolated guinea-pig ileum. Eur. J. Pharmacol. 2006, 530, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Kinoshita, K.; Koyama, K.; Takahashi, K. Anti-emetic principles of Magnolia obovata bark and Zingiber officinale rhizome. Planta Med. 1994, 60, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kinoshita, K.; Koyama, K.; Takahashi, K.; Kondo, S.; Watanabe, K. Structure-antiemetic-activity of some diarylheptanoids and their analogues. Phytomedicine 2002, 9, 146–152. [Google Scholar] [CrossRef]
- Walstab, J.; Kruger, D.; Stark, T.; Hofmann, T.; Demir, I.E.; Ceyhan, G.O. Ginger and its pungent constituents non-competitively inhibit activation of human recombinant and native 5-HT3 receptors of enteric neurons. Neurogastroenterol. Motil. 2013, 25, 439–447. [Google Scholar] [CrossRef]
- Lohning, A.E.; Marx, W.; Isenring, L. In silico investigation into the interactions between murine 5-HT3 receptor and the principle active compounds of ginger (Zingiber officinale). J. Mol. Graph. Model. 2016, 70, 315–327. [Google Scholar] [CrossRef]
- Darmani, N.A.; Chebolu, S.; Zhong, W.; Trinh, C.; McClanahan, B.; Brar, R.S. Additive antiemetic efficacy of low-doses of the cannabinoid CB(1/2) receptor agonist Delta(9)-THC with ultralow-doses of the vanilloid TRPV1 receptor agonist resiniferatoxin in the least shrew (Cryptotis parva). Eur. J. Pharmacol. 2014, 722, 147–155. [Google Scholar] [CrossRef]
- Koo, M.S.; Kim, K.S.; Lee, H.J.; Jeong, J.S.; Lee, J.W. Antiemetic efficacy of capsicum plaster on acupuncture points in patients undergoing thyroid operation. Korean J. Anesthesiol. 2013, 65, 539–543. [Google Scholar] [CrossRef]
- Dezieck, L.; Hafez, Z.; Conicella, A.; Blohm, E.; O’Connor, M.J.; Schwarz, E.S. Resolution of cannabis hyperemesis syndrome with topical capsaicin in the emergency department: A case series. Clin. Toxicol. (Phila) 2017, 55, 908–913. [Google Scholar] [CrossRef]
- Pelissier, F.; Claudet, I.; Gandia-Mailly, P.; Benyamina, A.; Franchitto, N. Use of Capsaicin Cream in Cannabis Hyperemesis Syndrome. J. Emerg. Med. 2017, 52, 760–761. [Google Scholar] [CrossRef]
- Deiml, T.; Haseneder, R.; Zieglgansberger, W.; Rammes, G.; Eisensamer, B.; Rupprecht, R. Alpha-thujone reduces 5-HT3 receptor activity by an effect on the agonist-reduced desensitization. Neuropharmacology 2004, 46, 192–201. [Google Scholar] [CrossRef]
- Slatkin, N.E. Cannabinoids in the treatment of chemotherapy-induced nausea and vomiting: Beyond prevention of acute emesis. J. Support Oncol. 2007, 5, 1–9. [Google Scholar]
- Izzo, A.A.; Borrelli, F.; Capasso, R.; Di Marzo, V.; Mechoulam, R. Non-psychotropic plant cannabinoids: New therapeutic opportunities from an ancient herb. Trends Pharmacol. Sci. 2009, 30, 515–527. [Google Scholar] [CrossRef]
- Mechoulam, R.; Peters, M.; Murillo-Rodriguez, E.; Hanus, L.O. Cannabidiol--recent advances. Chem. Biodivers. 2007, 4, 1678–1692. [Google Scholar] [CrossRef]
- Pertwee, R.G. Emerging strategies for exploiting cannabinoid receptor agonists as medicines. Br. J. Pharmacol. 2009, 156, 397–411. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, C.; Filippis, D.D.; Iuvone, T.; Blasio, A.; Steardo, A.; Esposito, G. Cannabidiol in medicine: a review of its therapeutic potential in CNS disorders. Phytother. Res. 2009, 23, 597–602. [Google Scholar] [CrossRef]
- Yang, K.H.; Galadari, S.; Isaev, D.; Petroianu, G.; Shippenberg, T.S.; Oz, M. The nonpsychoactive cannabinoid cannabidiol inhibits 5-hydroxytryptamine3A receptor-mediated currents in Xenopus laevis oocytes. J. Pharmacol. Exp. Ther. 2010, 333, 547–554. [Google Scholar] [CrossRef]
- Ueta, K.; Suzuki, T.; Sugimoto, M.; Uchida, I.; Mashimo, T. Local anesthetics have different mechanisms and sites of action at recombinant 5-HT3 receptors. Reg. Anesth. Pain Med. 2007, 32, 462–470. [Google Scholar] [CrossRef]


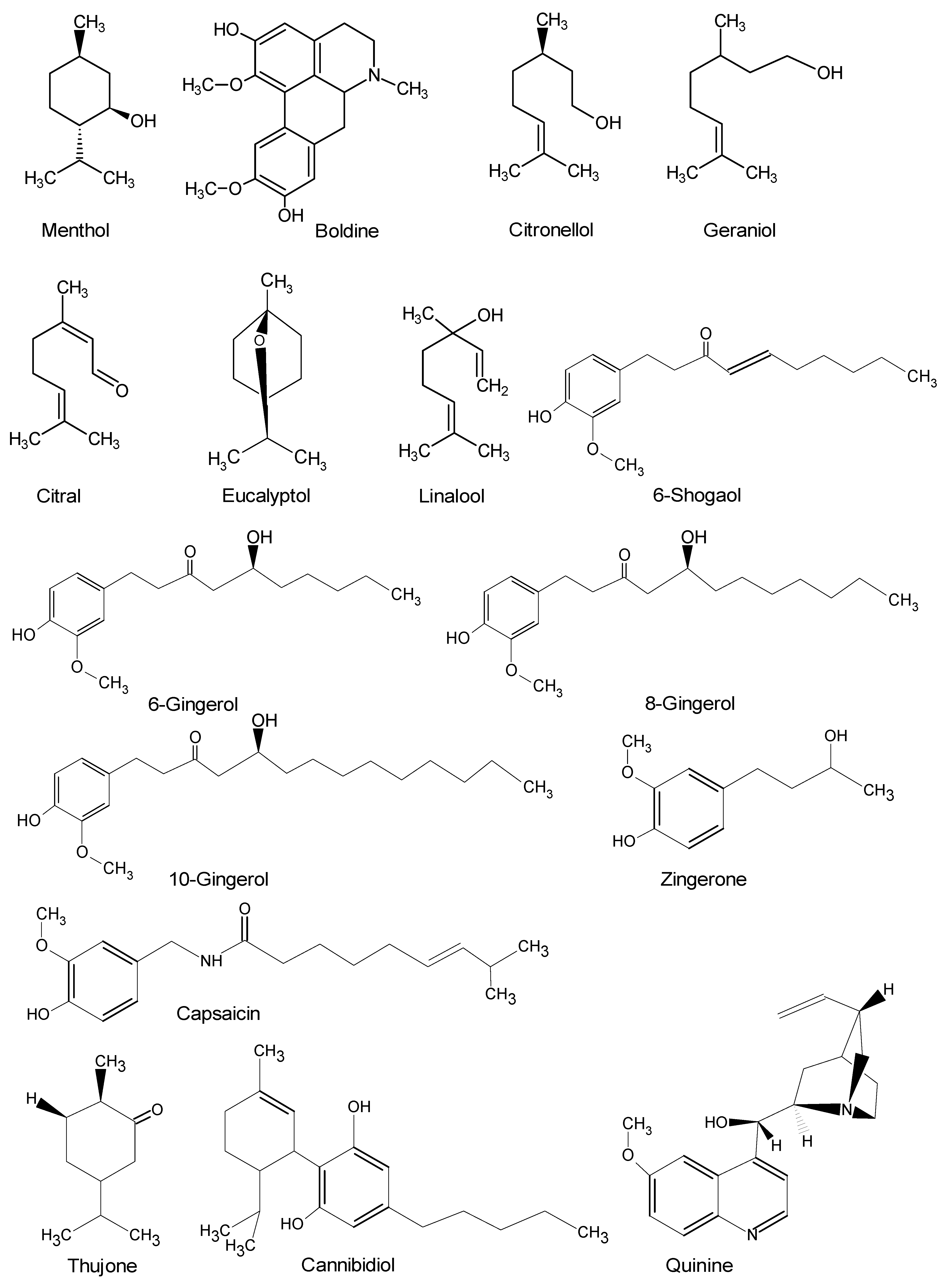{kind=link}
| Compound | Concentration | Preparation | References |
|---|---|---|---|
| Menthol | >100 µM | [14C] guanidinium influx into N1E-115 cells. Isotonic contractions of the ileum isolated rat. Equilibrium competition binding studies. | [44] |
| IC50 = 163 µM | Xenopus laevis oocytes | [43] | |
| IC50 = 4.75 µM IC50 = 489 µM | HEK293 cells Xenopus laevis oocytes | [15] | |
| [45] | |||
| Boldine | IC50 = 5.94 µM | HEK293 cells | [15] |
| Citronellol | IC50 = 64.3 µM | Xenopus laevis oocytes | [45] |
| Geraniol | IC50 = 188 µM | Xenopus laevis oocytes | [45] |
| Citral | IC50 = 120 µM | Xenopus laevis oocytes and HEK293 cells | [55] |
| Eucalyptol | IC50 = 258 µM | Xenopus laevis oocytes and HEK293 cells | [55] |
| Linalool | IC50 = 141 µM | Xenopus laevis oocytes and HEK293 cells | [55] |
| 6-shogaol | 100 µM IC50 = 3.2–10 µM | [14C] guanidinium influx into N1E-115 cells. Equilibrium competition binding studies. Isotonic contractions of the isolated guinea-pig ileum. Visceral afferent neurons and HEK293 cells. | [56,57] |
| 8-gingerol | 1–3 µM 100 µM | [14C] guanidinium influx into N1E-115 cells. Equilibrium competition binding studies. Isotonic contractions of the isolated guinea-pig ileum. | [56] |
| 6-gingerol | 88 µM 100 µM IC50 = 9–30 µM | [14C] guanidinium influx into N1E-115 cells. Equilibrium competition binding studies. Isotonic contractions of the isolated guinea-pig ileum. Visceral afferent neurons and HEK293 cells | [56,57,58] |
| 10-gingerol | IC50 = 9–15 µM | [14C] guanidinium influx into N1E-115 cells. Equilibrium competition binding studies. Isotonic contractions of the isolated guinea-pig ileum. | [56] |
| Zingerone | IC50 = 1.19 mM | Visceral afferent neurons | [58] |
| Capsaicin | IC50 = 98.1 µM | Xenopus laevis oocytes | [45] |
| Eugenol | IC50 = 1159 µM | Xenopus laevis oocytes | [45] |
| Vanillin | IC50 = 4744 µM | Xenopus laevis oocytes | [45] |
| Thujone | - | HEK293 cells | [59] |
| Cannabidiol | IC50 = 0.6 µM | Xenopus laevis oocytes | [60] |
| Quinine | IC50 = 1.06 µM | Xenopus laevis oocytes | [13] |
| Compound | Target protein | Effect | References |
|---|---|---|---|
| Menthol | GABAA | Potentiation | [52,53,54] |
| Glycine | Potentiation | [51] | |
| Nicotinic receptors | Reduction Up-regulation | [47,61,62] [63] | |
| TRP channels | Potentiation | [49,50] | |
| Na+ channels | Blocking | [64] | |
| Ca2+ channels | Inhibition | [65,66,67] | |
| K+ channels | Activation | [68] | |
| TRP channels | Activation Inhibition | [49,50,69] [70] | |
| Boldine | TRP channels | Inhibition | [71] |
| Citral | TRP channels | Activation | [72] |
| Eucalyptol | Na+ channels | Inhibition | [73] |
| TRP channels | Activation | [74,75,76] | |
| Linalool | Na+ channels | Inhibition | [77] |
| Nicotinic receptors | Reduction | [78] | |
| TRP channels | Activation | [74,79] | |
| Gingerol | L-type Ca2+ channels | Inhibition | [80] |
| 6-gingerol | Na+ channels | Blockage | [81] |
| K+ channels | Inhibition | [82] | |
| 6-shogaol | Na+ channels | Blockage | [81] |
| Capsaicin | K+ channels | Inhibition | [82] |
| Eugenol | T-type Ca2+ channel | Inhibition | [83,84] |
| GABAA receptors | Activation | [85,86] | |
| K+ channels | Inhibition | [87,88] | |
| Cannabidiol | TRP channels | Activation | [74,89,90] |
| α7-nicotinic receptors | Inhibition | [91] | |
| Glycine receptors | Activation | [92] | |
| Thujone | TRP channels | Activation | [93] |
| α7-nicotinic receptors | Inhibition | [94] | |
| GABAA receptors | Inhibition | [95] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Kury, L.T.; Mahgoub, M.; Howarth, F.C.; Oz, M. Natural Negative Allosteric Modulators of 5-HT3 Receptors. Molecules 2018, 23, 3186. https://doi.org/10.3390/molecules23123186
Al Kury LT, Mahgoub M, Howarth FC, Oz M. Natural Negative Allosteric Modulators of 5-HT3 Receptors. Molecules. 2018; 23(12):3186. https://doi.org/10.3390/molecules23123186
Chicago/Turabian StyleAl Kury, Lina T., Mohamed Mahgoub, Frank Christopher Howarth, and Murat Oz. 2018. "Natural Negative Allosteric Modulators of 5-HT3 Receptors" Molecules 23, no. 12: 3186. https://doi.org/10.3390/molecules23123186
APA StyleAl Kury, L. T., Mahgoub, M., Howarth, F. C., & Oz, M. (2018). Natural Negative Allosteric Modulators of 5-HT3 Receptors. Molecules, 23(12), 3186. https://doi.org/10.3390/molecules23123186