
Chemical Characterization and Hypoglycaemic Activities In Vitro of Two Polysaccharides from Inonotus obliquus by Submerged Culture


Abstract
:1. Introduction
2. Results
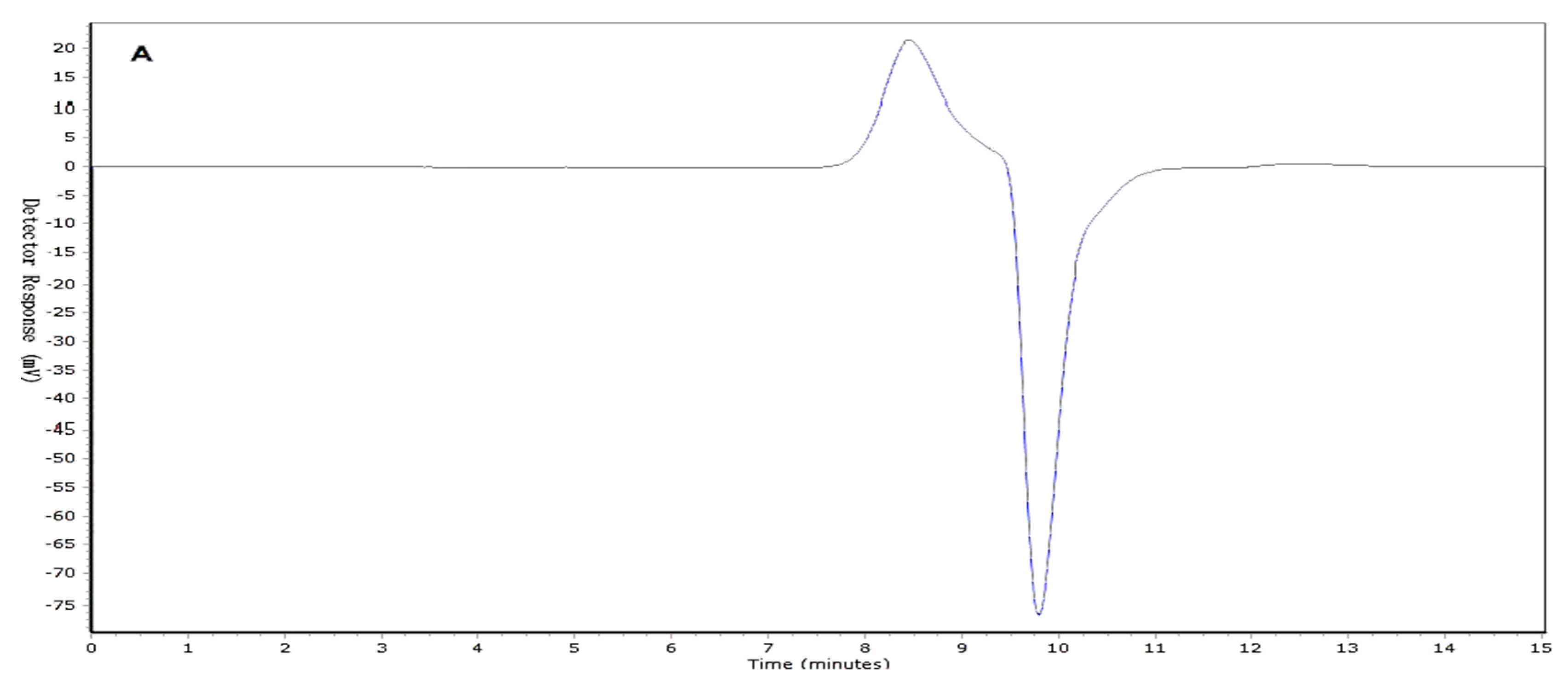
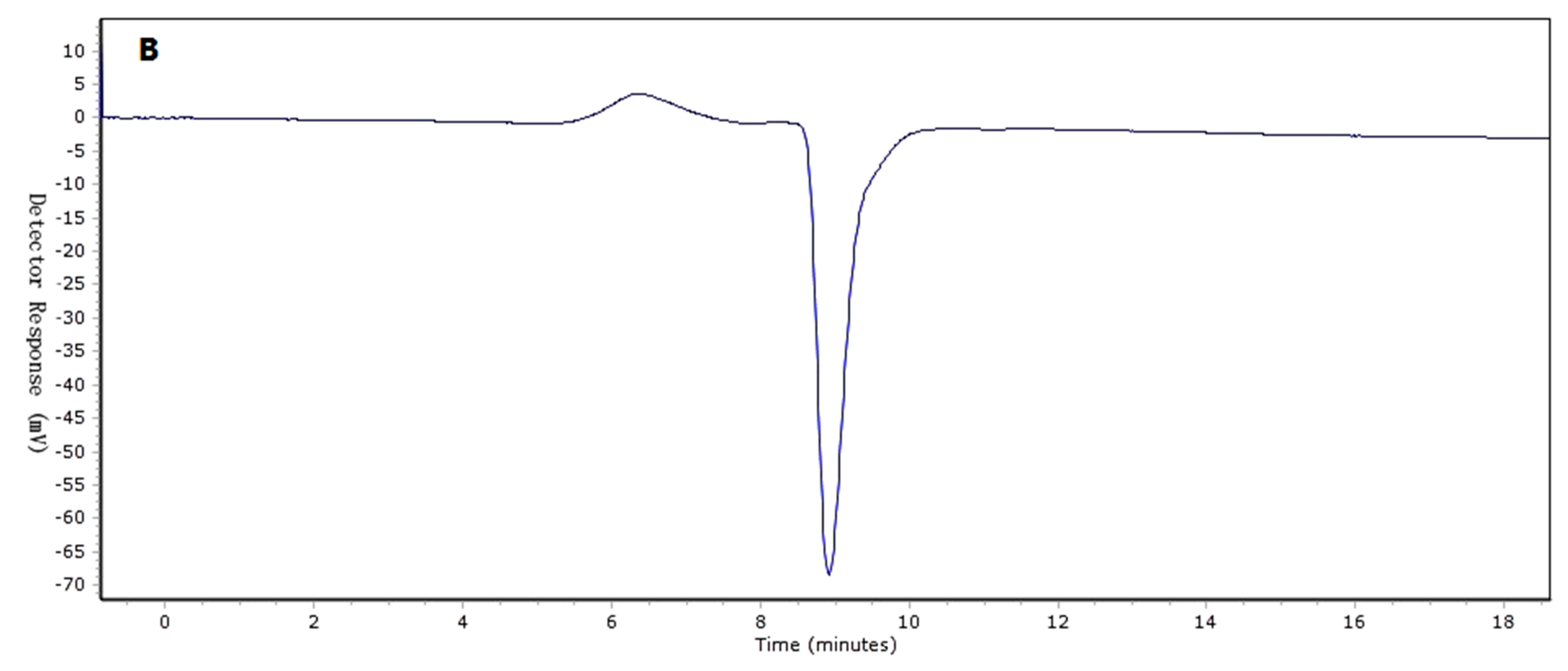
2.1. Determination of Molecular Weight (Mw) Distribution
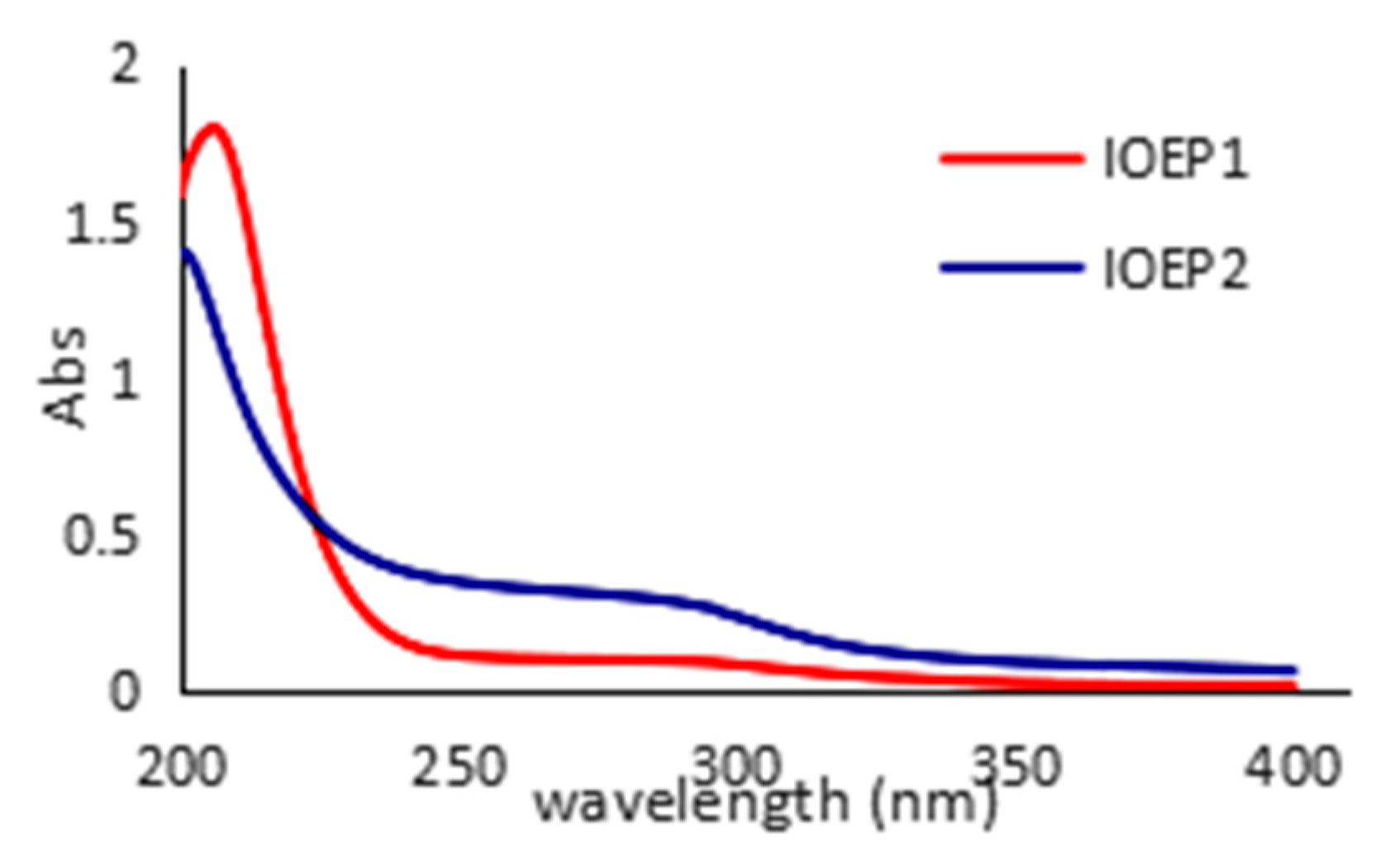
2.2. Analysis of Physicochemical Properties and Monosaccharide Compositions
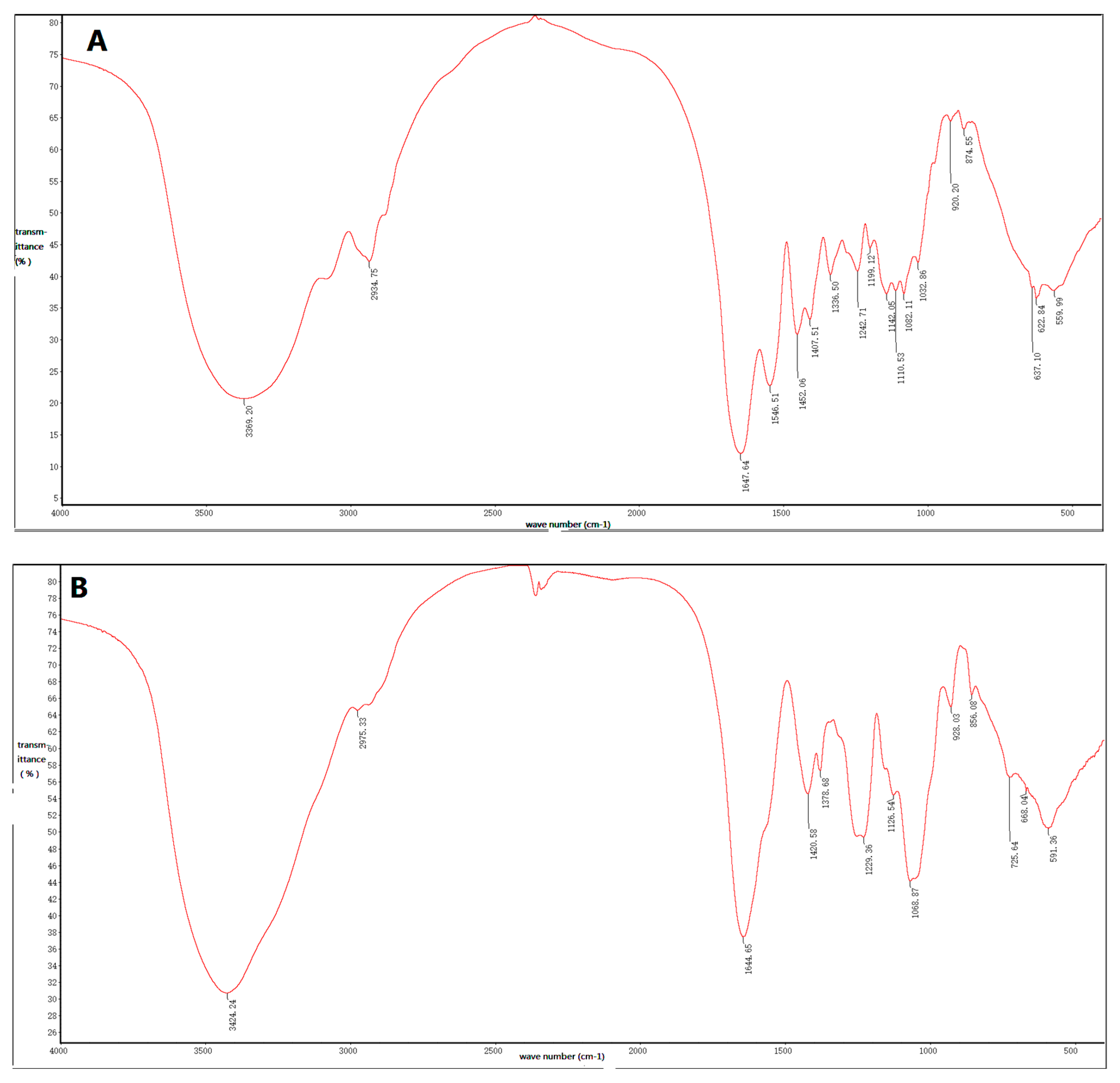
2.3. Fourier-Transform Infrared Spectroscopic Analysis
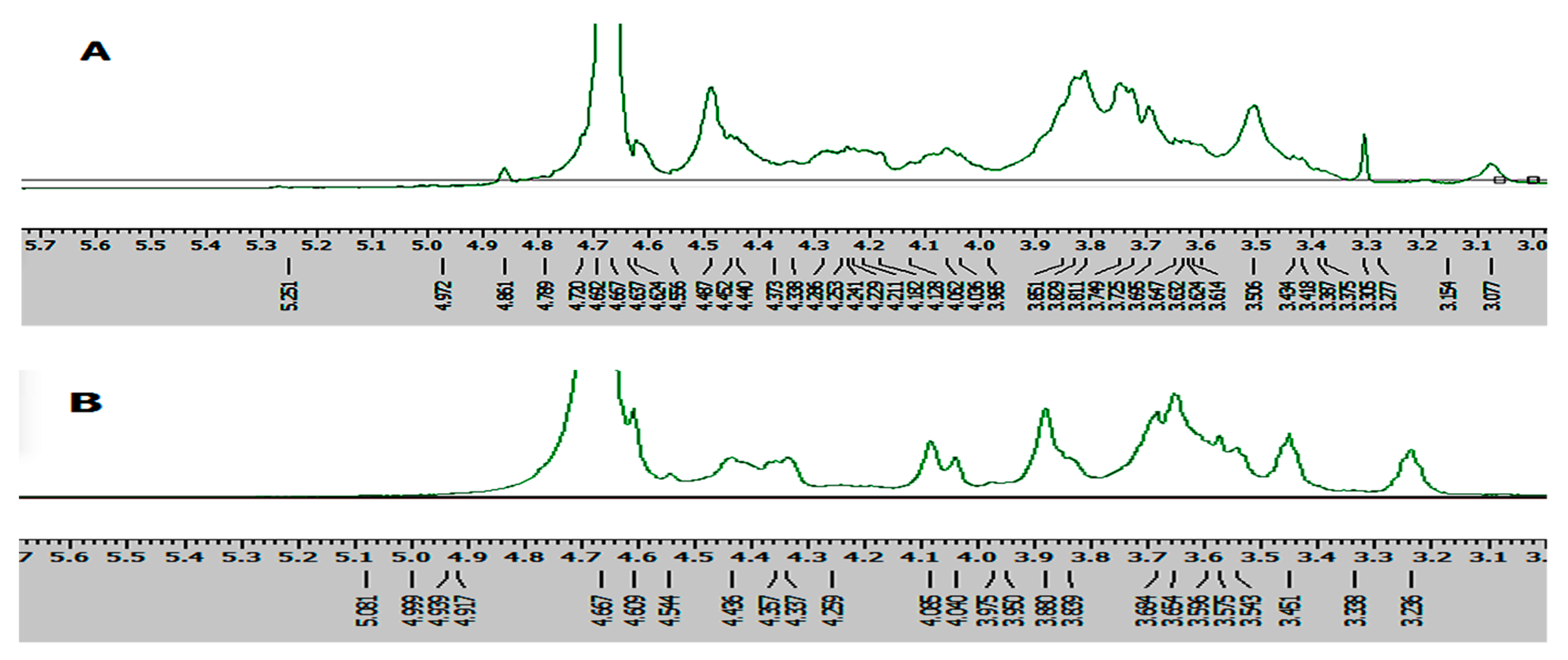
2.4. 1H-NMR Analysis
2.5. Hypoglycaemic Activities In Vitro
2.5.1. Cytotoxicity of IOEP1 and IOEP2 at Different Concentrations against HepG2 Cells
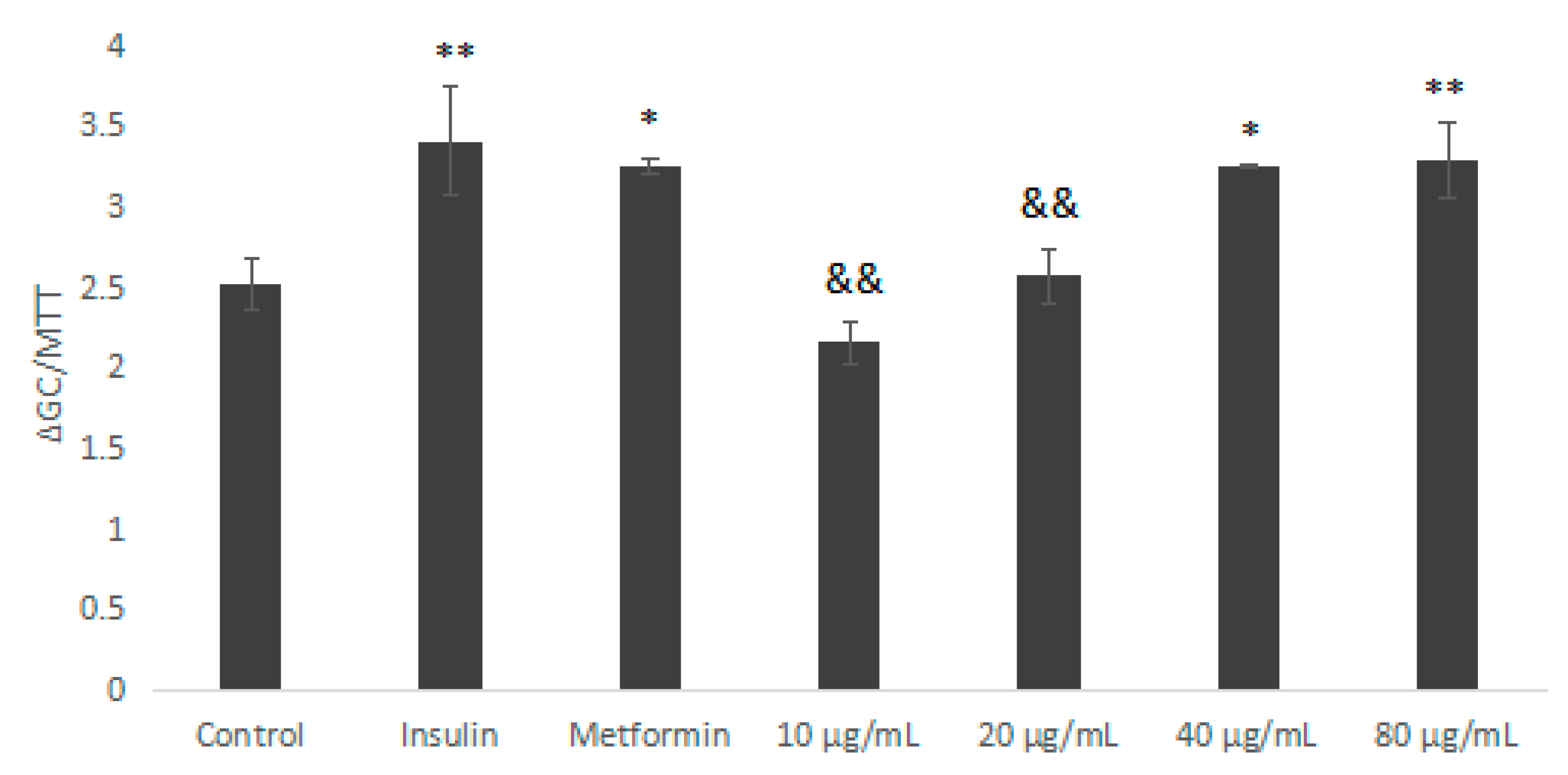
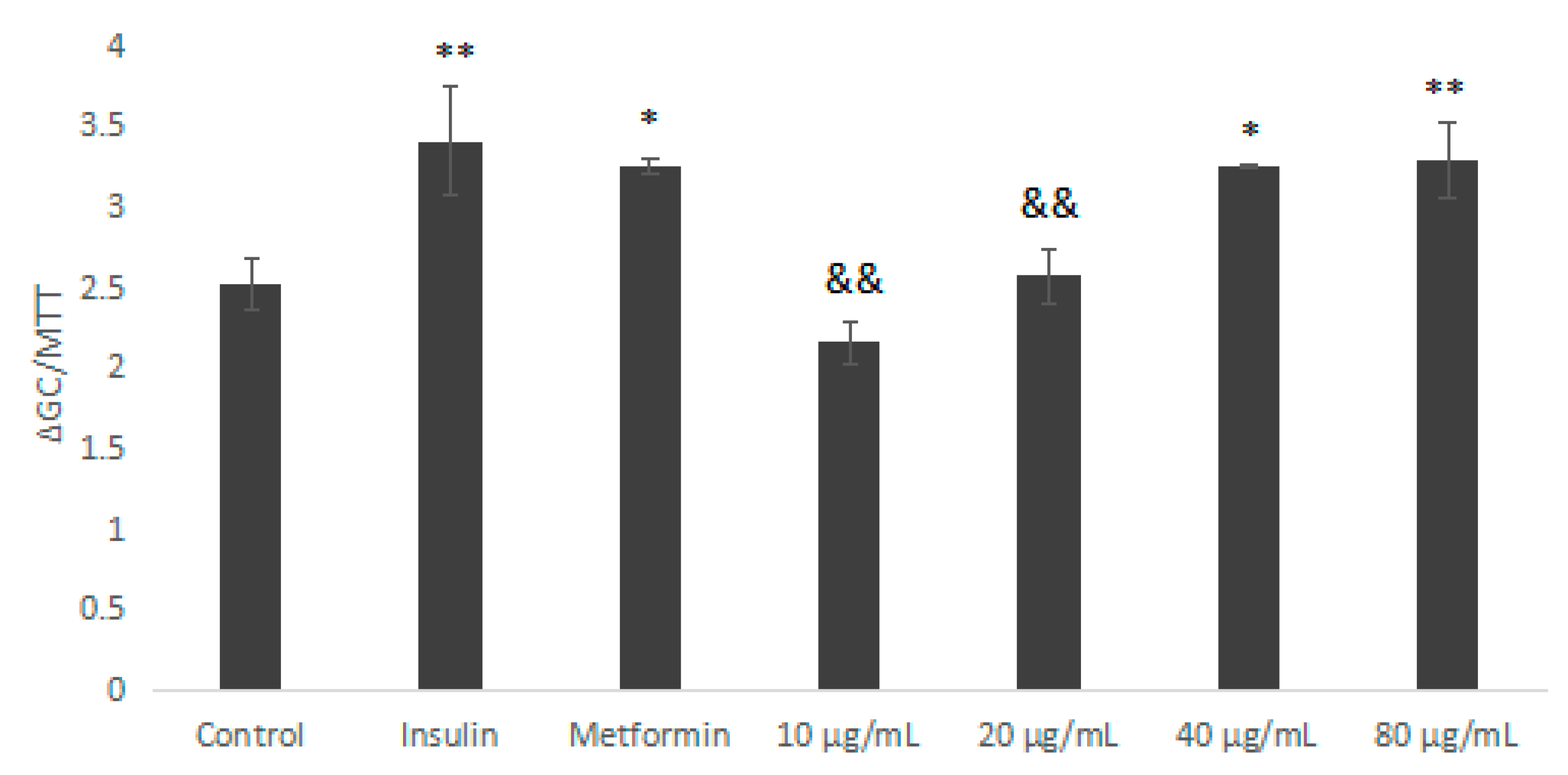
2.5.2. Glucose Consumption Experiment of IOEP1 and IOEP2 on HepG2 Cells
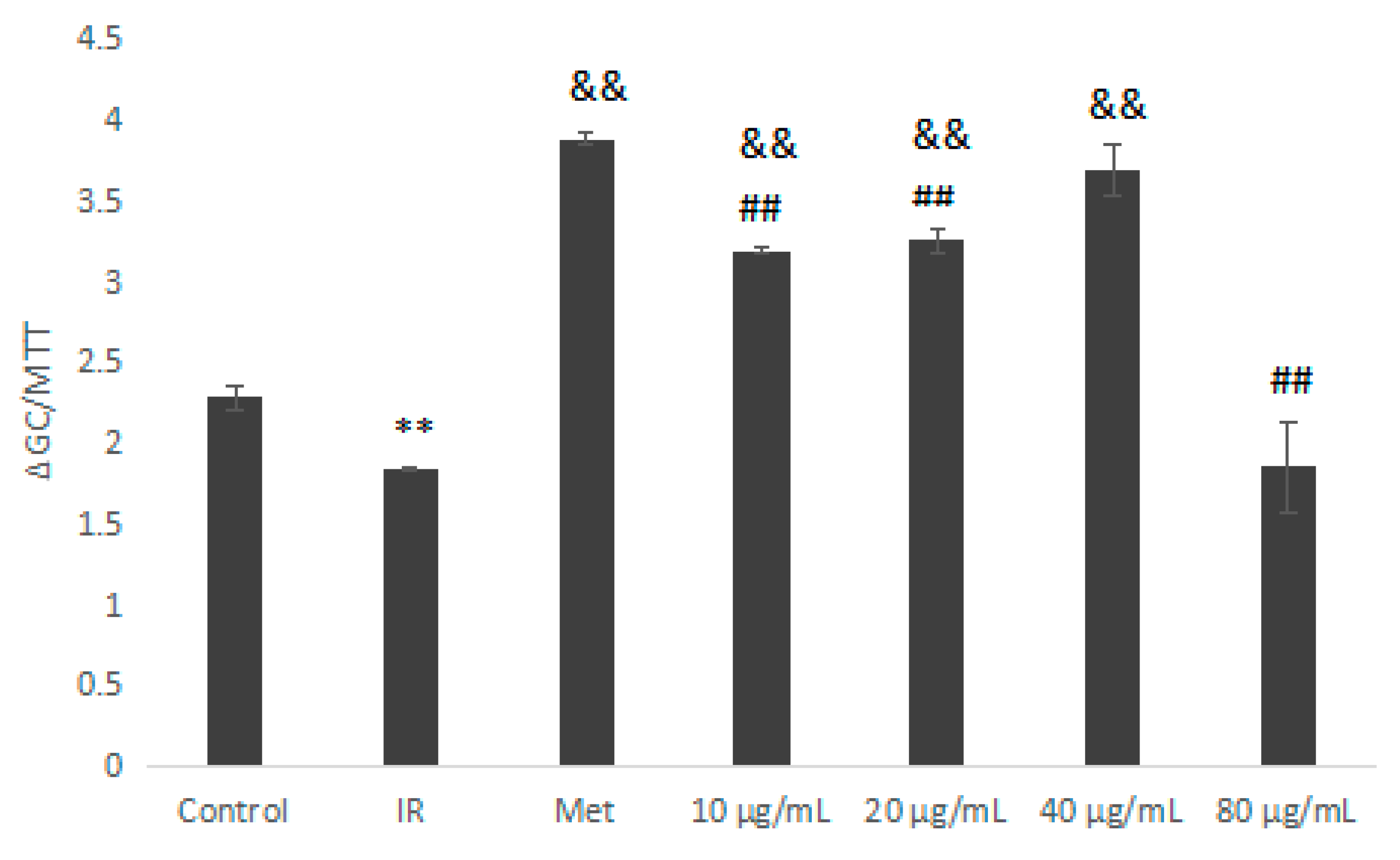
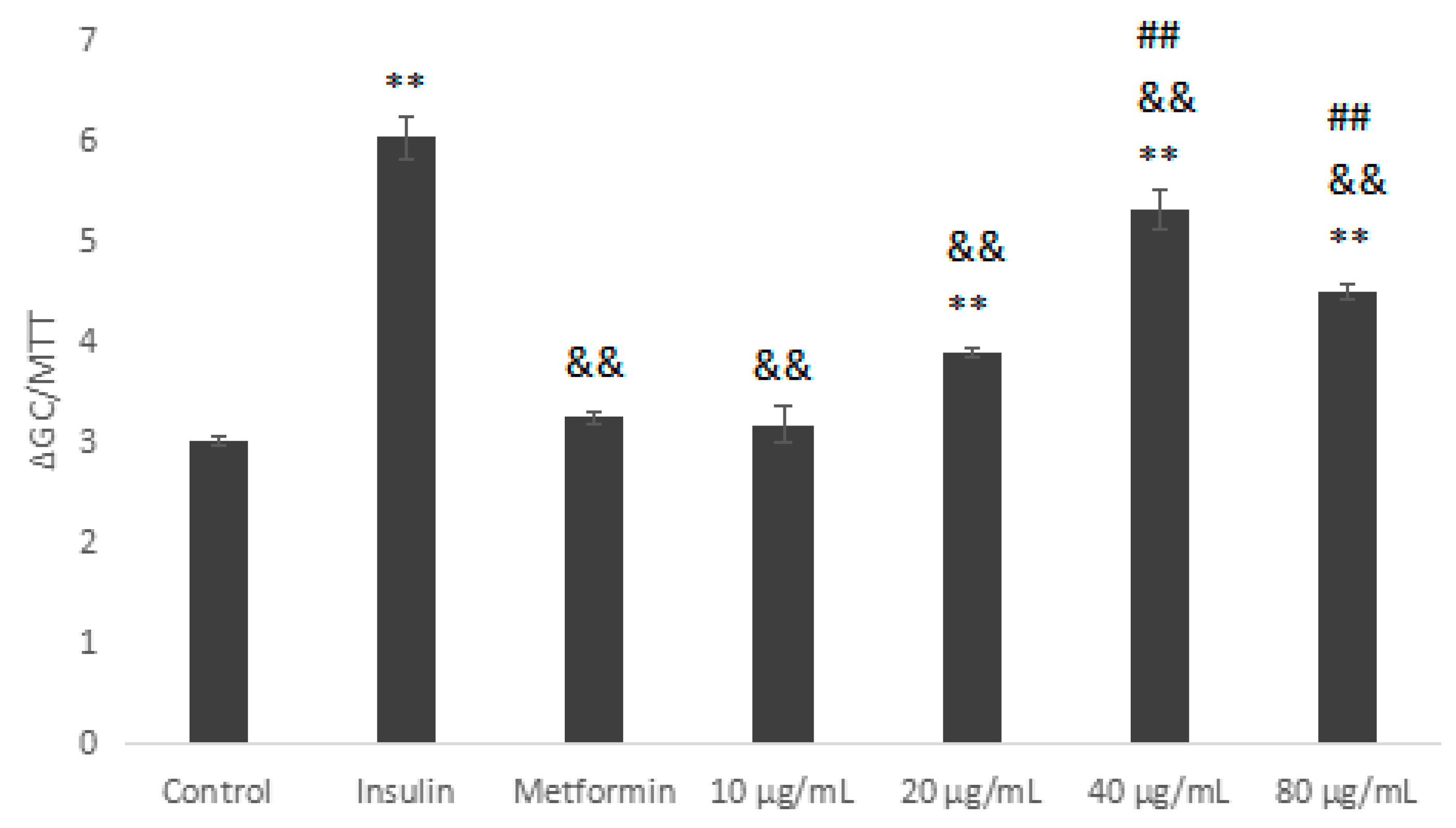
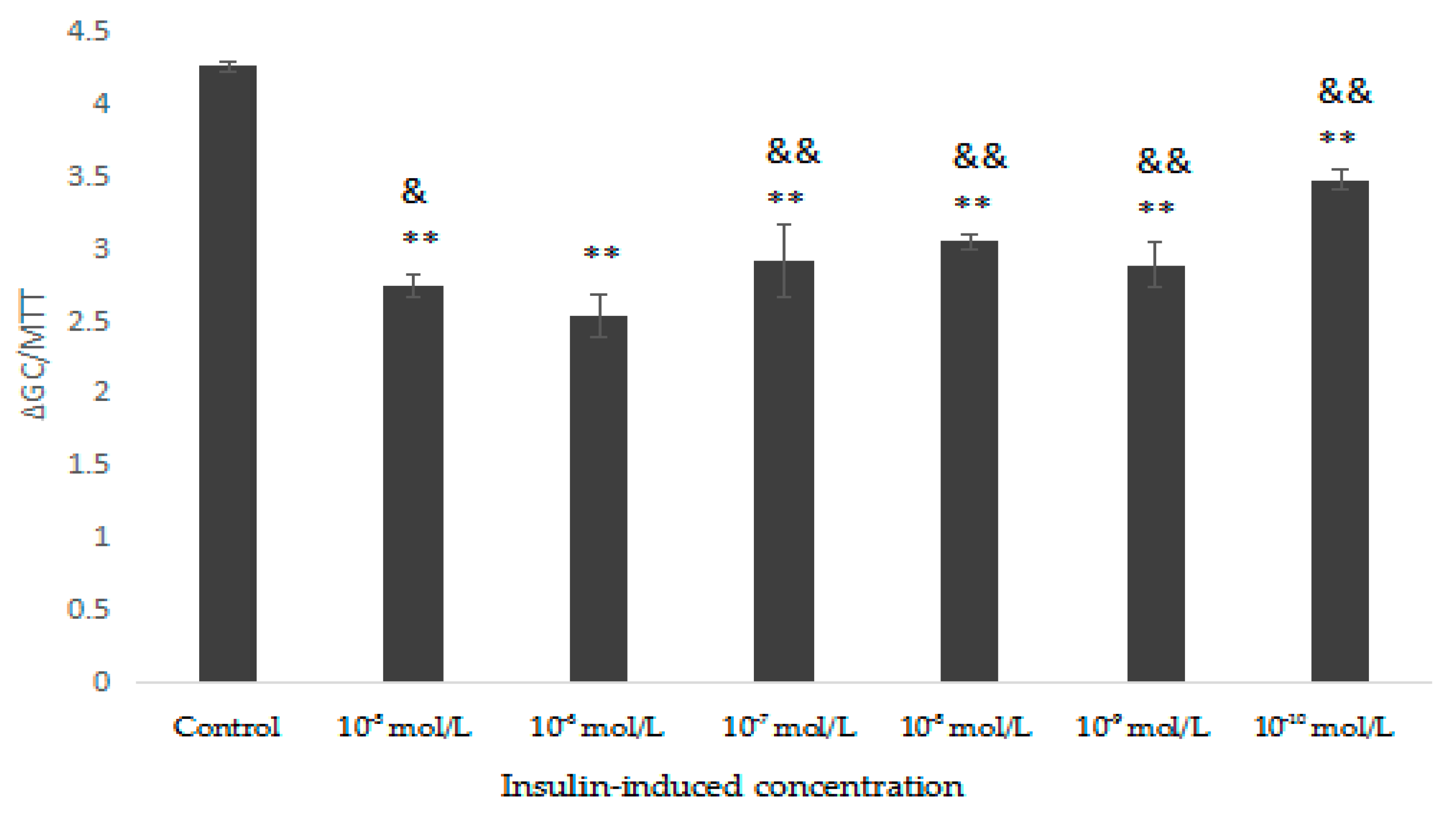
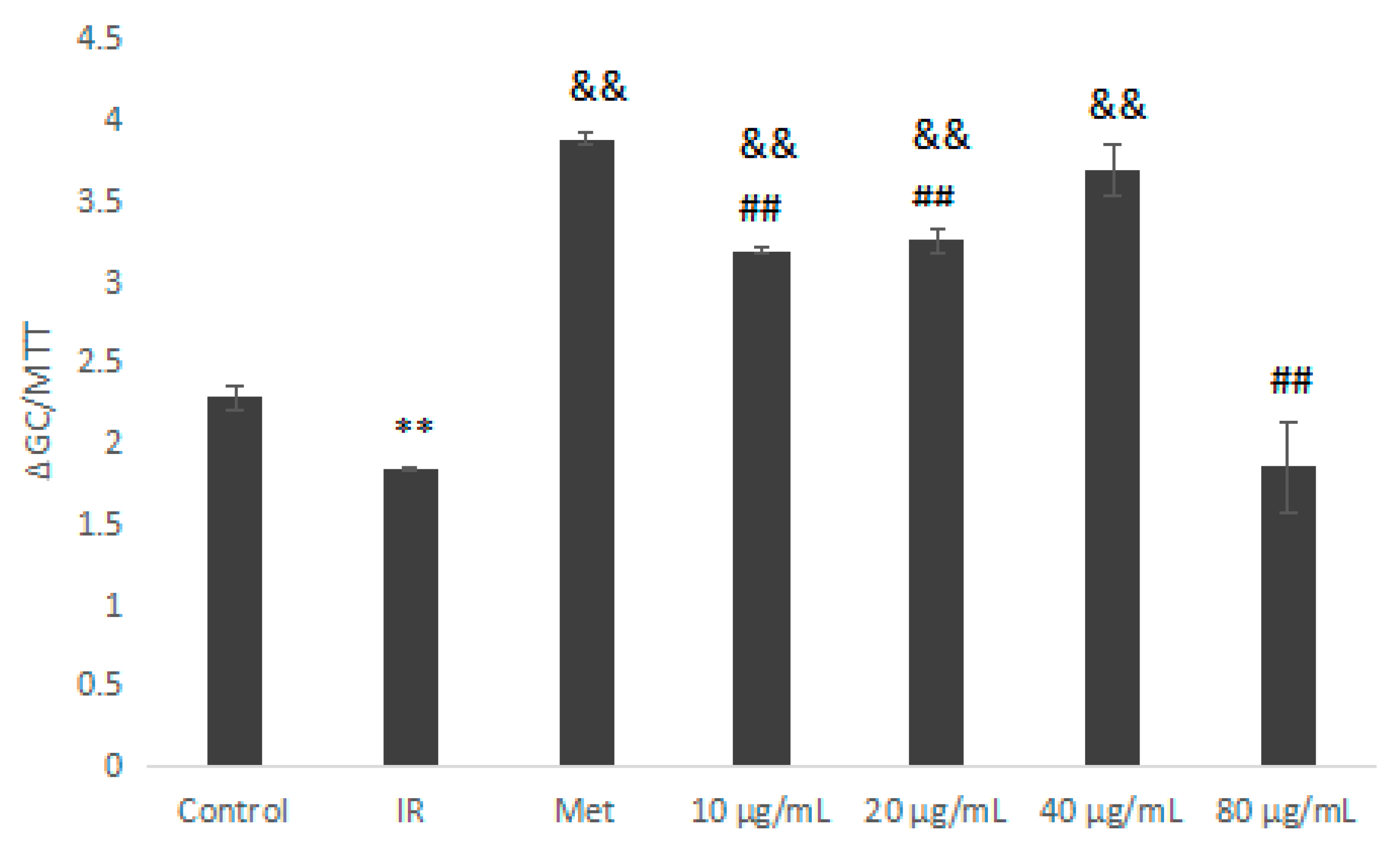
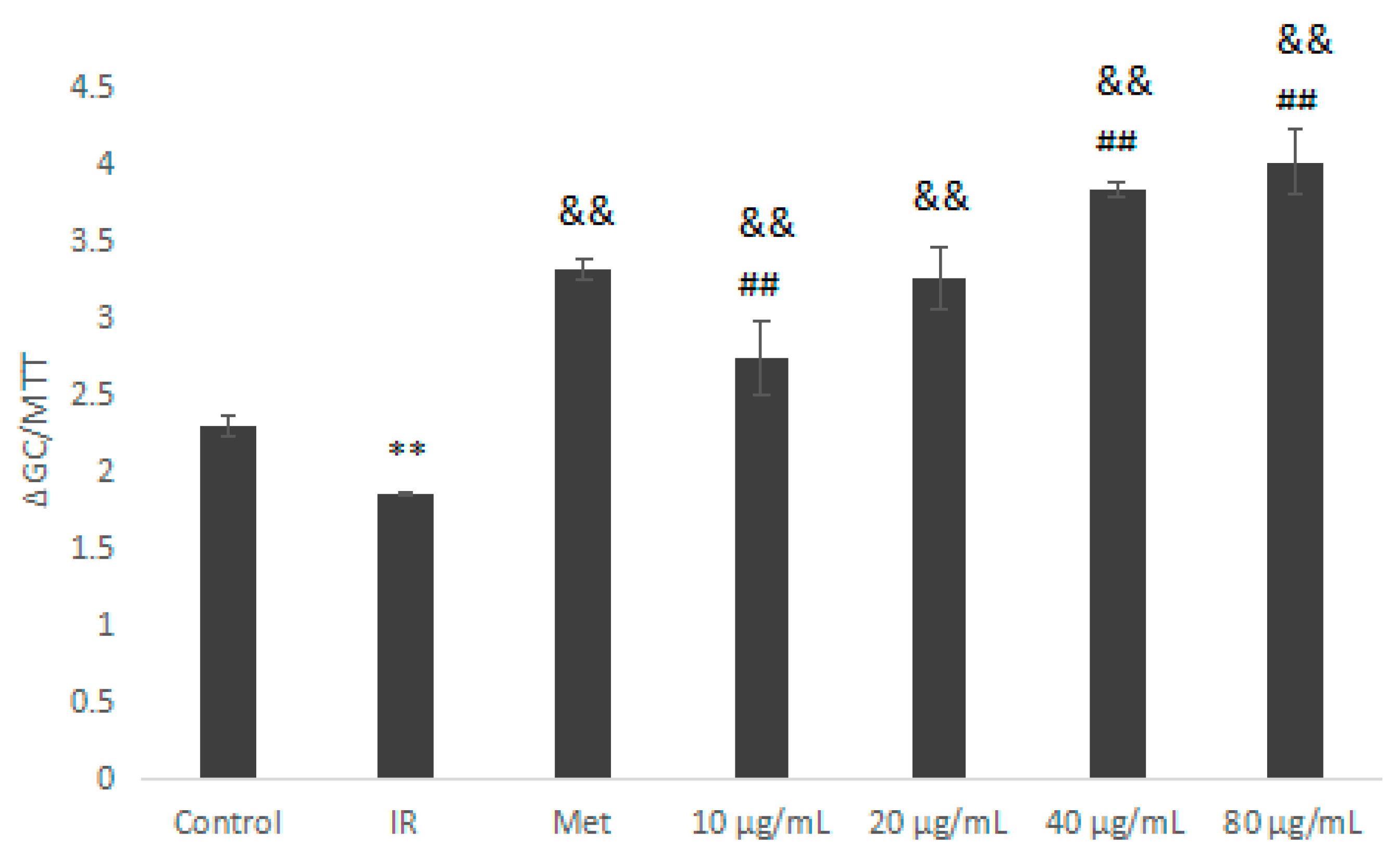
2.5.3. Glucose Consumption Assay on Insulin Resistant HepG2 Cells
3. Materials and Methods
3.1. Chemicals and Regents
3.2. Liquid Culture
3.3. Extraction, Isolation and Purification of Polysaccharides
3.4. Analysis of Chemical Composition
3.5. Determination of Homogeneity and Molecular Weight
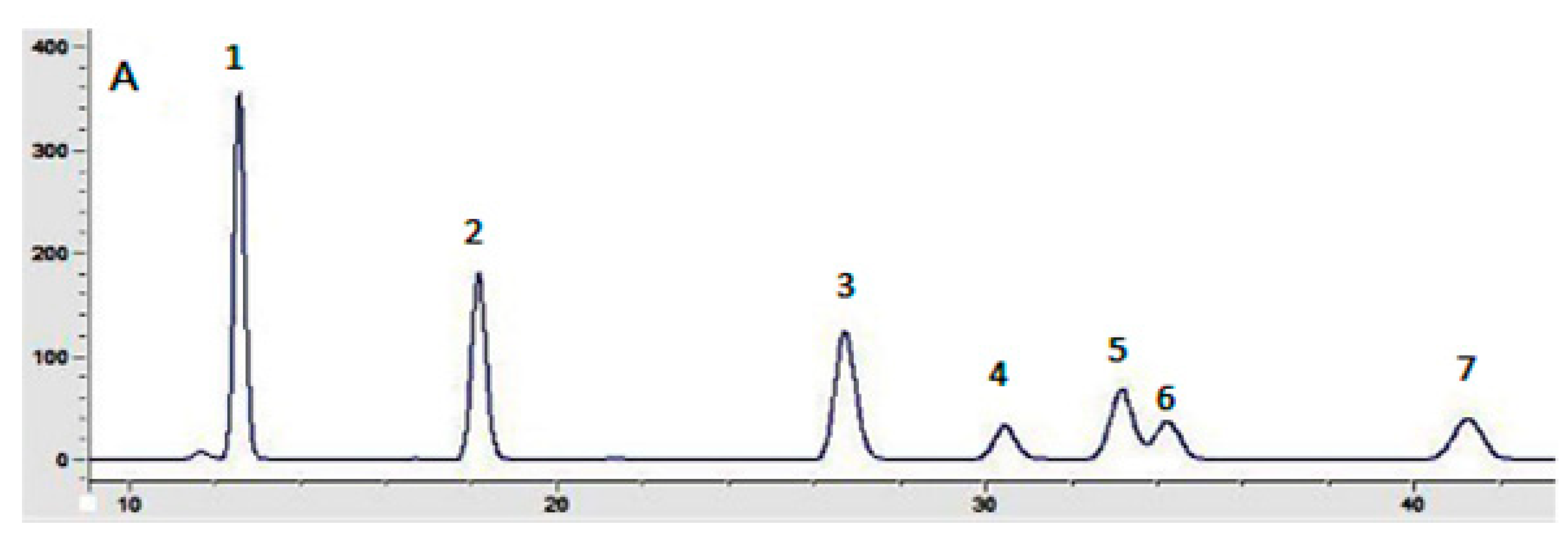
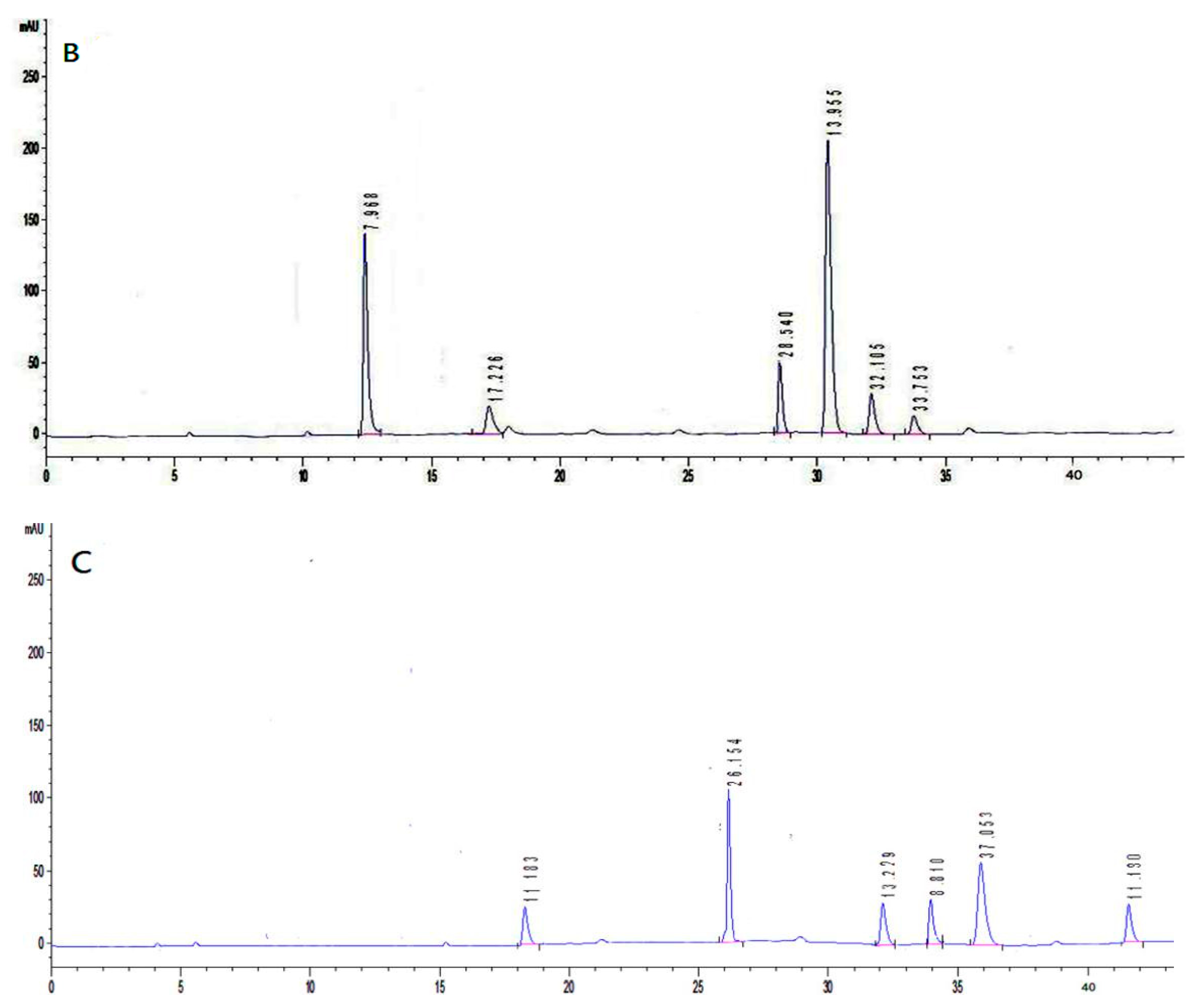
3.6. Monosaccharide Composition Analysis
3.7. FT-IR Analysis
3.8. 1H-NMR Analysis
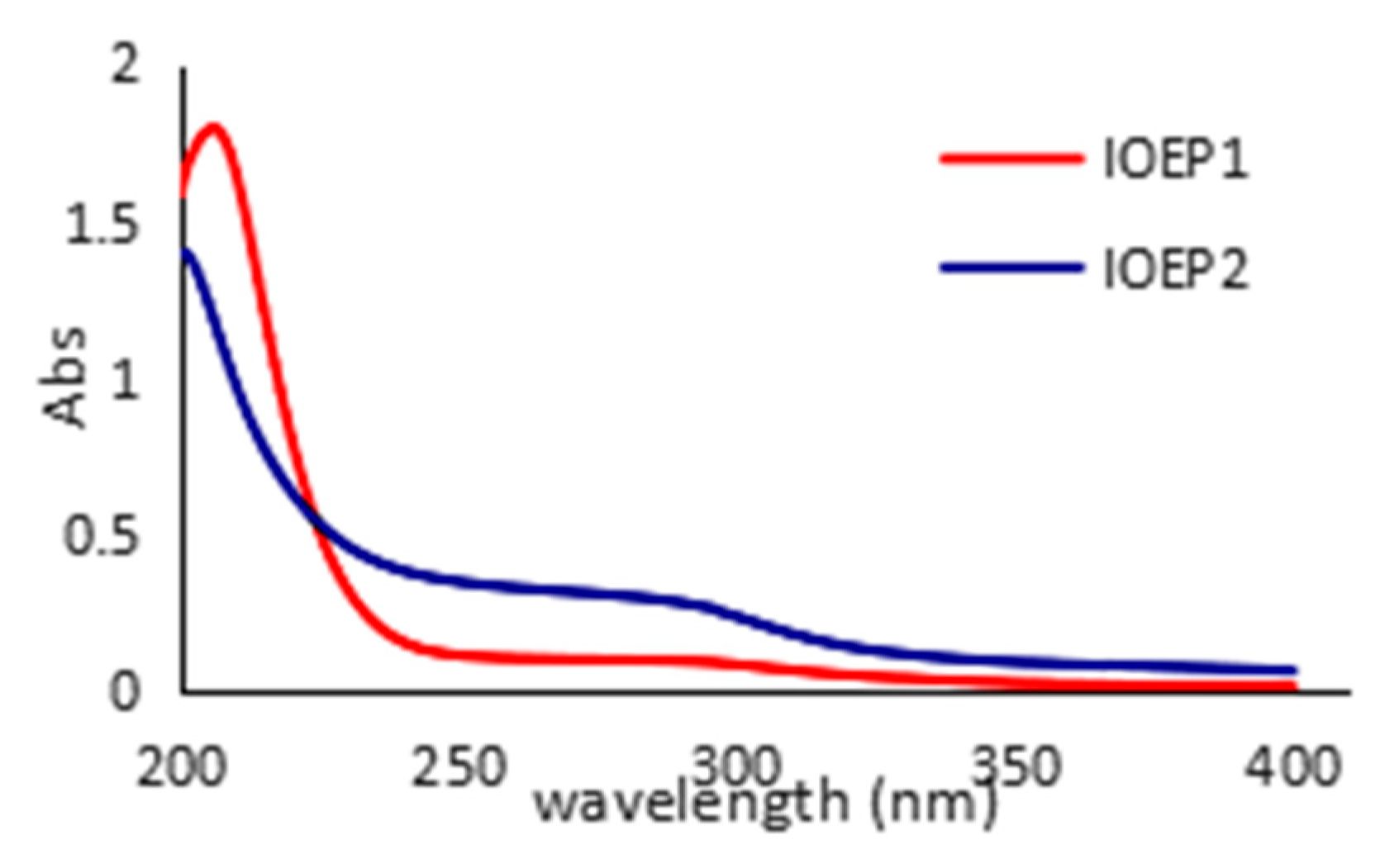
3.9. Ultraviolet Spectrum
3.10. Hypoglycaemic Activity Assay In Vitro
3.10.1. Cytotoxicity Assay on HepG2 Cells
3.10.2. Glucose Consumption Assay on HepG2 Cells
3.10.3. Glucose Consumption Experiment on Insulin-Resistance (IR) HepG2 Cells
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wang, Y.T.; Lin, D.B.; Wang, X.L.; Zhu, W.; Ye, J.L.; Li, G.H. The impact of a noval peach gum-derived polysaccharide on postprandial blood glucose control in streptozotocin-induced diabetic mice. Int. J. Boil. Macromol. 2017, 98, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Gan, M.J.; Albanese-O’ Neill, A.; Haller, M.J. Type 1 Diabetes: Current Concepts in Epidemiology, Pathophysiology, Clinical Care, and Research. Curr. Probl. Pediatr. Adolesc. Health Care 2012, 42, 269–291. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, T.; Chait, A.; Plutzky, J. Cardiovascular disease risk in type 2 diabetes mellitus: Insights from mechanistic studies. Lancet 2008, 371, 1800–1809. [Google Scholar] [CrossRef]
- KoKil, G.R.; Veedu, R.N.; Ramm, G.A.; Prins, J.B.; Parekh, H.S. Type 2 Diabetes Mellitus: Limitations of Conventional Therapies and Intervention with Nucleic Acid-Based Therapeutics. Chem. Rev. 2015, 115, 4719–4743. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.E.; Ao, Z.H. Antihyperglycemic and antilipidperoxidative effects of dry matter of culture broth of Inonotus obliquus in submerged culture on normal and alloxan-diabetes mice. J. Ethnopharmacol. 2008, 118, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, D. Lipid-lowering drugs in the management of hyperlipidaemia. Pharmacology and Therapeutics. Pharmacol. Ther. 1988, 79, 205–230. [Google Scholar] [CrossRef]
- May, L.D.; Lefkowitch, J.H.; Kram, M.T.; Rubin, D.E. Mixed hepatocelluar-cholestatic liver injury after pioglitazone therapy. Ann. Intern. Med. 2002, 136, 449–452. [Google Scholar] [CrossRef]
- Liu, L.H.; Huang, J.H. Study on pumpkin polysaccharide reducing the blood glucose on the diabetic model mouses by Alloxan. Chin. J. Trauma Disabil. Med. 2007, 15, 27–29. [Google Scholar]
- Xu, X.Q.; Zhang, X.; Chen, C. Stimulated production of triterpenoids of Inonotus obliquus using methyl jasmonate and fatty acids. Ind. Crop. Prod. 2016, 85, 49–57. [Google Scholar] [CrossRef]
- Taji, S.; Yamada, T.; Wada, S.; Tokuda, H.; Sakuma, K.; Tanaka, R. Lanostane-type triterpenoids from the sclerotia of Inonotus obliquus possessing anti-tumor promoting activity. Eur. J. Med. Chem. 2008, 43, 2373–2379. [Google Scholar] [CrossRef]
- Zheng, W.F.; Zhang, M.M.; Zhao, Y.X.; Wang, Y.; Miao, K.J.; Wei, Z.W. Accumulation of antioxidant phenolic constituents in submerged cultures of Inonotus obliquus. Bioresour. Technol. 2009, 100, 1327–1335. [Google Scholar] [CrossRef] [PubMed]
- Handa, N.; Yamada, T.; Tanaka, R. An unusual lanostane-type triterpenoid, spiroinonotsuoxodiol, and other triterpenoids from Inonotus obliquus. Phytochemistry 2010, 71, 1774–1779. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.N.; Hui, J.; Kou, W.; Xin, R.; Jia, F.; Wang, N.; Hu, F.Q.; Zhang, H.L.; Liu, H.S. Identification of Inonotus obliquus and analysis of antioxidation and antitumor activities of polysaccharides. Curr. Microbiol. 2008, 57, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.M.; Chen, H.X.; Fu, L.L.; Zhang, X. Phytochemical characteristics and hypoglycaemic activity of fraction from mushroom Inonotus obliquus. J. Sci. Food Agric. 2010, 90, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Miao, K.; Liu, Y.; Zhao, Y.; Zhang, M.; Pan, S. Chemical diversity of biologically active metabolites in the sclerotia of Inonotus obliquus and submerged culture strategies for up-regulating their production. Appl. Microbiol. Biotechnol. 2010, 87, 1237–1254. [Google Scholar] [CrossRef]
- Chen, H.; Yan, M.C.; Zhu, J.W.; Xu, X.Q. Enhancement of exo-polysaccharide production and antioxidant activity in submerged cultures of Inonotus obliquus by lignocellulose decomposition. J. Ind. Microbiol. Biotechnol. 2011, 38, 291–298. [Google Scholar] [CrossRef]
- Mishra, S.K.; Kang, J.H.; Kim, D.K.; Oh, S.H.; Kim, M.K. Orally administered aqueous extract of Inonotus obliquus ameliorates acute inflammation in dextran sulfate sodium (DSS)-induced colitis in mice. J. Ethnopharmacol. 2011, 38, 291–298. [Google Scholar] [CrossRef]
- Xu, X.Q.; Wu, Y.D.; Chen, H. Comparative antioxidative characteristics of polysaccharide-enriched extracts from natural sclerotia and cultured mycelia in submerged fermentation of Inonotus obliquus. Food Chem. 2011, 127, 74–79. [Google Scholar] [CrossRef]
- Liu, C.; Zhao, C.; Pan, H.H.; Kang, J.; Yu, X.T.; Wang, H.Q.; Li, B.M.; Xie, Y.Z.; Chen, R.Y. Chemical constituents from Inonotus obliquus and their biological activities. J. Nat. Prod. 2014, 77, 35–41. [Google Scholar] [CrossRef]
- Tanaka, R.; Toyoshima, M.; Yamada, T. New lanostane-type triterpenoids inonotsutriols D, and E, from Inonotus obliquus. Phytochem. Lett. 2011, 4, 328–332. [Google Scholar] [CrossRef]
- Kim, Y.J.; Park, J.; Min, B.S.; Shim, S.H. Chemical constituents from the sclerotia of Inonotus Obliquus. J. Korean Soc. Appl. Boil. Chem. 2011, 54, 287–294. [Google Scholar] [CrossRef]
- Diao, B.Z.; Jin, W.R.; Yu, X.J. Protective effect of polysaccharides from Inonotus obliquus on Streptozotocin-induced diabetic symptoms and their potential mechanisms in rats. Evid.-Based Complement. Altern. Med. 2014, 2014, 841496. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Zhao, T.; Mao, G.H.; Zhang, M.; Zheng, D.H.; Feng, W.W.; Wang, W.W.; Wu, X.Y.; Yang, L.Q. Isolation, purification and characterization of selenium-containing polysaccharides and proteins in selenium-enriched Radix puerarse. J. Sci. Food Agric. 2014, 94, 349–358. [Google Scholar] [CrossRef]
- Hu, T.; Huang, Q.L.; Wong, K.H.; Yang, H. Structure, molecular conformation, and immunomodulatory activity of four polysaccharide fractions from Lignosus rhinocerotis sclerotia. Int. J. Boil. Macromol. 2017, 94, 423–430. [Google Scholar] [CrossRef]
- Chokboribal, J.; Tachaboonyakiat, W.; Sangvanich, P.; Ruangpornvistuti, V.; Jettanacheawchankit, S.; Thunyakitpisal, P. Deacetylation affects the physical properties and bioactivity of acemannan, an extracted polysaccharide from Aloe vera. Carbohydr. Polym. 2015, 133, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Chawananorasest, K.; Saengtongdee, P.; Kaemchantuek, P. Extraction and characterization of Tamarind seed polysaccharide (TSP) from three different sources. Molecules 2016, 21, 775. [Google Scholar] [CrossRef]
- Wu, Y.; Lei, Y.; Li, E.T.; Li, Y.Y.; Lu, Y.; Wang, P.J.; Zhou, H.L.; Liu, J.G.; Hu, Y.L.; Wang, D.Y. Optimization of Glycyrrhiza polysaccharide liposome by response surface methodology and its immune activities. Int. J. Boil. Macromol. 2017, 216, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Zhang, Y.F.; Liu, Z.K.; Zhao, M.Q.; Liu, P.F. Purification, characterization, and antioxidant activity of polysaccharide isolated from Cortex Periplocae. Molecules 2017, 22, 1866. [Google Scholar] [CrossRef]
- Wang, X.T.; Zhu, Z.Y.; Zhao, L.; Sun, H.Q.; Meng, M.; Zhang, J.Y.; Zhang, Y.M. Structural characterization and inhibition on α-d-glucosidase activity of non-starch polysaccharides from Fagopyrum tartaricum. Carbohydr. Polym. 2016, 153, 679–685. [Google Scholar] [CrossRef]
- Sirajunnisa, A.R.; Vijayagopal, V.; Sivaprakash, B.; Viruthagiri, T.; Surendhiran, D. Optimization, kinetics and antioxidant activity of extropolysaccharides produced from rhizosphere isolate, Pseudomonas fluorescens CrN6. Carbohydr. Polym. 2016, 135, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xue, J.; Tong, S.S.; Dong, W.X.; Wu, P.P. Structure Characterization and hypoglycaemic activities of two polysaccharides from Inonotus Obliquue. Molecules 2018, 23, 1948. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.C.; Dong, W.W.; Fang, L.Y.; Liao, B.L. Purification, preliminary characterization and hepatoprotective effects of polysaccharides from Dandelion Root. Molecules 2017, 22, 1409. [Google Scholar] [CrossRef] [PubMed]
- Cong, M.; Shang, Q.; Dong, W.; Liao, F.; Ding, K. Structure and activities of a novel heteroxylan from Cassia obtusifolia seeds and its sulfated derivative. Carbohydr. Res. 2014, 393, 43–50. [Google Scholar] [CrossRef]
- Zha, S.H.; Zhao, Q.S.; Chen, J.J.; Wang, LW.; Zhang, G.F.; Zhang, H.; Zhao, B. Extraction, purification and antioxidant activities of the polysaccharides from maca (Lepidium meyenii). Carbohydr. Polym. 2014, 111, 584–587. [Google Scholar] [CrossRef]
- Simas, F.F.; Corin, P.A.J.; Wagner, R.; Sassaki, G.L.; Bonkerner, A.; Lacomini, M. Comparison of structure of gum exudate polysaccharides from the trunk and fruit of the peach tree (Prunus persica). Carbohydr. Polym. 2008, 71, 218–228. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, J.; Zhang, L. Structure and chain conformation of β-glucan isolated from Auricularia auricula-judae. Biopolymers 2008, 89, 614–622. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, W.; Yu, J.P.; Zou, S.; Wang, J.J.; Yao, W.B.; Gao, X.D. Characterization and hypoglycemic effect of a polysaccharide extracted from the fruit of Lycium Barbarum L. Carbohydr. Polym. 2013, 98, 8–16. [Google Scholar] [CrossRef]
- Jia, L. Ameliorative Effects and Mechanism of Lyceum Barbarum Polysaccharide on Insulin Resistance of Hep G2 Cell. Master’s Thesis, University of South China, Hengyang, China, 2016. [Google Scholar]
Sample Availability: Samples of the compounds IOEP1 and IOEP2 are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IOEP1 (%) | IOEP2 (%) |
|---|---|---|
| Carbohydrate (%) | 87.04 ± 1.33 a | 78.73 ± 1.57 b |
| Protein (%) | 0.831 ± 0.73 a | 1.521 ± 0.34 b |
| Concentration (μg/mL) | Cell survival rate (%) | ||
|---|---|---|---|
| IOEP1 | IOEP2 | ||
| Control Metformin Insulin | - | 100 | 100 |
| - | 100.1 ± 2.31 | 100.1 ± 2.31 | |
| - | 98.40 ± 4.69 | 98.40 ± 4.69 | |
| 10 | 97.74 ± 0.10 | 97.69 ± 2.05 | |
| 20 | 94.43 ± 1.54 | 95.88 ± 4.48 | |
| 40 | 93.77 ± 4.26 | 95.29 ± 1.66 | |
| 80 | 90.78 ± 2.01 | 93.45 ± 1.23 | |
| 160 | 87.90 ± 2.01 | 92.11 ± 1.68 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, J.; Tong, S.; Wang, Z.; Liu, P. Chemical Characterization and Hypoglycaemic Activities In Vitro of Two Polysaccharides from Inonotus obliquus by Submerged Culture. Molecules 2018, 23, 3261. https://doi.org/10.3390/molecules23123261
Xue J, Tong S, Wang Z, Liu P. Chemical Characterization and Hypoglycaemic Activities In Vitro of Two Polysaccharides from Inonotus obliquus by Submerged Culture. Molecules. 2018; 23(12):3261. https://doi.org/10.3390/molecules23123261
Chicago/Turabian StyleXue, Jiao, Shisheng Tong, Zhaorun Wang, and Ping Liu. 2018. "Chemical Characterization and Hypoglycaemic Activities In Vitro of Two Polysaccharides from Inonotus obliquus by Submerged Culture" Molecules 23, no. 12: 3261. https://doi.org/10.3390/molecules23123261
APA StyleXue, J., Tong, S., Wang, Z., & Liu, P. (2018). Chemical Characterization and Hypoglycaemic Activities In Vitro of Two Polysaccharides from Inonotus obliquus by Submerged Culture. Molecules, 23(12), 3261. https://doi.org/10.3390/molecules23123261