1. Introduction
Lignans are a large group of phenylpropanoid dimers with a different degree of oxidation in the side-chain and a different substitution in the aromatic group [
1,
2]. They are classified in groups according to their oxygenation and cyclization patterns. The most prominent member of this group of natural products is podophyllotoxin (PTOX,
1). Its antitumour activity prompted several studies, and resulted in the introduction of successful clinical drugs. This aryltetralin lignan is a lead compound for the semi-synthetic derivatives etoposide (
27), teniposide (
28), and etopophos (
29) (
Figure 1), which have an important role in cancer therapy [
3,
4]. In addition, analogues of podophyllotoxin (
1) were evaluated for the treatment of rheumatoid arthritis, psoriasis, and malaria with good results [
3,
5]. Furthermore, there are reviews published referring to the semisynthesis of PTOX derivatives, applications, mode of action and structure-activity relationships [
6,
7,
8].
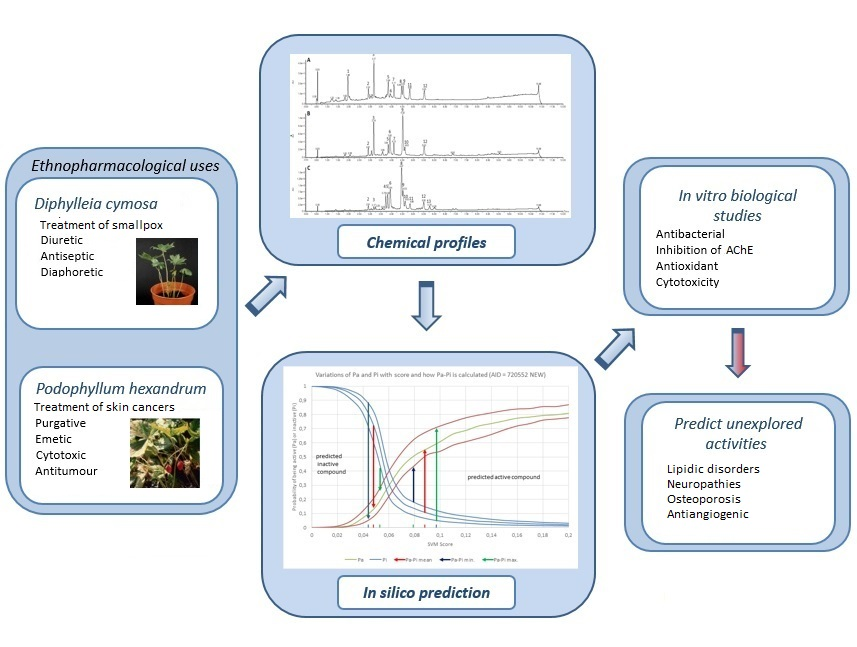
Arylnaphatalene lignans such as diphyllin (
24) and its glycosides
25 and
26 (
Figure 1) were isolated from some traditional medicinal plants and have been reported to possess a wide range of pharmacological activities, including antitumour, anti-leshmania, antifungal, antiviral and antibacterial [
9,
10].
Currently, there are few plant sources of podophyllotoxin (
1) and its related lignans occur in a particular taxonomic group, but in low amounts. Podophyllotoxin (
1) is still obtained from wild
Podophyllum populations, and this is a major constraint in supplying the lignan to the pharmaceutical industry that is under pressure to meet demand. To overcome this situation, several studies focussing on its production by biotechnological strategies and synthetic approaches have been reported [
11,
12].
Podophyllotoxin (
1) is found in the rhizomes, roots and leaves of both
Podophyllum hexandrum Royle and
Diphylleia cymosa Michaux (Berberidaceae), while the occurrence of diphyllin (
24) is reported in the latter species but not in
Podophyllums [
13].
P. hexandrum and
D. cymosa are herbaceous perennials found growing in moist shady conditions [
14], and are known for their medicinal use in American and Asian cultures. Both genera are taxonomically closely related, and some common features are their habitat, morphology, karyotype and chemical profile, whilst the differences are related to floral biology [
13,
15,
16]. A study based on four molecular markers and morphology confirms the close relationship between
Diphylleia and
Podophyllum [
17].
P. hexandrum is sometimes treated as a monotypic genus
Sinopodophyllum [
18].
P. hexandrum is commonly named as the Himalayan Mayapple or Indian Mayapple and Indian
Podophyllum. There are ethnobotanical records based on its healing properties in Asian culture for the treatment of skin cancers as well as due to its purgative, emetic, cytotoxicity, antitumour and antileukaemic properties [
11,
19]. Overall, the parts used for medicinal purposes are mainly the rhizomes, roots and fruits.
D. cymosa has been called the Southern Mayapple and Umbrella Leaf [
20]. There is an account that describes the American Cherokee Indians using an infusion of the plant as a diuretic, antiseptic, diaphoretic and for the treatment of smallpox [
21]. According to an earlier clinical study, the resin demonstrated none of the biological properties associated with
Podophyllum [
22]. Considerable interest has been centered on
P. hexandrum due to the PTOX content and its related lignans. With regard to
D. cymosa, the species has been investigated less than the
Podophyllums and only a few studies have been reported [
13,
23,
24].
This paper reports on the evaluation of ethanolic extracts from the leaves and roots of
D. cymosa, and from the rhizomes and roots of
P. hexandrum for the antibacterial activity, inhibition of AChE, as well as for the antioxidant activity and cytotoxicity combined with an in silico target fishing approach. The latter was used to predict new activities for known lignans from both species. The lignans profiling was based on the chromatographic analyses (UPLC-DAD-ESI-MS/MS) which were included with the aim of identifying the major phenolic components in the extracts (
Figure 2).
2. Results
A growing amount of work has been applied to investigating
P. hexandrum due to its content of PTOX (
1) and related lignans. However, it is surprising to note that studies into
D. cymosa are extremely limited when compared with the
Podophyllum species, even though this species is endemic in the Southern Appalachian Mountains of the Eastern North America [
22,
26].
There has been a decrease in the wild populations of
P. hexandrum in India due to the over collection of rhizomes and roots of this species [
27]. In addition, the species shows a short season of availability, and thus plants are limited in the field. A number of studies have been undertaken to achieve mass propagation [
28] as well as to establish plant derived-cultures for the production of podophyllotoxin [
29,
30], whereas their low yields were far from meeting commercial needs. Enhancement of the lignan was attempted by other systems, including transgenic cultures, addition of a precursor feeding to the culture medium, the use of an elicitor such as methyl jasmonate, and the production by endophytes [
11]. With regard to the latter, there are reports on the production of PTOX (
1) by the endophytic fungi
Fusarium solani [
31] and
Trametes hirsuta [
32] isolated from
P. hexandrum. These species of endophytes could be a promising source for large-scale production of PTOX, whereas the yields must be improved.
2.1. Chromatographic Profiling by UPLC-DAD-ESI-MS/MS
Several lignans have been already reported for
D. cyomsa and
P. hexandrum. From the leaves and roots of
D. cymosa, Broomhead and Dewick [
13] isolated the lignans PTOX (
1), 4′-demethyl-podophyllotoxin (
6), 4′-demethyldesoxypodophyllotoxin (
9), diphyllin (
24), diphyllin glucoside (
25), diphyllin diglucoside (
26) and 4′-demethyldesoxypodophyllotoxin 4-
O-glucoside (
10) (
Figure 1), this being the only study reporting the isolation and characterization of compounds from
D. cymosa.
A phytochemical study of
P. hexandrum led to the isolation of the lignans PTOX (
1), 4′-demethylpodophyllotoxin (
6), 4′-demethyldesoxypodophyllotoxin (
9), PTOX glucoside (
3), desoxy-podophyllotoxin (deoxypodophyllotoxin,
5), 4′-demethylpodophyllotoxin glucoside (
7), 4′-demethylisopicropodophyllone (
15), podophyllotoxone (
11), 4′-demethylpodophyllotoxone (
12), picropodophyllotoxin (
13), isopicropodophyllone (
14), 4′-demethyldeoxypodophyllotoxin (4′-demethyldesoxypodophyllotoxin,
9), α-peltatin (
18) and β-peltatin (
19) (
Figure 1) [
13].
In this study, the chemical characterization of
D. cymosa and
P. hexandrum was performed using UPLC-DAD-ESI-MS. The obtained chromatographic profiles indicated the presence of compounds of different polarities in the EtOH extracts of
D. cymosa and
P. hexandrum (
Figure 3). The crude extracts as well as lignans previously isolated from
P. hexandrum such as PTOX (
1), deoxypodophyllotoxin (
5), 4′-demethylpodophyllotoxin (
6), podophyllotoxone (
11), α-peltatin (
18) and β-peltatin (
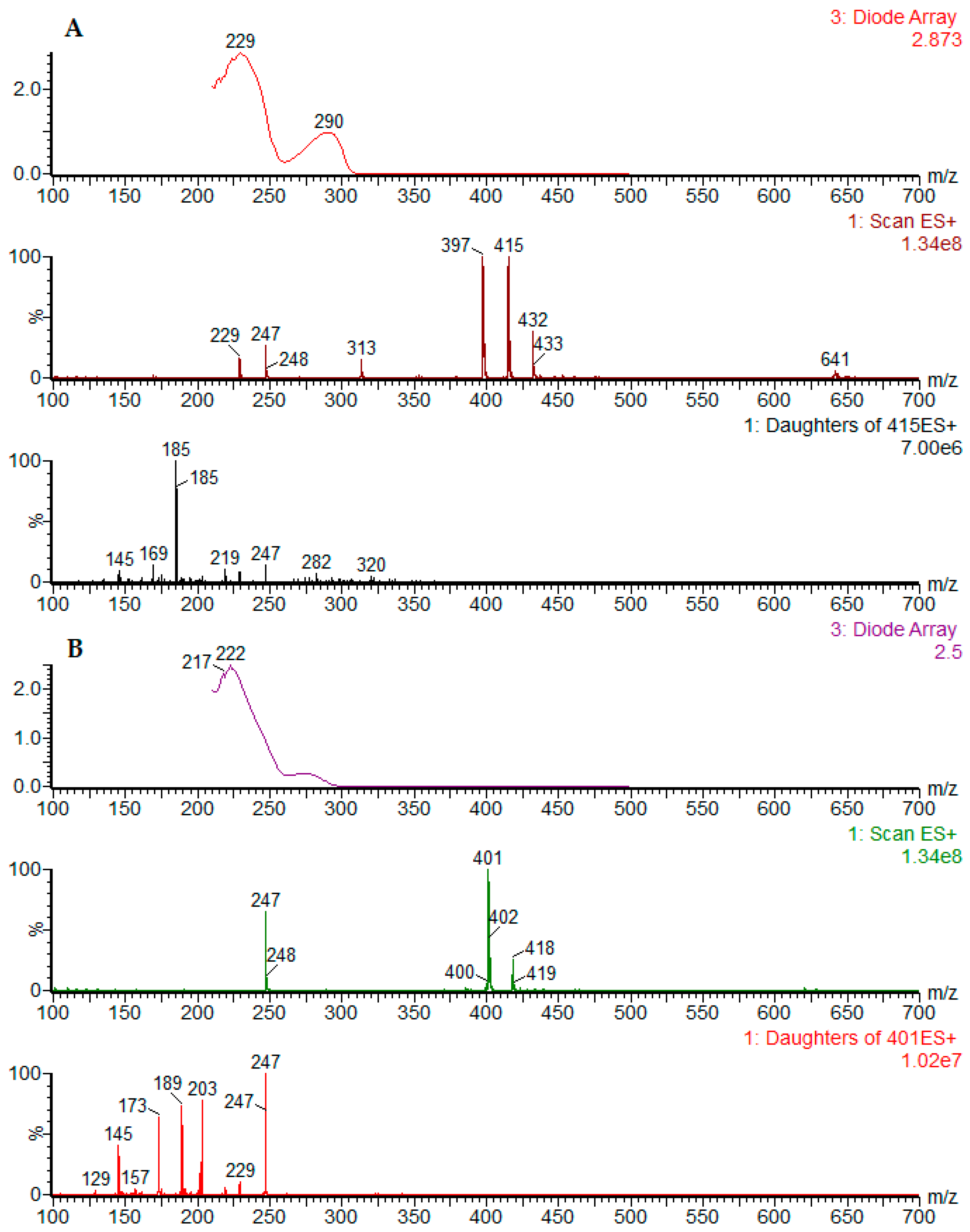
19) were evaluated in the same chromatographic conditions. The UV and ESI
+-MS spectra for the reference compounds podophyllotoxin (
1) and α-peltatin (
18) are presented in
Figure 4 (
Figure 4A,B). The UV and MS spectra for all reference compounds and identified chromatographic peaks are available in the
Supplementary Material.
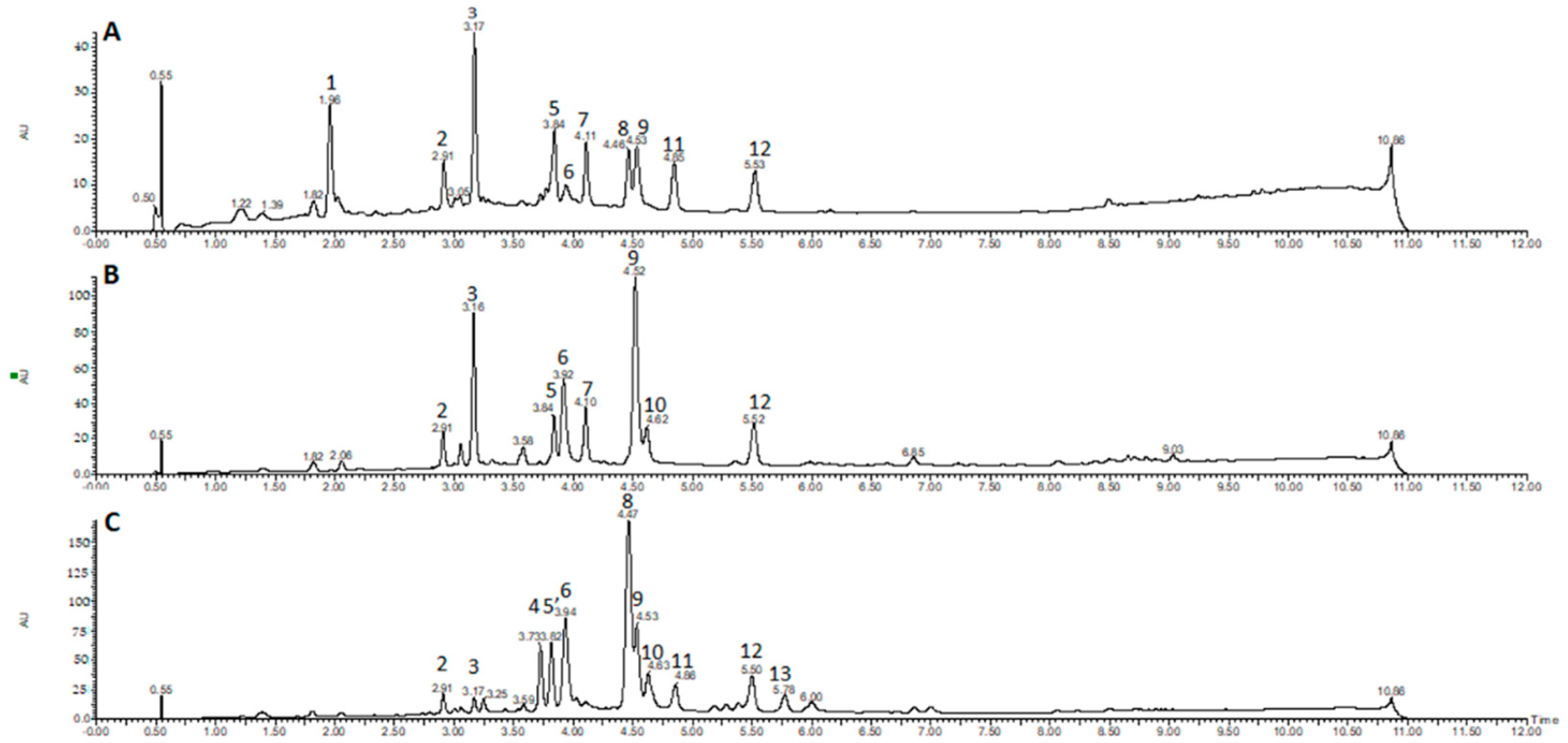
The chromatographic profile obtained by UPLC-DAD for the EtOH extracts of
D. cymosa leaves (
Figure 3A),
D. cymosa roots (
Figure 3B) and
P. hexandrum rhizomes and roots (
Figure 3C) showed peaks with UV absorption spectra with λ
max around 260–290 nm which is compatible with the chemical structure of lignans due to conjugation of the aromatic rings. It was possible to identify peaks with UV absorption pattern characteristic of aryltetralin lignans related to podophyllotoxin (peaks 4, 8, 11 and 13, λ
max at 290 nm) and peltatins (peaks 6 and 10, λ
max at 275 nm), and arylnaphtalene lignans, such as diphyllin (peaks 5, 7 and 12; λ
max at ca. 260 nm). Peaks with UV spectra characteristic of other phenolic compounds were also identified, such as phenolic acids (peak 1, λ
max at 246, 295 and 326 nm) and flavonoids (peaks 2, 3 and 9, λ
max at 265 and ca. 350 nm), which are compounds with more conjugated chromophores.
UPLC-ESI-MS/MS analyses were carried out in order to identify the major constituents of the extracts (
Table 1). Chromatographic peak 1 (RT = 1.96 min) showed
m/
z at 353 and 355 in the negative and positive ionization modes, respectively. The UV absorption profile of compound
1 was indicative of phenolic acids (λ
max ~ 295 and 326 nm). The fragmentation of the parent ion at
m/
z 353 in a MS2 experiment in the negative mode afforded daughter ions at
m/
z 191 [M − H − caffeoyl]
−, 179 [M − H − quinic]
−, and 173 [quinic acid − H − H
2O]
− suggesting the identity of compound
1 as one of the regioisomers of caffeoylquinic acid. The fragmentation pattern of caffeoylquinic acids have been extensively described [
33].
The chromatographic peaks 2, 3 and 9 had a UV absorption profile indicative of flavonols, with λ
max ~ 255–265 and 350 nm. MS spectra associated to peak 2 (RT = 2.91 min) showed a peak of [M − H]
− at
m/
z 463 and [M + H]
+ at
m/
z 465. The MS2 fragmentation of the parent ion at at
m/
z 465 in the positive mode afforded a daughter ion at
m/
z 303 [M + H − hexose]
+, indicating that peak 2 could correspond to a hexoside of the flavonol quercetin. The observed fragmentation pattern of the ion at
m/
z 303 generated a peak at
m/
z 165, which is indicative of the presence of a hydroxyl group at C
3 of flavonols. A similar UV absorption profile was observed for peak 3 (RT = 3.17 min), which showed a MS spectra with a peak of [M − H]
− at
m/
z 447 and [M + H]
+ at
m/
z 449. The MS2 fragmentation of the parent ions at
m/
z 447 and
m/
z 449 afforded daughter ions at
m/
z 285 [M − H − hexose] and 287 [M + H − hexose] in the negative and positive ionization modes, respectively. The 16 a.m.u. difference observed between peaks 2 and 3, along with the similarity in the UV spectra and in the fragmentation pattern obtained in MS2 experiments, suggest that peak 2 is a hexoside of the flavonol kaempferol. This is the major peak observed in the chromatogram of
D. cymosa leaves (
Figure 3A). The compound corresponding to peak 9 (RT = 4.53 min) showed a UV and MS profiles similar to those observed for peak 3. The MS spectra registered for this compound presented a peak at
m/
z 285 corresponding to the deprotonated molecule [M − H]
−. The fragmentation pattern of the parent ion at
m/
z 285 was similar to that observed for peak 3, suggesting that peak 9 corresponds to the aglicone kaempferol. This flavonol is the major constituent of the EtOH extract of
D. cymosa roots.
Analysis of the UV and MS spectra associated with peaks 4 (RT = 3.73 min) and 8 (RT = 4.46 min) showed λmax at 290 nm, indicative of aryltetralin lignans. MS spectra associated with peaks 4 and 8 showed signals of [M + H]+ at m/z 577 and 415, respectively. The same pattern was observed in the negative ionization mode, with signals of [M − H]− at m/z 575 and 459 [M − H + formiate]− for peaks 4 and 8, respectively. The difference of 162 Da between the two compounds indicates the presence of a hexose residue. The fragmentation of the parent ion at m/z 415 in the positive ionization mode generated the daughter ion at m/z 247 which is compatible with the neutral loss of a trimethoxybenzyl group (C9H12O3). These results, along with the analysis of the isolated lignan podophyllotoxin in the same conditions, allowed the identification of compound 8 as podophyllotoxin (1) and compound 4 as an O-hexosyl derivative of podophyllotoxin. PTOX (1) is the major component of the EtOH extract of P. hexandrum rhizomes and roots. On the other hand, this lignan was not found in the extract of D. cymosa roots.
Chromatographic peaks 5 (RT = 3.84 min), 7 (RT = 4.11 min) and 12 (RT = 5.52 min) showed UV spectra similar to those observed for arylnaphtalene lignans (λ
max at 275 nm). The MS spectra registered for these compounds showed peaks at
m/
z 543, for compounds
5 and
7, and
m/
z 379 for compound
12, corresponding to the deprotonated molecules [M − H]
− in the negative mode. The difference of 162 Da between the compound
12 and compounds
5 and
7 indicates the presence of a hexose residue in the latter. The fragmentation of ion at
m/
z 379 in the negative mode originated the daughter ions at
m/
z 319 and 391. These fragments were reported for the lignan diphyllin [
34], which suggest that compound
12 is diphyllin (
24) while compounds
5 and
7 are
O-hexosyl derivatives of diphyllin.
In the UPLC profile of the EtOH extract of rhizomes and roots of P. hexandrum, the peak eluted at 3.84 min (peak 5′) showed a different UV absorption profile, with λmax at 287 nm, characteristic of aryltetralin lignans. The MS spectra associated with this compound showed a peak at m/z 399 [M − H]− and 401 [M + H]+ in the negative and positive ionization modes, respectively. Analysis of the lignan 4′-demethylpodophyllotoxin (6) in the same conditions allowed us to assign this lignan as the compound responsible for peak 5′.
The compound corresponding to peaks 6 (RT = 3.92 min) and 10 (RT = 4.62 min) showed UV absorption pattern similar to what is described for peltatins. The MS spectra associated with chromatographic peak 6 peak showed peaks at m/z 399 and 401 for the deprotonated and protonated molecules, respectively. The MS2 fragmentation of the parent ion at m/z 401, in the positive ionization mode, generated the daughter ion at m/z 247 which is compatible with the neutral loss of a trimethoxybenzyl group (C9H12O3). These results, along with the analysis of the lignan α-peltatin (18) in the same conditions, allowed the identification of compound 6 as α-peltatin (18). The MS spectra associated with peak 10 showed peaks at m/z 413 and 415 for the deprotonated and protonated molecules, respectively. The analysis of the isolated lignan β-peltatin (19) in the same conditions, allowed the identification of compound 10 as β-peltatin (14).
The UV spectra observed for the compound of peak 11 was very similar to that observed for peaks 4 and 8, suggesting that compound 11 (RT = 4.86) is an aryltetralin lignan. The MS spectrum registered for this compound presented peaks at m/z 411 and 413 in the negative and positive modes, respectively. The MS2 fragmentation of the parent ion at m/z 413 was very similar to that observed for the lignan podophyllotoxone (11) (RT = 5.28 min), suggesting that the compound corresponding to peak 11 could be its isomer isopicropodophyllone (14).
The peak 13 (RT = 5.78), which is only present in the EtOH extract of P. hexandrum rhizomes and roots, showed UV spectra compatible with aryltetralin lignans and the MS spectra presented peaks at m/z 444 [M − H + formiate]− and 399 [M + H]+. These results, along with the results obtained for the purified compound allowed the identification of compound 13 as desoxypodophyllotoxin (5).
2.2. In Silico Prediction of Biological Activity of Lignans
The Active-IT system was composed, at the time the calculations were run, of 1987 biological activity datasets modeled with SVM and Naïve Bayes machine learning methods. About 1815 datasets were obtained directly from the PubChem Bioassay database and the remaining 172 datasets were obtained from different sources, including the combination of several PubChem datasets. Some of these modeled datasets were reported before, such as AMES [
35], AChE [
36] and antifungal/antibacterial activities [
37], however most remain unpublished. The complete description of all modeled datasets is far beyond the scope of this paper and will not be discussed in detail.
The target fishing approach was performed using the Active-IT programme [
36]. Before being able to perform predictions of biological activities of the lignans with the Active-IT system, we first made an ultimate validation using some known activities of these compounds. About 12 of the 27 lignans (compounds from
1 to
27) used in this study appear in one or more datasets from the PubChem Bioassay. Among the 1815 PubChem PubChem Bioassay datasets within the Active-IT system about 243 have one or more lignans of the series, with a total of 309 activity points, with 128 classified as active and 181 classified as inactive. For example, the podophyllotoxin (
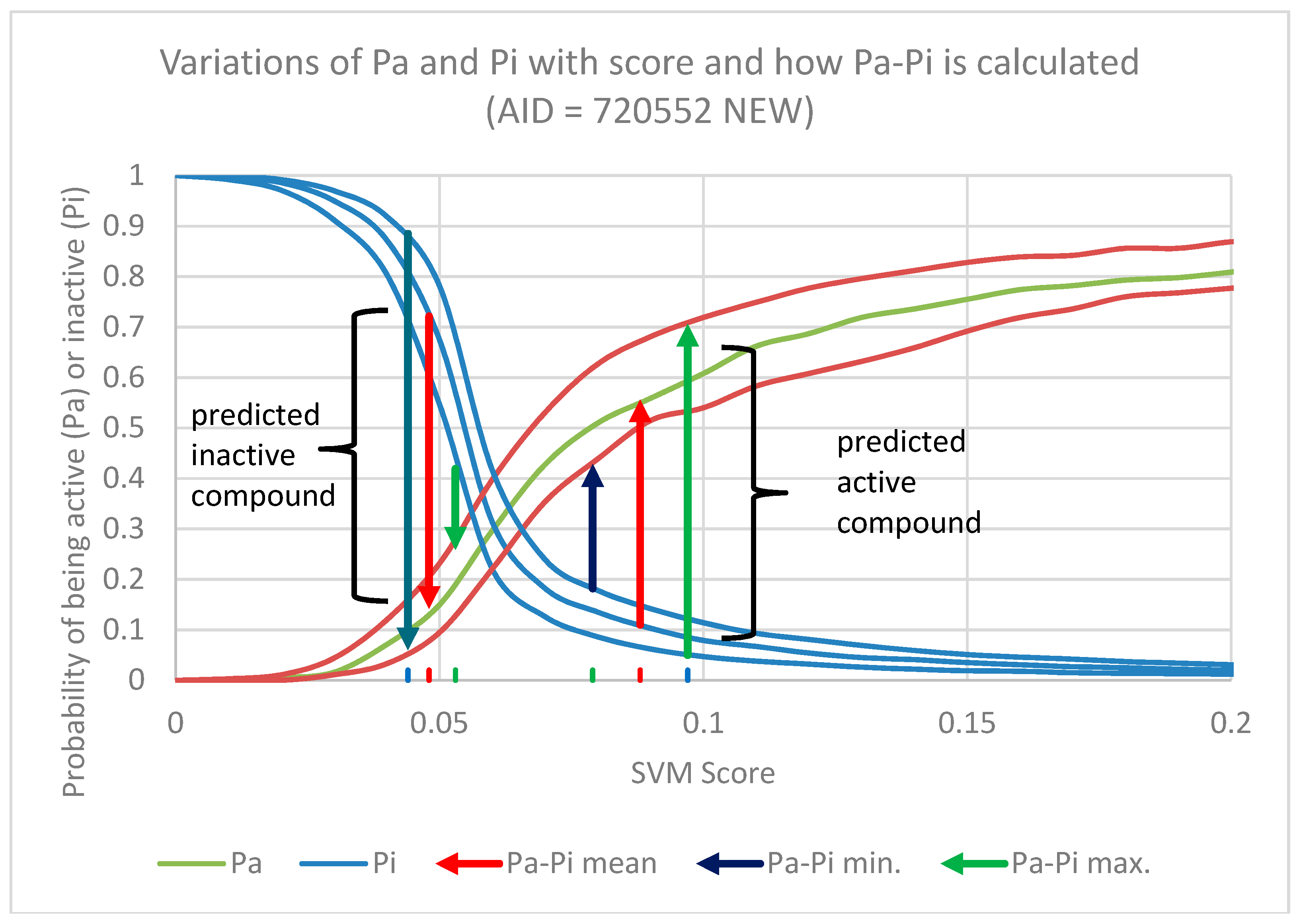
1) appears in 195 different datasets. As expected, the activity predictions of lignans using these models produced excellent results with an AUC of 0.96 for SVM and 0.82 for Naïve Bayes (
Figure 5A and
Table 2).
Therefore, we decided the best approach for validation was to re-build all of these 243 models, excluding the lignans that appear in each dataset and repeat the prediction. The details of all these models, as well as their internal validation are included in the
Supplementary Material (Table S3). The calculated values Pa-Pi of lignans that were excluded from the models are shown in
Table S4 of the Supplementary Material. As expected the prediction was a little worse, but the results are still very good, considering most datasets have a cell-based format, with an AUC of 0.71 for SVM and 0.73 for Naïve Bayes (
Figure 5A and
Table 2). It is worth pointing out that while the SVM method experienced a large decrease in AUC (0.25 AUC units) the Naïve Bayes had a far smaller decrease (0.09 AUC units). This is an evidence that the Naïve Bayes method has a smaller dependence from the input and is less unresponsive to small variations of the dataset composition. The SVM has a lower resilience as it is much more dependent on the input dataset.
However, the validation with AUC only tells us whether the global prediction was accurate or not. In the chemoinformatics it is more important to define whether the most probable active compounds appear in the highest positions in the ranking.
Several metrics can be used to decide the better cutoff to be applied in a classification schema. We used the three metrics based on a contingency matrix to decide the best cutoff to be used:
- (1)
F-score is a measure of the accuracy of the test, calculated by the harmonic mean of recall or sensitivity [TP/(TP + FN)] and precision [TP/(TP + FP)];
- (2)
Matthews Correlation Coefficient (MCC) is a balanced measure of the quality of binary classification and is the most informative single score to establish the quality of a binary classifier prediction in a confusion matrix context [
38];
- (3)
Enrichment Factor (EF) is a measure of how many more active compounds we find relative to a random distribution, it is calculated from the proportion of true active compounds selected in relation to the proportion of true active compounds in the entire dataset [
39].
The complete results of Pa-Pi values calculated for all lignans, as well the predictions made, are included in the
Supplementary material (Table S4). The best cutoff for the classification of lignan compounds as active or inactive was Pa-Pi ≥ 0.15 for both SVM and Naïve Bayes methods, using a positive lower limit of Pa-Pi as an additional filter. In
Table 3 are presented a summary of the results when this threshold value is applied. In this table it is possible to note that SVM outperforms Naïve Bayes method for both datasets in the prediction of active compounds, and both methods present a higher specificity as few inactive compounds are misclassified as active (low false positive rate). This draws attention to the fact that the misclassified inactive compounds are almost the same for datasets with (OLD) or without the lignans (NEW) and, despite the fact that there are several active missed, both methods present a good precision as most of the compounds predicted as active are really active. Both methods lost performance when the lignans are deleted from the modeling dataset, but the effect is more pronounced in SVM, as discussed before for AUC results.
The main approach used in this work to make predictions about the biological activities of lignans was the grouping of several models in activity classes and this procedure must be validated as well. The 243 datasets used in the validation were classified in 137 activity classes, with the more populated being the models associated with leukemia cancer with 16 different datasets. From these 137 activity classes only 125 showed a predominance of active (47 cases) or inactive compounds (78 cases), and 12 showed an equal number of active and inactive. Thus, the predominance of active or inactive lead us to classify 47 classes as “probable active” as the lignans are prone to be active and 78 classes as “probable inactive” as the lignans are prone to be inactive.
To make the prediction, all 27 lignans were submitted to 243 models and the activity class score was calculated as described in the Material and Methods section. To build this score, the individual Pa-Pi values were not taken in account, only the number of cases where the value of Pa-Pi is above the threshold and with a positive minimum value.
The best threshold was determined by analyzing the value of MCC for each threshold (
Table 3), being Pa-Pi ≥ 0.3 for Naïve Bayes and Pa-Pi ≥ 0.25 for SVM. The score of each activity class, in these cases, was calculated by the number of lignans classified as active among all the models belonging to the same class divided by the number of lignans used in the calculations (27) and the square root of the number of models of this class. The division by the square root of the number of models works like a normalization process because as the number of models in the same class increases more compounds are prone to be predicted as active for this class. From our experience, if we divided by the number of models, those classes with a large number of models are penalized, and if we take only the number of compounds the classes with a large number of models are privileged. The division by square root brings some balance to the prediction.
Using this approach we reconstructed the ROC curves using the scores of activity classes and the results are show in the
Figure 5B and
Table 3. As we can see the values of AUC metric although a little smaller than those obtained when we use the Pa-Pi scores of each individual compound are high enough to say that this approach sounds viable and allow us to use it to classify the whole set of lignans instead of analyzing a much larger number of results when we consider each compound independently. It is worth noting that all lignans were used in this activity class validation and not only those for which experimental results are available. Another interesting result was that for Naïve Bayes the AUC increases when the data points are grouped into activity classes in relation to the analysis of all experimental data.
A partial list of predicted biological activities as well as the targets associates with the diseases are described in the
supplementary material (
Table S1 for SVM,
Table S2 for Naïve Bayes and
Table S3 for results when both methods are taken together). In
Table 4 is presented a summary about the activities predicted until the 50 position in the ranking of 753 activity classes using both machine-learning methods (
Table S3 of the Supplementary Material).
2.3. Antibacterial Activity
Infectious diseases, especially those caused by bacteria, are a major concern in several countries mostly due to antimicrobial resistance, which is a global public health issue that could hold back the control of many bacterial diseases [
40]. Therefore, the search for new antimicrobial compounds from natural sources is immensely valuable. The in silico target fishing showed a probable antibacterial activity for aryltetralin lignans, as presented previously.
In our study, the ethanolic extracts from roots of
D. cymosa and from rhizomes and roots of
P. hexandrum were the most effective extracts against Gram-positive bacteria, reaching total inhibition of the microbial growth against
B. cereus and proeminent inhibition rates (58.43 ± 2.7% and 67.56 ± 0.8%, respectively) against
S. aureus (
Table 5). PTOX also presented antibacterial activity against
S. aureus (53.34 ± 8.8%) and
E. coli (51.57 ± 9.08%). No antibacterial activity of the extracts and PTOX was observed against the Gram-negative pathogens EHEC,
P. aeruginosa and
Salmonella Typhi.
2.4. Anticholinesterasic Activity
We evaluated the AChE inhibitory activity of
D. cymosa and
P. hexandrum extracts as well as of PTOX. All extracts showed promising inhibition of AChE activity in the quantitative assay. The EtOH extract of leaves from
D. cymosa showed the higher percentage of inhibition at 400 μg/mL (
Table 6). In the bioautographic assay, the extract of
P. hexandrum showed a higher intensity of white spots, and this result did not correspond to the quantitative assay. This may have happened due to the possibility of false-positive results when the evaluated extract presents some classes of secondary metabolites, such as tannins and phenolics, which do not directly inhibit the enzyme but can induce enzymatic denaturation [
57,
58]. On the other hand, the in silico studies did not confirm the AChE inhibition activity (position 577 in 753 models) by the group of 27 lignans investigated in the present study.
2.5. Antioxidant Activity
Although, inhibition of AChE is still considered as the main therapeutic strategy to treat Alzheimer’s disease, other events are implicated in the physiopathology of this disease. The role of oxygen reactive species (ROS) have been extensively investigated. Therefore, we evaluated the antioxidant potential of the EtOH extracts of D. cymosa leaves, roots and P. hexandrum rhizomes and roots.
The extract of leaves from
D. cymosa showed antioxidant activity only in the β-carotene/linoleic acid co-oxidation assay, while the extract of roots showed promising antioxidant activity in all evaluated models. The extract of
P. hexandrum roots and rhizomes presented significant antioxidant potential in all evaluated models. However, the isolated lignan PTOX (
1) showed no antioxidant activity (
Table 6).
2.6. Cytotoxicity
The results of the cytotoxicity assay, performed in THP-1 cells using the SRB method, indicated a moderate toxicity for the extracts of leaves from
D. cymosa and of roots and rhizomes from
P. hexandrum, which showed cell viability lower than 80% at 200 and 400 μg/mL, with CC
50 of 368.0 ± 13.8 and 338.9 ± 15.1 µg/mL (
Table 6). On the other hand, the extract of roots from
D. cymosa was cytotoxic at all tested concentrations, with a CC
50 value of 100.0 ± 5.3 µg/mL. The lignan PTOX (
1) showed a CC50 value of 400.0 ± 10.3 µg/mL (
Table 6). In silico prediction indicated a small probability of lignans to present THP-1 cytotoxicity (position 316 in 753 activity classes).
3. Discussion
A broad range of biological activities have been associated with lignans, thus making them an interesting class of secondary metabolites. Even though lignans are known for their toxicity, other biological activities of lignan-rich plant extracts are worth investigating.
The chromatographic characterization of the phenolic content of the EtOH extracts of
D. cymosa leaves,
D. cymosa roots and
P. hexandrum rhizomes and roots indicated a similar chemical composition for the three extracts. The caffeoylquinic acid was only identified in
D. cymosa leaves, while PTOX hexoside, deoxypodohyllotoxin (
5) and 4′-demethylpodohyllotoxin (
6) were only found in
P. hexandrum. PTOX was found in the EtOH extracts of
D. cymosa leaves and
P. hexandrum rhizomes and roots, being the major constituent of the latter. The EtOH extract of roots from
D. cymosa showed the best antioxidant potential among the evaluated extracts. These results can be explained by the presence of kaempferol as the major constituent of this extract. Numerous studies have shown that flavonoids, such as quercetin and kaempferol, and their heterosides have a wide range of biological activities, including antioxidant, anti-inflammatory and antimicrobial activities [
59,
60]. Recently, Wang and co-workers [
61] observed a DPPH and ABTS radical scavenging activity for kaempferol as well as an inhibition of concanavalin A (Con A)-induced NO or ROS production in LPS-induced RAW 264.7 macrophage cells [
61]. In another study, kaempferol was able to scavenge the superoxide anion, hypochlorous acid, chloramine and nitric oxide [
62] as well as showed scavenging ability on superoxide anion produced by electrochemical reduction of oxygen [
63].
The lignan content of
Podophyllum and
Diphylleia species varies both qualitatively and quantitatively, according to the data previously published. UPLC-DAD-MS methods are largely employed for the identification and quantification of lignans in the aforementioned species [
64,
65,
66,
67]. Sharma and Arora identified four aryltetralin lignans in the MeOH extract of rhizomes from
P. hexandrum [
64]. In another study, Sharma and Kumar evaluated the extracts of leaves and roots of
P. hexandrum obtained from different locations by HPLC-ESI-MS, and found that the podophyllotoxin content was twice as high in the roots in comparison with the content found in the leaves of
P. hexandrum found in high altitudes [
65]. Avula e coworkers evaluated the content of podophyllotoxin (
1), 4′-demethylpodophyllotoxin (
6), α-peltatin (
18) and β-peltatin (
19) in samples from
P. peltatum. 4′-demethylpodophyllotoxin (
6) and α-peltatin (
18) were the main lignans observed for this species, while the content of PTOX varied from 0.004–0.77% when plants colletcted from various colonies within the same site were evaluated [
66]. UPLC-ESI-MS methods can also be employed for the pharmacokinetic studies. The lignans podophyllotoxin (
1), 4-
epi-podophyllotoxin (
2), and 4′-demethylpodophyllotoxin (
6) were simultaneously evaluated in rat plasma using a UPLC-ESI-MS method after oral administration of the EtOH extract of
Diphylleia sinensis, 367 mg/kg, to Wistar rats [
67]. It is noteworthy to mention that this is the first report of the characterization of flavonoids and caffeoylquinic acid in
D. cymosa.
Some predictions of biological activities observed in this study were in accordance with the ethnopharmacological uses for both plant species, as well as for the other isolated
podophyllum lignans. However, some new predict activities such as angiogenesis, osteoporosis, myotonic dystrophy and autoimmune diseases were also observed (
Table 4). A reasonable agreement could be noted between predictions made with SVM and Naïve Bayes modeling methods, although the use of both methods can produce results that are more reliable, as indicated in the validation step. Thus, all predictions discussed below will be based in a unique rank of 753 activity classes, where the best rank between both methods and the averaged rank were used to produce the final ordered list.
The in silico approach showed a high probability of lignans have an anti-inflammatory activity (position 1 in 753 activity classes). This activity has been described before for lignans [
11]. The most probable target related with this activity is NF-κB activation. The nuclear factor NF-κB pathway has been considered a classical proinflammatory signaling pathway [
68].
The cytotoxic activity of lignans has been much explored, as well as their mechanism of action. The results from in silico prediction showed a low cytotoxicity against THP-1 cells (316/753), which is in accordance with our experimental results. Podophyllotoxin have been evaluated in several models using THP-1 cells, with moderate to low toxicity reported. The effect of podophyllotoxin on IL-1β and TNF expression was evaluated using THP-1 cells at 10 µM and no cytotoxicity was observed [
69]. However, according with our calculations, lignans could present cytotoxicity against human lymphoblastic (position 2 in 753 activity classes), isogenic chicken DT 40 (position 19 in 753 activity classes), HEK293 (position 32 in 753 activity classes) and MAGI-CCR5 (position 50 in 753 activity classes) cells. Derivatives have been synthetized [
11] to obtain new antitumour compounds. Many targets involved in cancer therapy have also been predicted as potential targets for lignans, such as AP1 endonuclease (position 6 in 753 activity classes), tumor antigen p53 (position 7 in 753 activity classes), GLI family zinc (position 11 in 753 activity classes), RecQ-like DNA helicase 1 (position 14 in 753 activity classes). Other targets can be found in
Table 4 and
Table S3 of Supplementary Material.
The lignans also showed high probability to be active against
Salmonella typhimurum (position 3 in 753 activity classes),
Mycobacterium tuberculosis (position 4 in 753 activity classes),
Staphylococcus aureus (position 23 in 753 activity classes),
Pseudomonas aeruginosa (position 18 in 753 activity classes) and
Escherichia coli (position 52 in 753 activity classes). In this study, all these microorganisms were evaluated, with the exception of
M. tuberculosis. These predictions are in agreement with ours in vitro results from the antibacterial assay. The activity against
M. tuberculosis is consistent with the ethnopharmacological use in the Eastern world folk medicine [
42].
Regarding the in vitro antibacterial activity, the ethanolic extracts from rhizomes and roots of
P. hexandrum and from roots of
D. cymosa were the most effective samples against Gram-positive bacteria. PTOX (1) showed significative activity against
S. aureus and
E. coli, while no antibacterial activity was observed against the other Gram-negative pathogens. The antibacterial activity of lignans have already been reported. Nanjundaswamy and coleagues [
70] reported a relevant antibacterial activity of two synthetic precursors of PTOX against
E. coli,
P. aeruginosa and
Salmonella Typhi. Other authors have also indicated antibacterial activity of extracts from the rhizomes of
P. hexandrum [
71] and analogues of PTOX against
P. aeruginosa [
72], contrasting the results of the present study. This finding supports the ethnopharmacologial uses of both plant species. No mention of antimicrobial activity from
D. cymosa was reported so far.
Interestingly, PTOX did not elicit a higher inhibition of the growth of most of the evaluated microorganisms, in comparison with the crude extracts. This is probably due to the synergistic effect of secondary metabolites in the crude extracts.
Considering
S. typhimurum, one target related to the predicted activity is the
PhoP regulation. This target is composed by two genes
PhoP and
PhoQ, associated with virulence, survival inside the macrophages and defensing resistance of
S. typhimurum [
73]. Against
M. tuberculosis, the possible target is a transaminase BioA, an enzyme involved in biotin biosynthesis, representing a potential target to develop new antitubercular agents [
74]. The putative target involved in the activity against
S. aureus is the Quorum sensing (QS), defined by Reuter and co-workers [
75] as the exchange of chemical signals in bacterial populations, that depends on the bacterial density. QS is responsible for virulence in the clinically relevant bacteria. It has been suggested as a promising target for developing new anti-infective compounds. It was not found a specific target related to
E. coli.
The extracts of roots from D. cymosa and rhizomes and roots of P. hexandrum showed the best antioxidant potential, while the extract of leaves from D. cymosa showed an anticholinesterasic activity. All the extracts showed moderate toxicity to the THP-1 cells, and the cytotoxic activity observed for the extracts of leaves of D. cymosa and rhizomes and roots of P. hexandrum were similar to that observed for PTOX (1), which did not exhibit antioxidant and anticholinesterase activity. These results indicate that PTOX (1) could be the major cytotoxic lignan in these extracts, while other phenolic constituents could be responsible for the antioxidant and anticholinesterase activities observed. However, in the extract of roots of D. cymosa, which showed the lowest CC50 value, the lignan PTOX (1) was not found, indicating that other unidentified minor compound is the responsible for the observed cytotoxicity.
According to the in silico prediction (position 49 in 753 models), the lignans can act DNA damage-inducible transcript 3 protein (C/EBP homologous protein, CHOP) which has been proposed as a target of treatments for some neurodegenerative diseases as Alzheimer’s diseases [
52]. Furthermore, according to Naïve Bayes prediction (
Table S2 in the Supplementary Material) the lignans can possibly act over the protein Tau indicating a potential application to treat the Alzheimer’s disease and other diseases [
76].
Another important effect of lignans is the immunosuppressive activity what can be associated to their uses as therapeutic agents against psoriasis and rheumatoid arthritis, as well to prevent the acute rejection of transplanted organs [
77,
78]. One putative target that could explain this activity is the sphingosine 1-phosphate receptor 1, predict as potential target for lignans (position 20 in 753 activity class), due to its involvement in immune system modulation [
79].
The anti-viral activity of lignans is well known since it was first cited in 1942 as a treatment for veneral wart (
Condyloma acuminatum), an ailment caused by a papilloma virus [
80]. There have been reported effects against HIV, herpes simplex, influenza, vaccinia viruses, and measles [
42]. These results also confirm our in silico predictions as the activities against HIV-1 and herpes appear in position 9 and 42, respectively, among 753 activity classes.
The hypolipidemic properties of lignans, as reported by Iwasaki and co-workers [
81] are in consonance with our prediction as potential target for these compounds the 1-acylglycerol-3-phosphate
O-acyltransferase, a protein activator of the lipase Atgl [
82], predict in position 41 among 753 activity classes.
There are no reports in literature of the evaluation of the anticholinesterase activity of Berberidaceae plants or of the lignan PTOX (1), but other lignans have already demonstrated in vitro AChE inhibitory activity [
83]. In the study by Hung and co-workers [
84], sixteen lignans were isolated from
Schizandra chinensis and were evaluated for the inhibition of AChE in vitro. Among the evaluated compounds, only five were active with IC
50 lower than 15 μM. Schisandrin was evaluated in vivo by Itoh et al. [
85] and was active at 3 mg/kg. El-Hassan et al. [
86] demonstrated that Syringaresinol inhibited AChE in vitro with an IC
50 value of 200 μg/mL. Still exploring the investigation of AChE inhibition activity by lignans, Salleh et al. [
87], isolated five lignans from the stem extract of
Beilschmiedia pulverulenta, which were evaluated in vitro in the microplate inhibition assay for the AChE, showing have IC
50 values in the range of 179.8 to 504 μM.
Regarding the antioxidant capacity of these compounds, studies by Wang et al. [
88] evaluated the ability of the extracts of
S. chinensis and
S. sphenanthera to scavenge the DPPH radical. The authors suggested that variations in lignan content between the extracts lead to different antioxidant activities. The
S. chinensis species showed higher activity due to the higher content of the lignans Schisandrol A and B and Schinsandrin B.
In a study by Dar et al. [
63], the antioxidant capacity of PTOX (
1) was evaluated in the DPPH sequestration and the TBARS lipid peroxidation assays. In both experiments, PTOX presented IC
50 value higher than 250 μg/mL.
These results, along with the data found in literature, indicates that PTOX (1) is probably not involved in the antioxidant activity observed for P. hexandrum and D. cymosa extracts, but other lignans or other phenolic constituents may account for the observed activity.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}