Lung Cancer Chemopreventive Activity of Patulin Isolated from Penicillium vulpinum

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
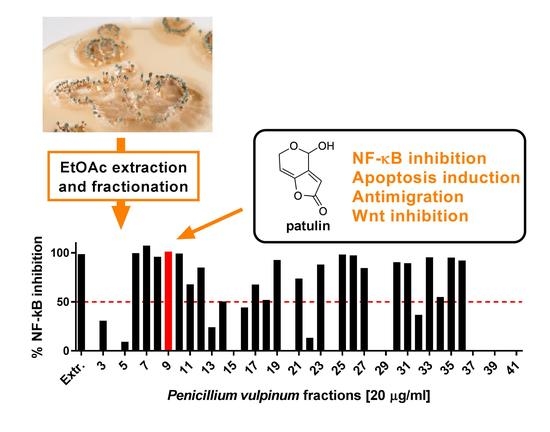
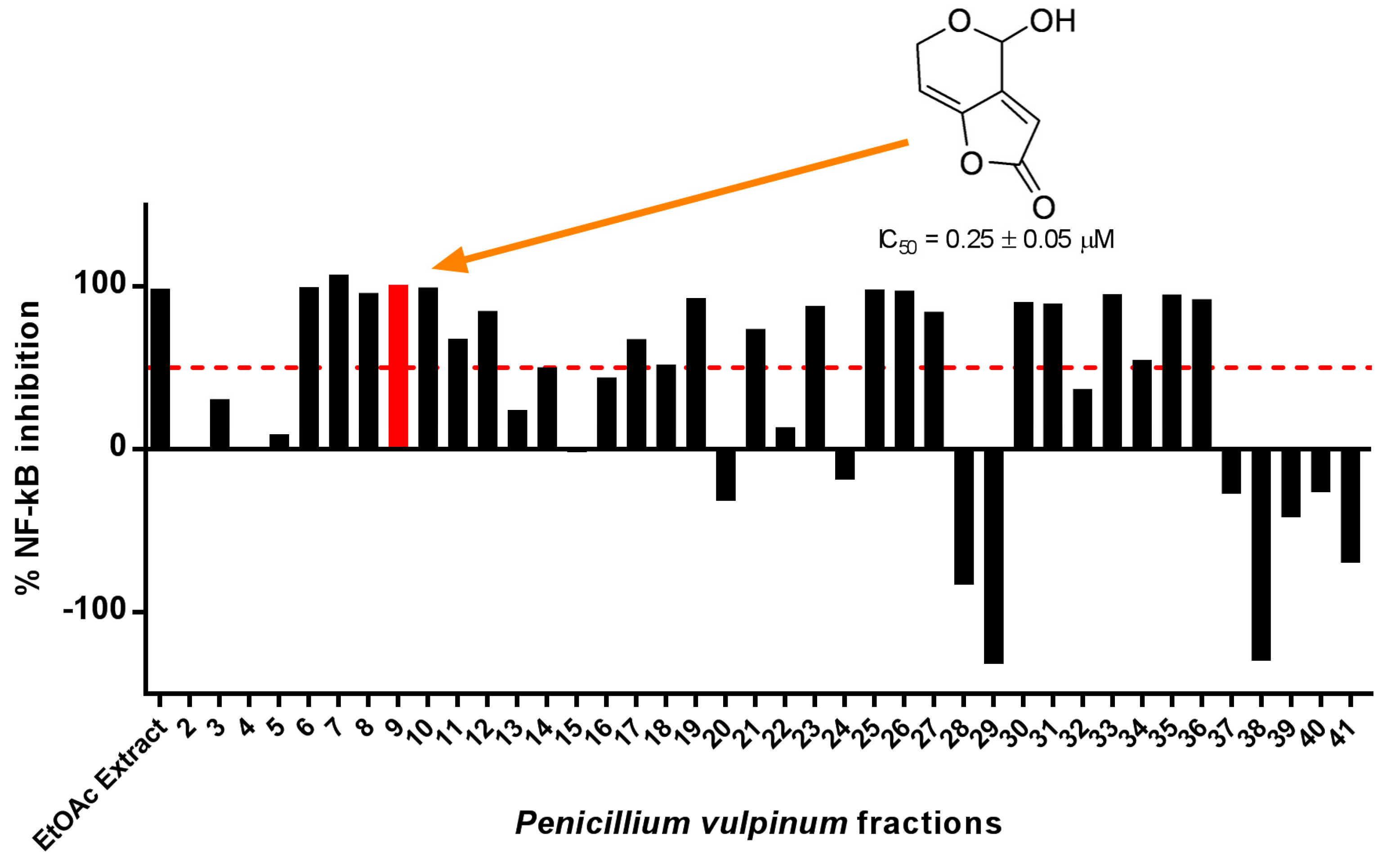
2.1. Patulin Isolation through Bioassay-Guided Fractionation
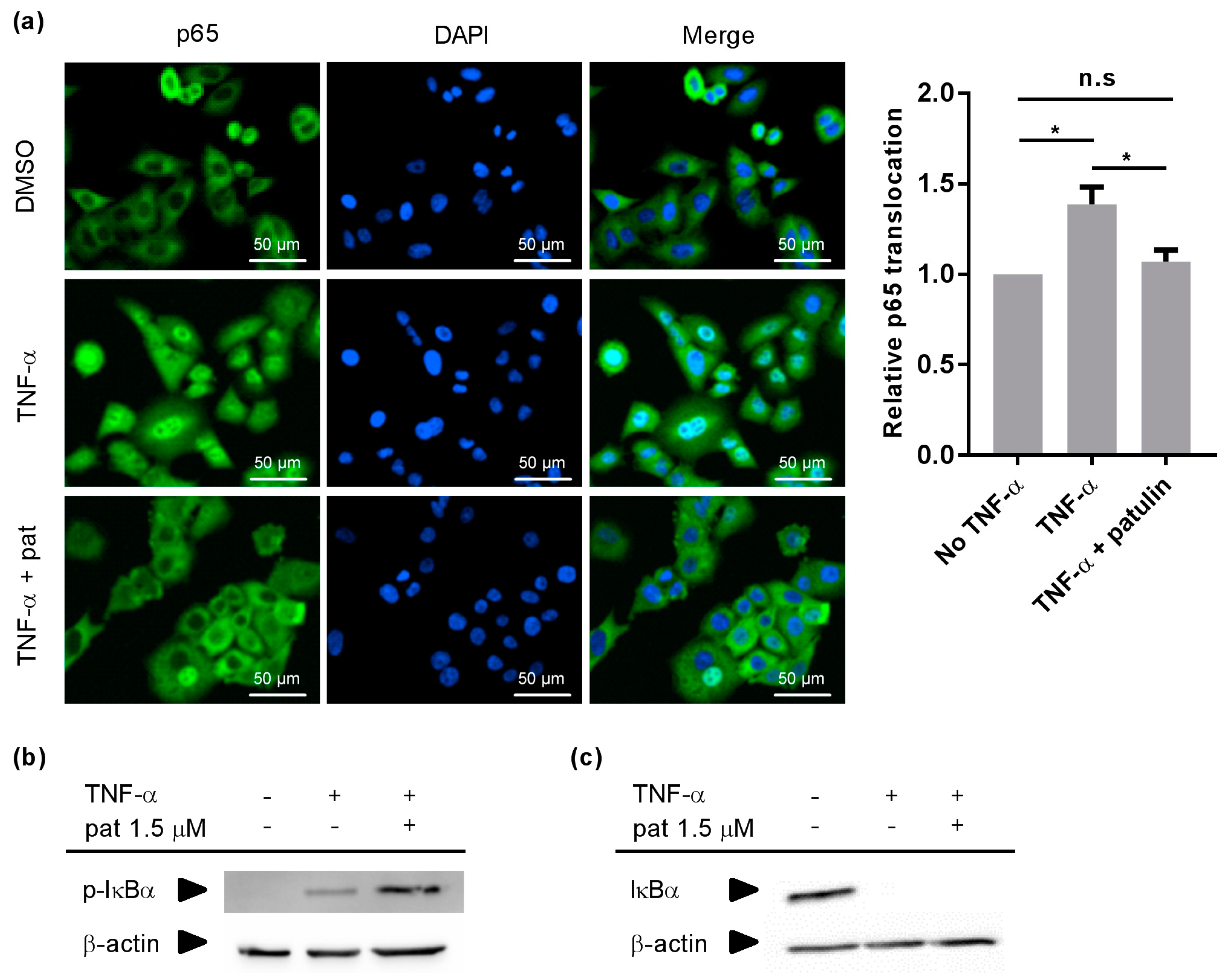
2.2. Patulin Inhibited NF-κB p65 Nuclear Translocation
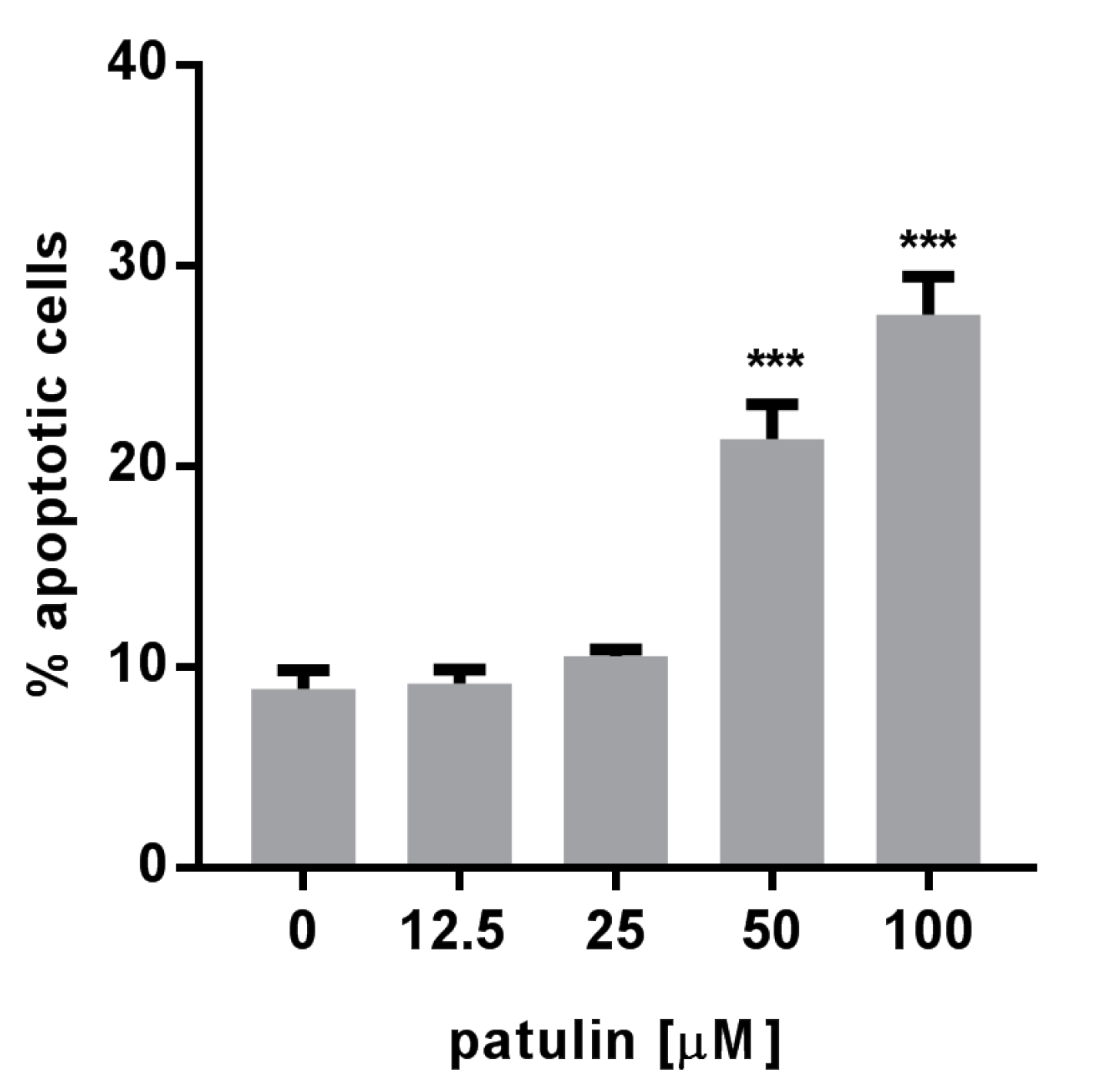
2.3. Patulin Triggered Apoptotic Cell Death in A549 Cells
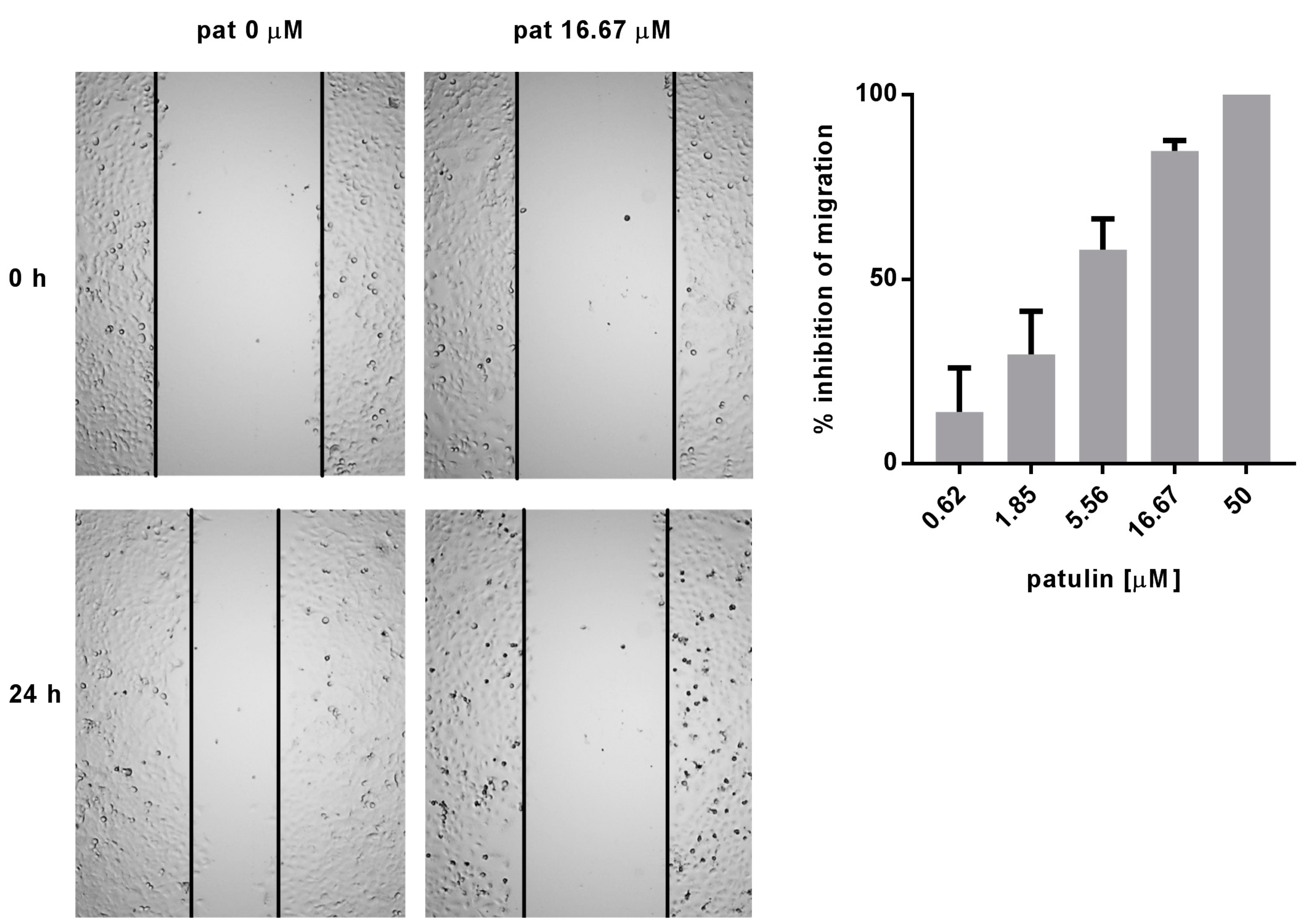
2.4. Patulin Inhibited A549 Cell Migration
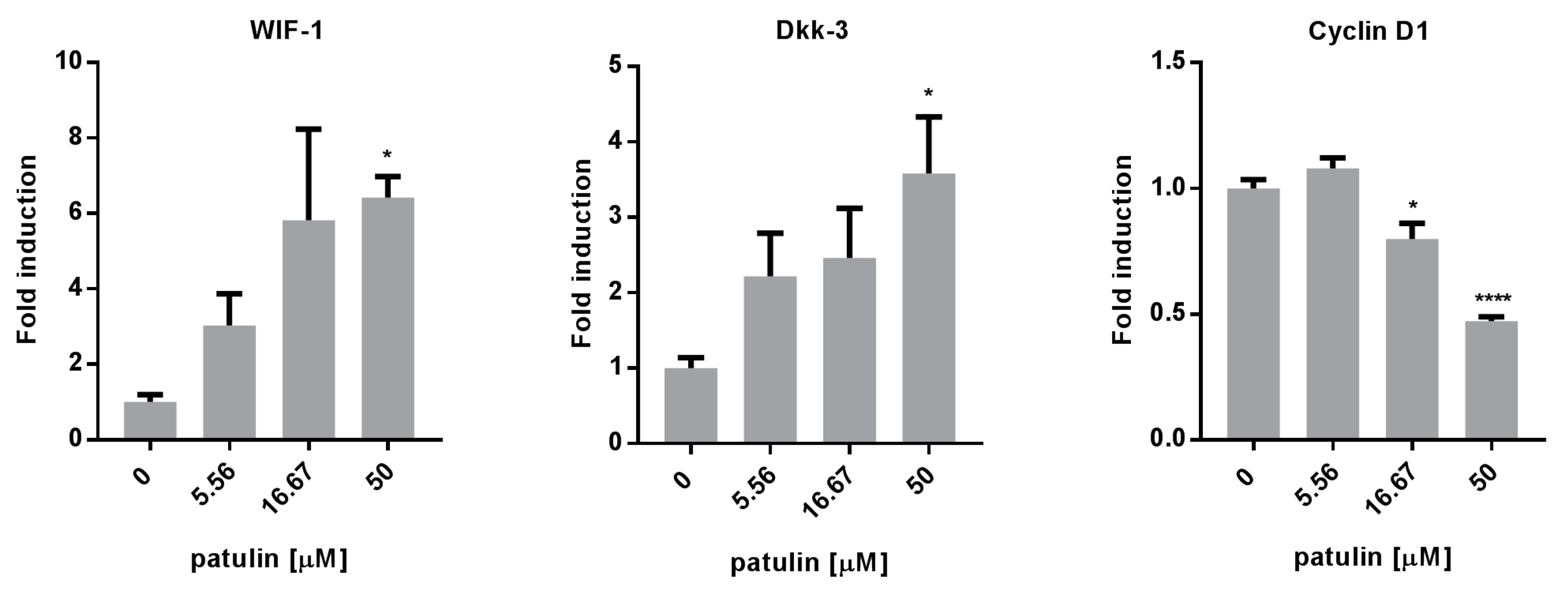
2.5. Patulin Inhibited the Wnt Pathway
3. Discussion
4. Materials and Methods
4.1. Fungal Culture
4.2. Extraction and Isolation of Patulin
4.3. NMR and HRMS Measurements
4.4. Cell Culture
4.5. Measure of NF-κB Activity
4.6. Immunocytochemistry
4.7. Western Blot Analysis
4.8. Cell Viability Assay
4.9. Apoptosis Assay
4.10. Cell Migration Assay
4.11. Quantitative Real Time PCR Analysis of mRNA Expression
4.12. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Forouzanfar, M.H.; Afshin, A.; Alexander, L.T.; Anderson, H.R.; Bhutta, Z.A.; Biryukov, S.; Brauer, M.; Burnett, R.; Cercy, K.; Charlson, F.J.; et al. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1659–1724. [Google Scholar] [CrossRef]
- Gravely, S.; Giovino, G.A.; Craig, L.; Commar, A.; D’Espaignet, E.T.; Schotte, K.; Fong, G.T. Implementation of key demand-reduction measures of the WHO Framework Convention on Tobacco Control and change in smoking prevalence in 126 countries: An association study. Lancet Public Health 2017, 2, e166–e174. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Ben-Neriah, Y.; Karin, M. Inflammation meets cancer, with NF-κB as the matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sung, B. NF-κB in cancer: A matter of life and death. Cancer Discov. 2011, 1, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Yamamoto, Y.; Wang, Q.M. The IKK NF-κB system: A treasure trove for drug development. Nat. Rev. Drug Discov. 2004, 3, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Liu, D.; Shishodia, S.; Ozburn, N.; Behrens, C.; Lee, J.J.; Hong, W.K.; Aggarwal, B.B.; Wistuba, I.I. Nuclear factor-κB (NF-κB) is frequently expressed in lung cancer and preneoplastic lesions. Cancer 2006, 107, 2637–2646. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Ogata, H.; Nishigaki, R.; Broide, D.H.; Karin, M. Tobacco smoke promotes lung tumorigenesis by triggering IKKβ and JNK1 dependent inflammation. Cancer Cell 2010, 17, 89. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, T.D.; Herscovitch, M. Inhibitors of NF-κB signaling: 785 and counting. Oncogene 2006, 25, 6887–6899. [Google Scholar] [CrossRef] [PubMed]
- Tada, M.; Ohtsu, K.; Chiba, K. Synthesis of patulin and its cyclohexane analogue from furan derivatives. Chem. Pharm. Bull. 1994, 42, 2167–2169. [Google Scholar] [CrossRef]
- Chen, G.; Bhojani, M.S.; Heaford, A.C.; Chang, D.C.; Laxman, B.; Thomas, D.G.; Griffin, L.B.; Yu, J.; Coppola, J.M.; Giordano, T.J.; et al. Phosphorylated FADD induces NF-κB, perturbs cell cycle, and is associated with poor outcome in lung adenocarcinomas. Proc. Natl. Acad. Sci. USA 2005, 102, 12507–12512. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Smedsgaard, J.; Frisvad, J.C. Penicillium expansum: Consistent production of patulin, chaetoglobosins, and other secondary metabolites in culture and their natural occurrence in fruit products. J. Agric. Food Chem. 2004, 52, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Boussabbeh, M.; Salem, I.B.; Rjiba-Touati, K.; Bouyahya, C.; Neffati, F.; Najjar, M.F.; Bacha, H.; Abid-Essefi, S. The potential effect of patulin on mice bearing melanoma cells: An anti-tumour or carcinogenic effect? Tumor Biol. 2016, 37, 6285–6295. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Soung, N.K.; Thimmegowda, N.R.; Jeong, S.J.; Jang, J.H.; Moon, D.-O.; Chung, J.K.; Lee, K.S.; Kwon, Y.T.; Erikson, R.L.; et al. Patulin induces colorectal cancer cells apoptosis through EGR-1 dependent ATF3 up-regulation. Cell. Signal. 2012, 24, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.-T.; Lo, Y.-C.; Wu, M.-S.; Li, C.-Y.; Kuo, Y.-P.; Lai, Y.-H.; Tsai, Y.; Chen, K.-C.; Chuang, T.-H.; Yao, C.-H.; et al. Mycotoxin patulin suppresses innate immune responses by mitochondrial dysfunction and p62/sequestosome-1-dependent mitophagy. J. Biol. Chem. 2016, 291, 19299–19311. [Google Scholar] [CrossRef] [PubMed]
- Bredel, M.; Scholtens, D.M.; Yadav, A.K.; Alvarez, A.A.; Renfrow, J.J.; Chandler, J.P.; Yu, I.L.Y.; Carro, M.S.; Dai, F.; Tagge, M.J.; et al. NFKBIA deletion in glioblastomas. N. Engl. J. Med. 2011, 364, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Meylan, E.; Oliver, T.G.; Feldser, D.M.; Winslow, M.M.; Bronson, R.; Jacks, T. Response and resistance to NF-κB inhibitors in mouse models of lung adenocarcinoma. Cancer Discov. 2011, 1, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.J. Wnt signaling pathway in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.X.; Chiang, A.C.; Zhang, X.H.F.; Kim, J.Y.; Kris, M.G.; Ladanyi, M.; Gerald, W.L.; Massagué, J. WNT/TCF signaling through LEF1 and HOXB9 mediates lung adenocarcinoma metastasis. Cell 2009, 138, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Yang, H.-P.; Zhou, X.-D.; Gong, L.; Tang, C.-L.; Wang, H.-J. The hypomethylation agent bisdemethoxycurcumin acts on the WIF-1 promoter, inhibits the canonical Wnt pathway and induces apoptosis in human non-small-cell lung cancer. Curr. Cancer Drug Targets 2011, 11, 1098–1110. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, T.; Nozaki, I.; Miyazaki, M.; Sakaguchi, M.; Pu, H.; Hamazaki, Y.; Iijima, O.; Namba, M. Antiproliferative activity of REIC/Dkk-3 and its significant down-regulation in non-small-cell lung carcinomas. Biochem. Biophys. Res. Commun. 2001, 289, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Lemjabbar-Alaoui, H.; Dasari, V.; Sidhu, S.S.; Mengistab, A.; Finkbeiner, W.; Gallup, M.; Basbaum, C. Wnt and Hedgehog are critical mediators of cigarette smoke-induced lung cancer. PLoS ONE 2006, 1, e93. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Green, B.A.; Farr, J.M.; Lopes, F.C.M.; Van Raay, T.J. Wnt drug discovery: Weaving through the screens, patents and clinical Trials. Cancers 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chen, J.; He, J.; Li, J.; Shi, J.; Cho, W.C.; Liu, X. Wnt signaling as potential therapeutic target in lung cancer. Expert Opin. Ther. Targets 2016, 20, 999–1015. [Google Scholar] [CrossRef] [PubMed]
- Puel, O.; Galtier, P.; Oswald, I.P. Biosynthesis and toxicological effects of patulin. Toxins 2010, 2, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.V.A.; Malainer, C.; Schwaiger, S.; Hung, T.; Atanasov, A.G.; Heiss, E.H.; Dirsch, V.M.; Stuppner, H. Screening of vietnamese medicinal plants for NF-κB signaling inhibitors: Assessing the activity of flavonoids from the stem bark of Oroxylum indicum. J. Ethnopharmacol. 2015, 159, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.-C.; Park, A.Y.; Guan, J.-L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteillier, A.; Allard, P.-M.; Gindro, K.; Wolfender, J.-L.; Cuendet, M. Lung Cancer Chemopreventive Activity of Patulin Isolated from Penicillium vulpinum. Molecules 2018, 23, 636. https://doi.org/10.3390/molecules23030636
Monteillier A, Allard P-M, Gindro K, Wolfender J-L, Cuendet M. Lung Cancer Chemopreventive Activity of Patulin Isolated from Penicillium vulpinum. Molecules. 2018; 23(3):636. https://doi.org/10.3390/molecules23030636
Chicago/Turabian StyleMonteillier, Aymeric, Pierre-Marie Allard, Katia Gindro, Jean-Luc Wolfender, and Muriel Cuendet. 2018. "Lung Cancer Chemopreventive Activity of Patulin Isolated from Penicillium vulpinum" Molecules 23, no. 3: 636. https://doi.org/10.3390/molecules23030636
APA StyleMonteillier, A., Allard, P.-M., Gindro, K., Wolfender, J.-L., & Cuendet, M. (2018). Lung Cancer Chemopreventive Activity of Patulin Isolated from Penicillium vulpinum. Molecules, 23(3), 636. https://doi.org/10.3390/molecules23030636