Transcript Profiling and Gene Identification Involved in the Ethylene Signal Transduction Pathways of Creeping Bentgrass (Agrostis stolonifera) during ISR Response Induced by Butanediol


Abstract
:1. Introduction
2. Results
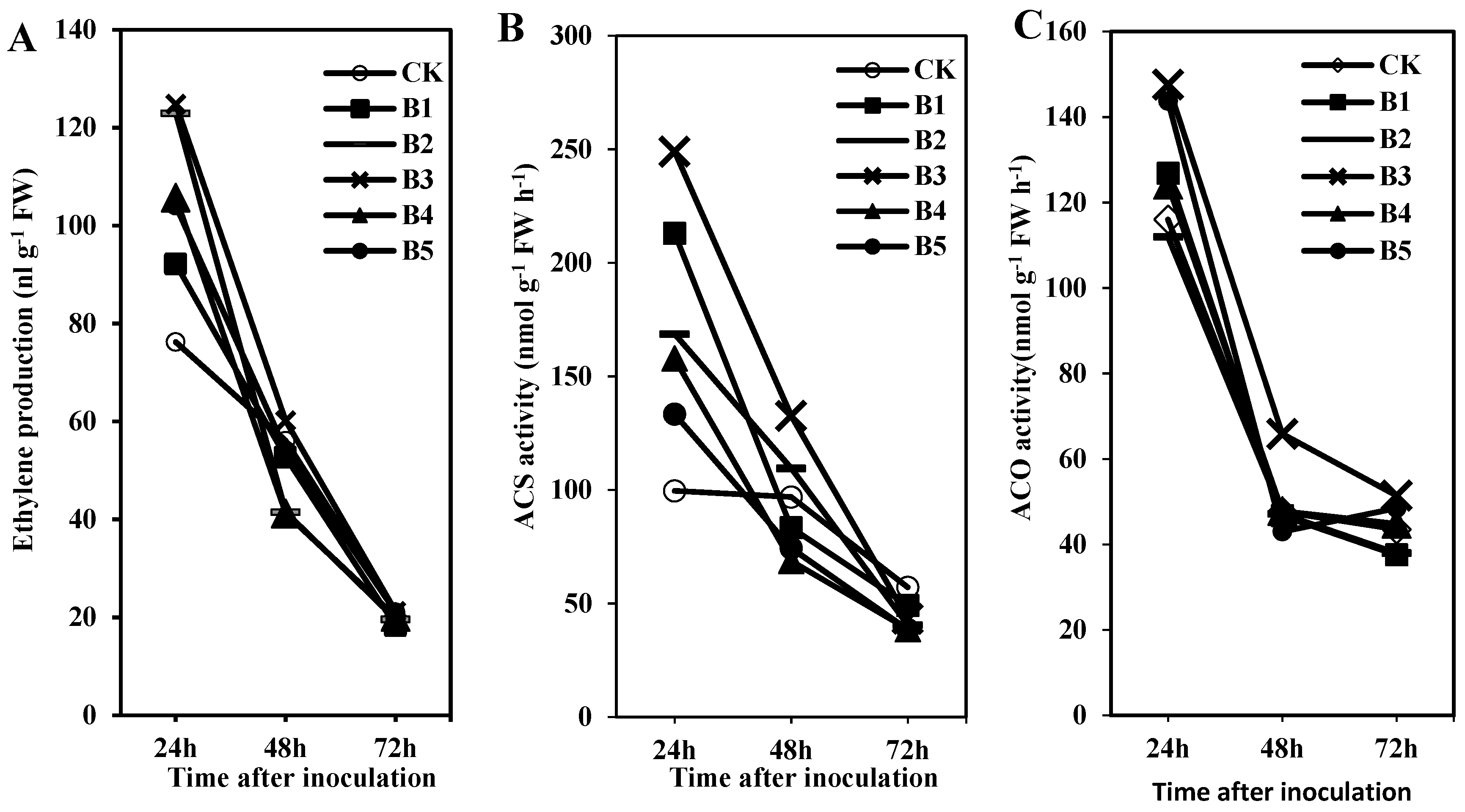
2.1. Ethylene Production and Related Enzyme Activities in Creeping Bentgrass under the ISR Response Were Induced by BDO
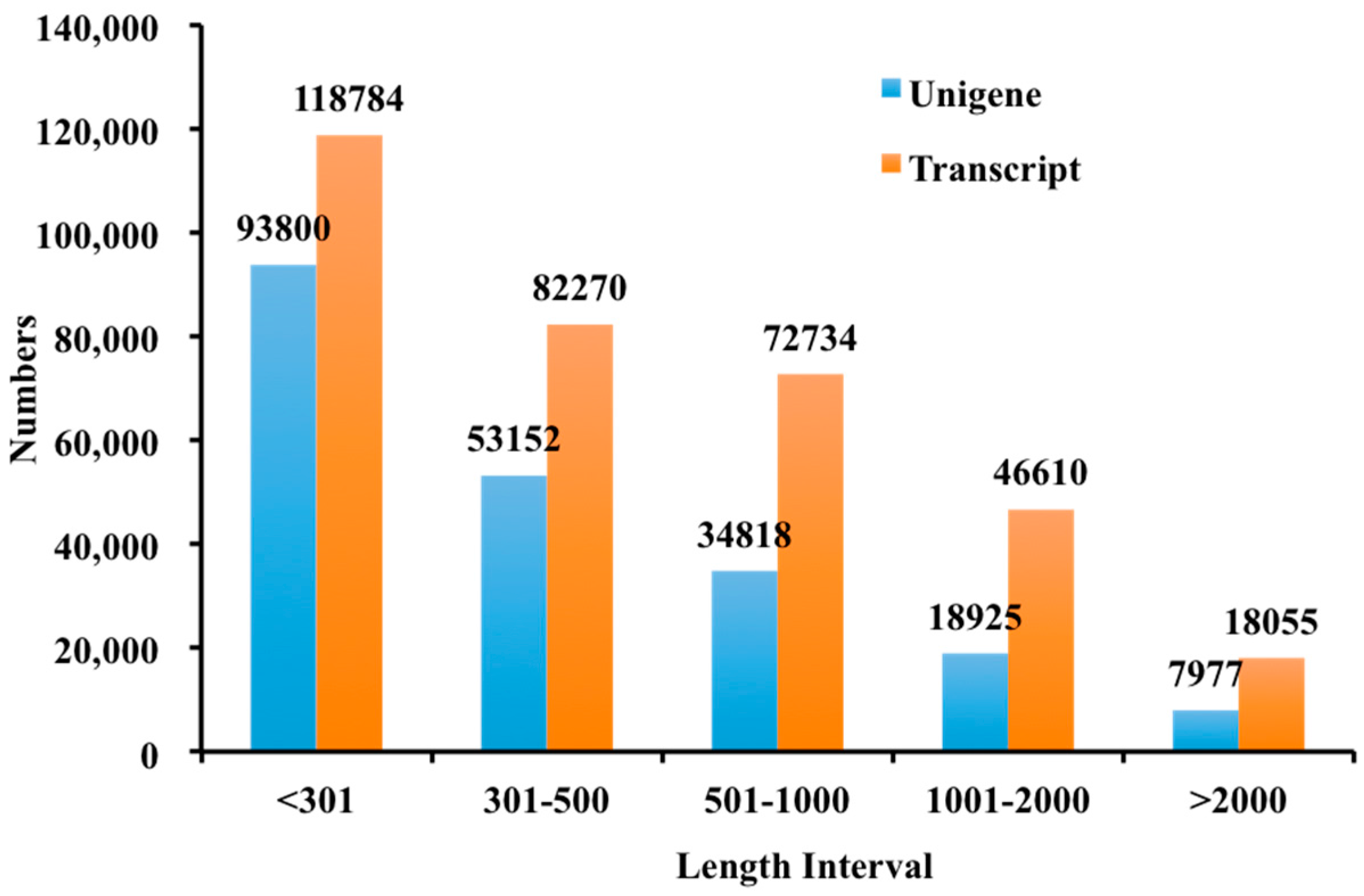
2.2. Illumina Sequencing and De Novo Assembly
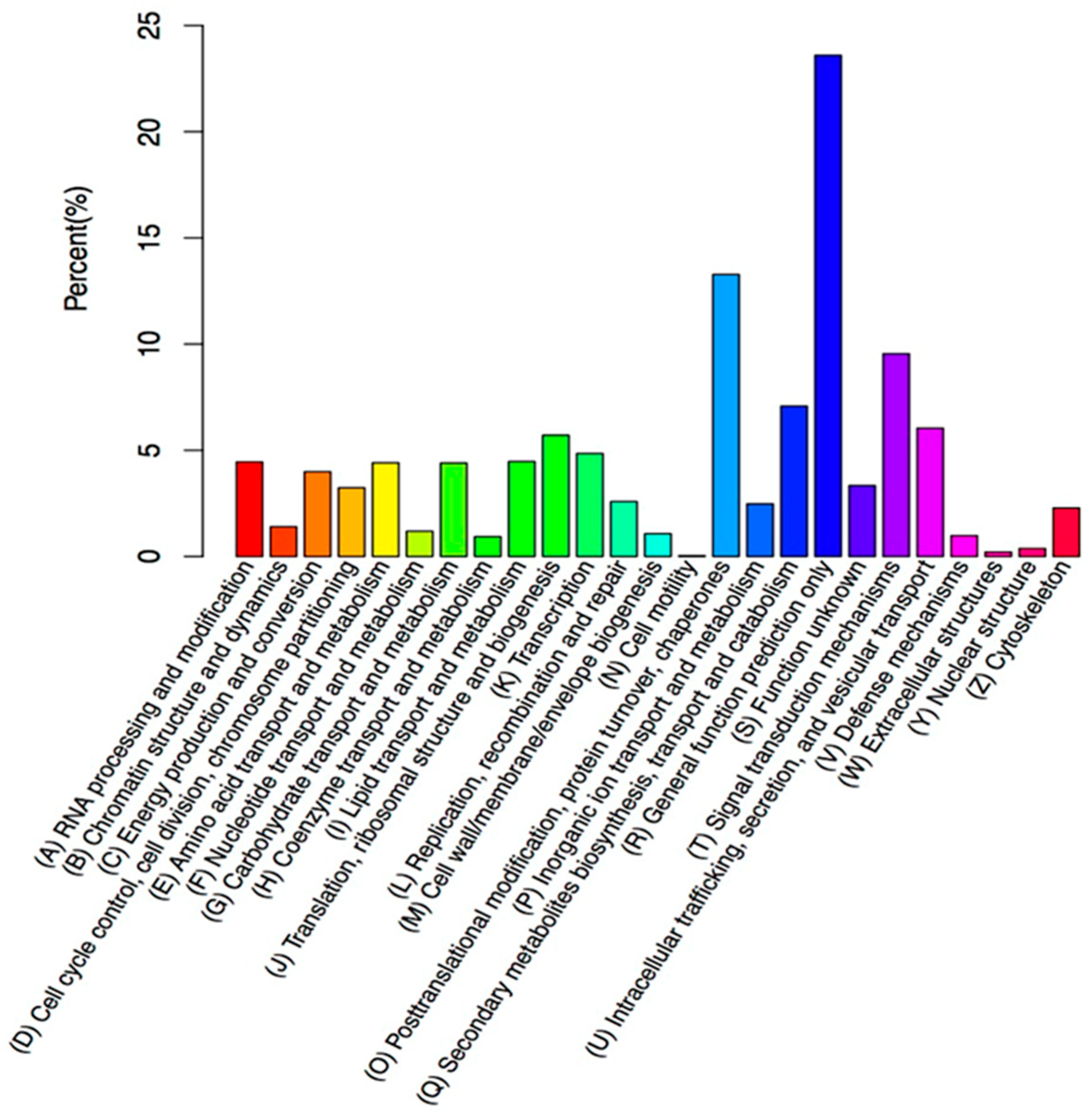
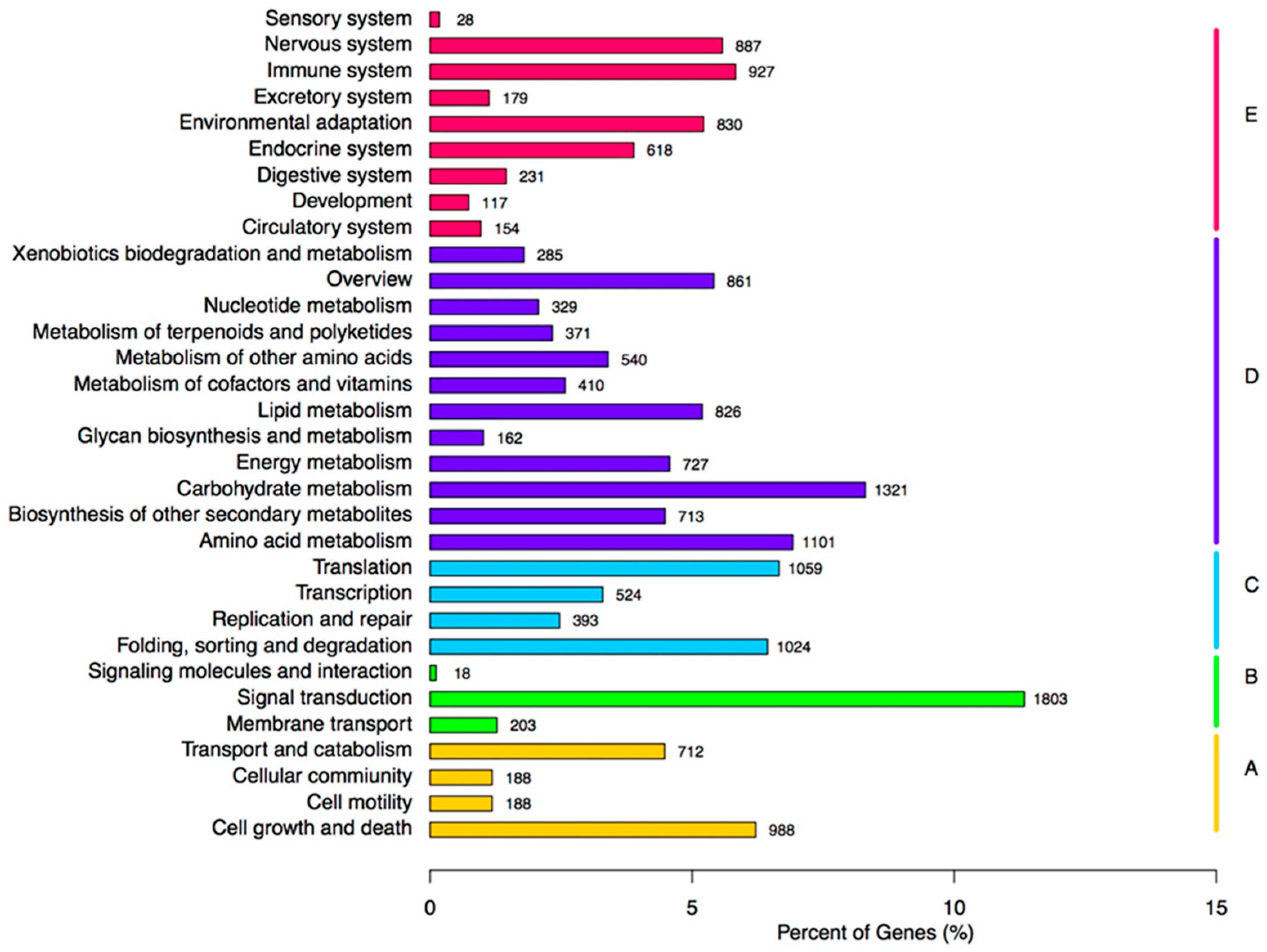
2.3. Functional Annotation of Assembled Unigenes
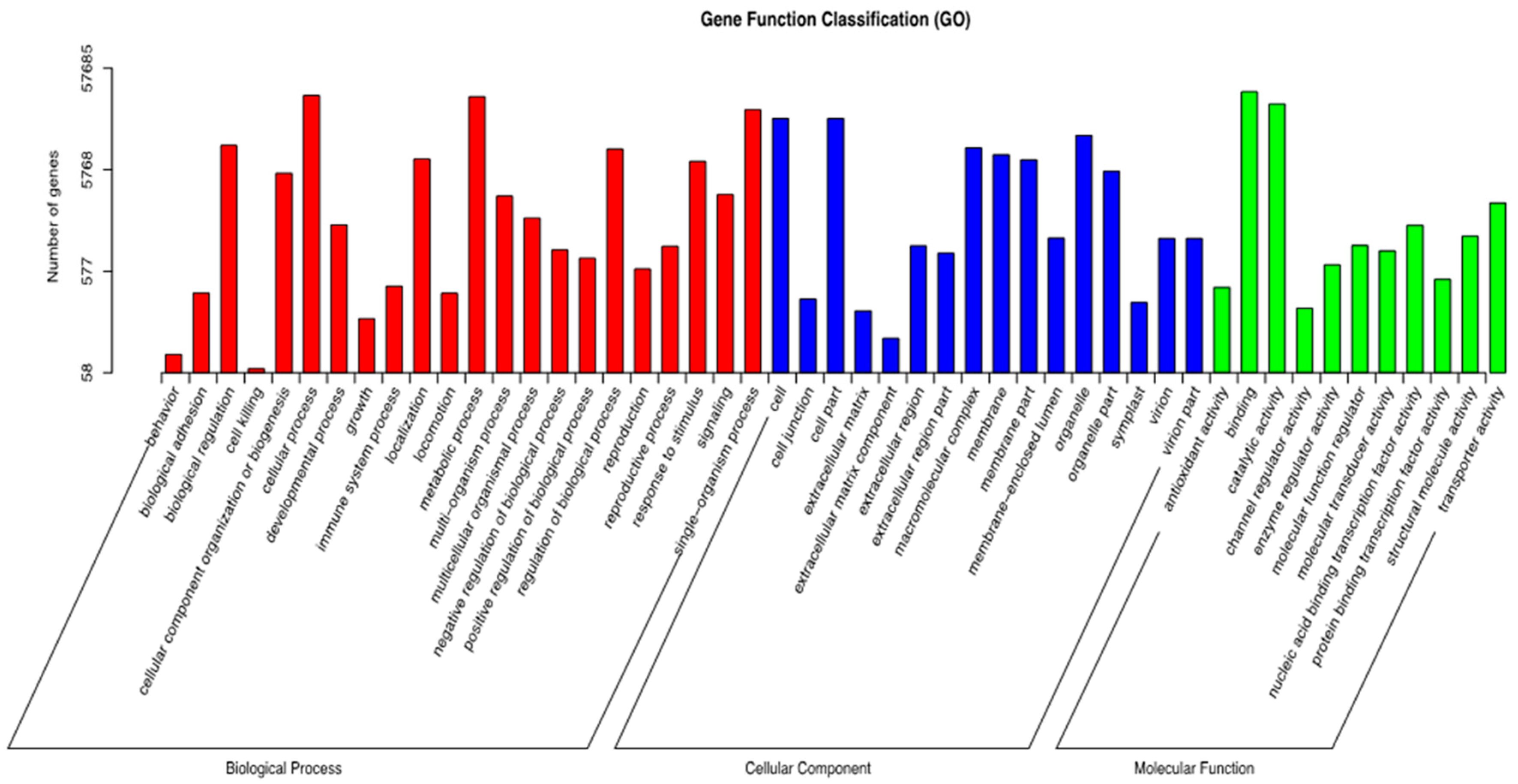
2.4. GO Classification
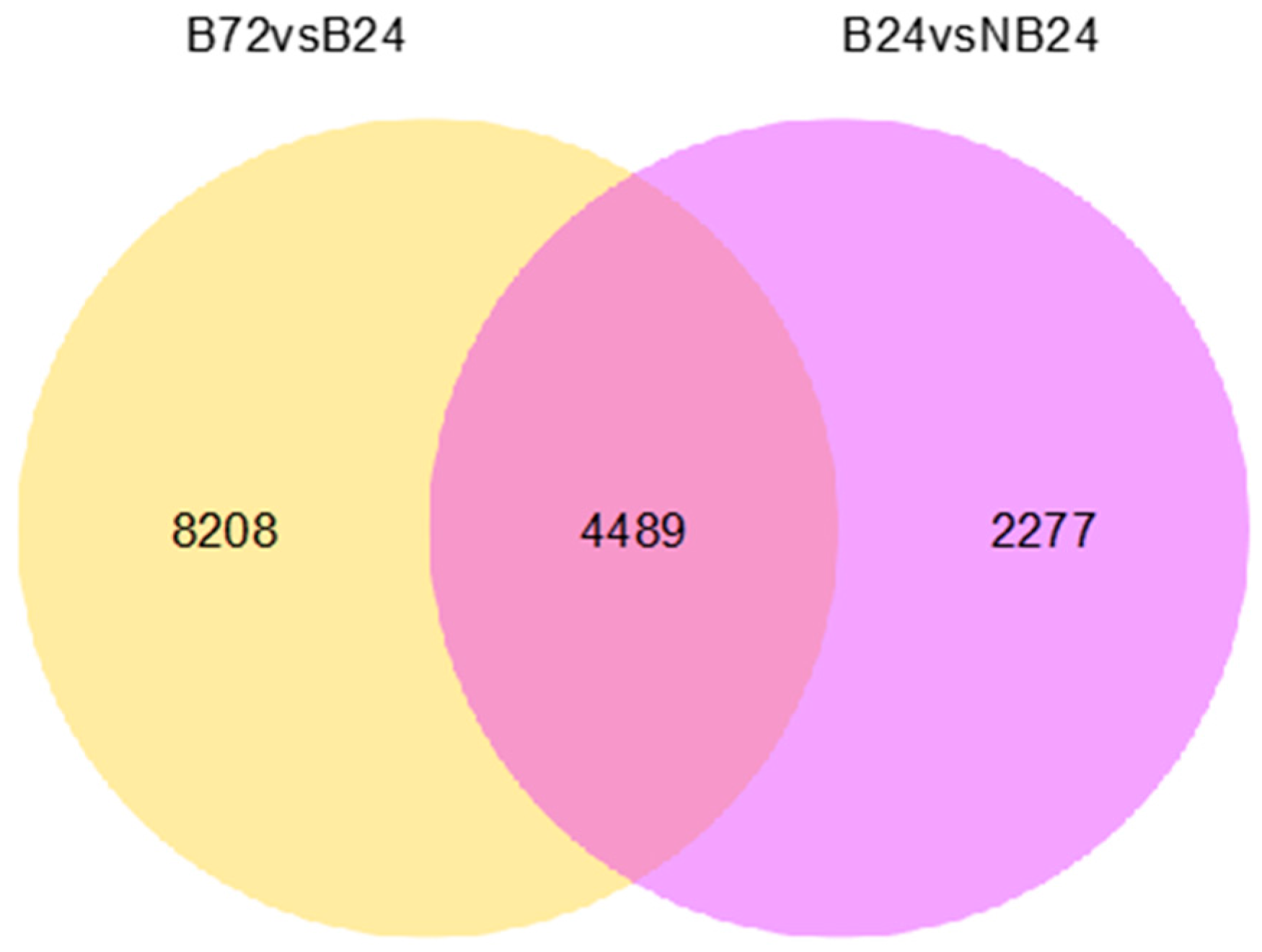
2.5. Differential Expression Analysis of Assembled Transcripts
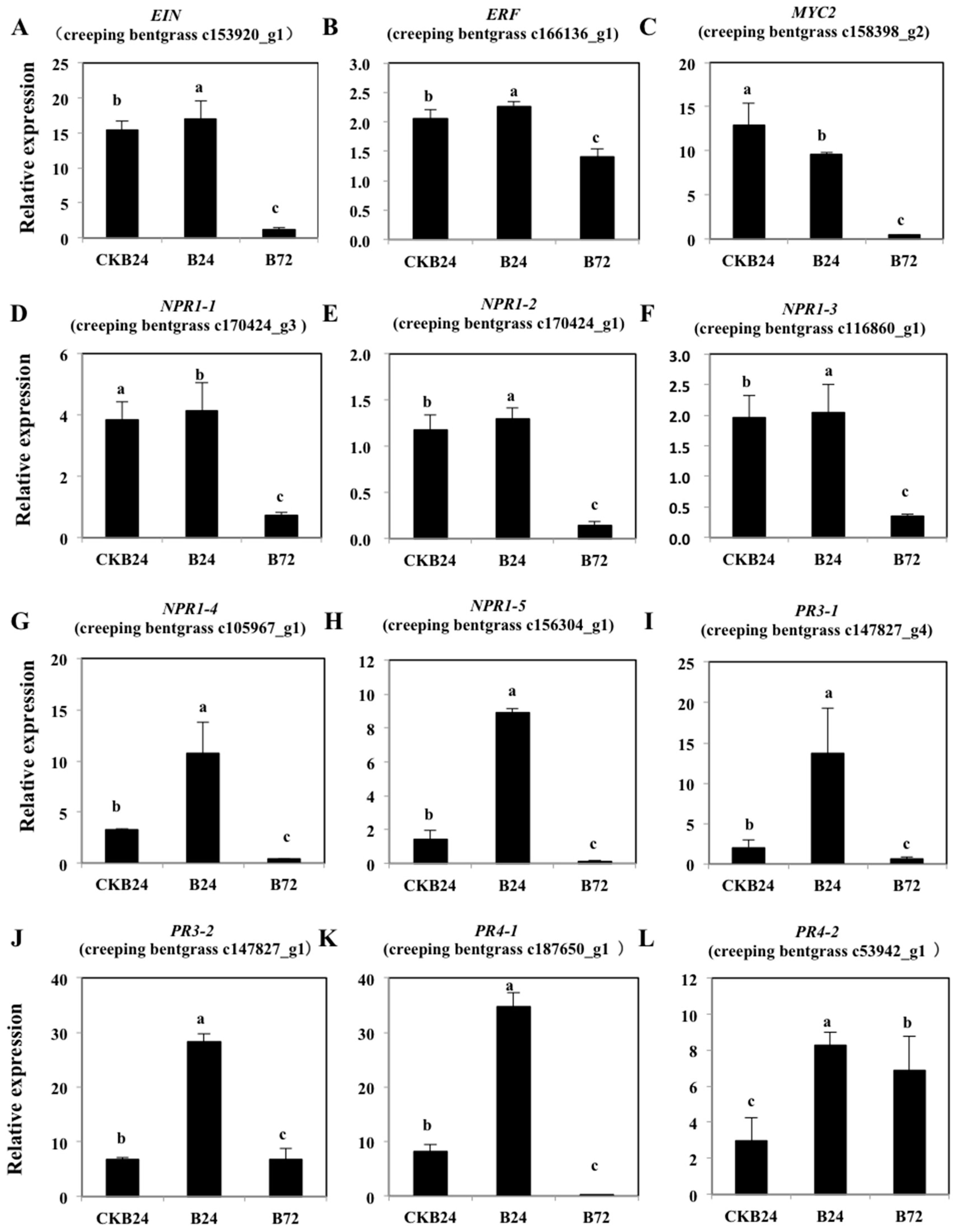
2.6. Expression Analysis of ET-Responsive and Defense-Related Genes in the BDO-Induced ISR Response
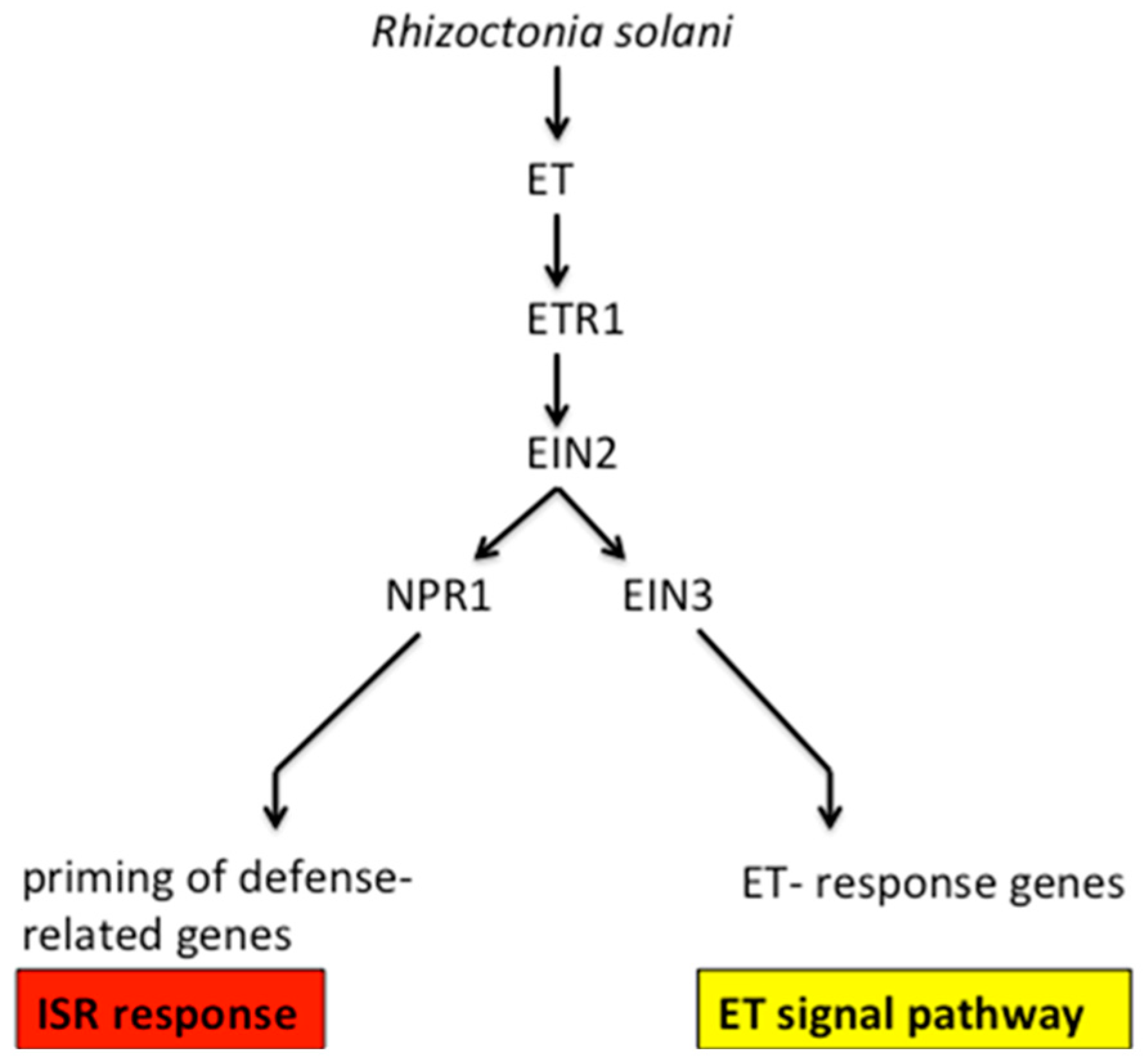
3. Discussion
4. Materials and Methods
4.1. Plant Growth Conditions and Treatments
4.2. Ethylene Measurement
4.3. 1-Aminocyclopropane-1-Carboxylic Acid Synthase (ACS) and 1-Aminocyclopropane-1-Carboxylic Acid Oxidase (ACO) Activity Assays
4.4. Total RNA, mRNA Isolation and Library Preparation for Transcriptome Sequencing
4.5. Library Preparation for Transcriptome Sequencing
4.6. Preprocessing and De Novo Assembly
4.7. Quantification of Gene Expression Levels and Differential Expression Analysis
4.8. Unigene Annotation and Classification
4.9. Quantitative Real-Time PCR Analysis
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by abscisic acid, salicylic acid and γ-aminobutyric acid in association with improved drought tolerance in creeping bentgrass (Agrostis stolonifera). Physiol. Plant. 2017, 159, 42–58. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, J.; Peng, Y.; Huang, B. Metabolic pathways regulated by γ-aminobutyric acid (GABA) contributing to heat tolerance in creeping bentgrass (Agrostis stolonifera). Sci. Rep. 2016, 6, 30338. [Google Scholar] [CrossRef] [PubMed]
- Merewitz, E.B.; Gianfagna, T.; Huang, B. Photosynthesis, water use, and root viability under water stress as affected by expression of SAG12-ipt controlling cytokinin synthesis in Agrostis stolonifera. J. Exp. Bot. 2011, 62, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.J.; Luo, T.Q. Control efficacy of 6 fungicides against 3 pathogens of turfgrass diseases. Acta Pratacul. Sin. 2013, 22, 124–131. [Google Scholar]
- Cortes-Barco, A.M.; Hsiang, T. Induced systemic resistance against three foliar diseases of Agrostis stolonifera by (2R,3R)-Butanediol or an isoparaffin mixture. Ann. Appl. Biol. 2010, 157, 179–189. [Google Scholar] [CrossRef]
- Ma, X.; Ma, H.L. Butanediol induced disease resistance against brown spot in creeping bentgrass. J. Gansu Agric. Univ. 2011, 46, 77–80. [Google Scholar]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.H.M.; Van peer, R.V. Suppression of soil-borne plant pathogens by fluorescent pseudomonads: Mechanisms and prospects. In Biotic Interactions and Soil-Borne Diseases; Beemster, A.B.R., Bollen, G.J., Eds.; Elsevier Scientific: Amsterdam, The Netherlands, 1991; pp. 217–230. [Google Scholar]
- Van Loon, L.C.; Bakker, P.A.H.M. Signalling in rhizobacteria-plant interactions. In Root Ecology; De Kroon, J., Visser, E.J.W., Eds.; Springer: Berlin, Germany, 2003; pp. 287–330. [Google Scholar]
- Van Loon, L.C.; Bakker, P.A.H.M. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, R.; Babu, S. A new bio-formulation containing plant growth promoting rhizobacterial mixture for the management of sheath blight and enhanced grain yield in rice. Biocontrol 2001, 46, 493–510. [Google Scholar] [CrossRef]
- Suzuki, S.; He, Y. Indole-3-acetic acid production in P. fluorescens HP72 and its asociation with suppression of creping bentgrass brown patch. Curr. Microbiol. 2003, 47, 138–143. [Google Scholar] [CrossRef]
- Johnson, P.R.; Ecker, J.R. The ethylene gas signal transduction pathway: A molecular perspective. Annu. Rev. Genet. 1998, 32, 227–254. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Shockey, J.A. The ethylene-response pathway: Signal perception to gene regulation. Curr. Opin. Plant Biol. 1999, 2, 352–358. [Google Scholar] [CrossRef]
- Bleecker, A.B.; Kende, H. Ethylene: A gaseous signal molecule in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.C.; Li, H. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, 131–151. [Google Scholar] [CrossRef]
- Solano, R.; Stepanova, A. Nuclear events in ethylene signaling: A transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE FACTOR1. Genes Dev. 1998, 12, 3703–3714. [Google Scholar] [CrossRef] [PubMed]
- Mandy, D.K.; Caren, C. Ethylene signaling: New levels of complexity and regulation. Curr. Opin. Plant Biol. 2008, 11, 479–485. [Google Scholar]
- Van Wees, S.C.M.; Luijendijk, M. Rhizobacteria-mediated induced systemic resistance (ISR) in Arabidopsis is not associated with a direct effect on expression of known defense-related genes but stimulates the expression of the jasmonate-inducible gene Atvsp upon challenge. Plant Mol. Biol. 1999, 41, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Vogel, J.P.; Garvin, D.F. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Abeles, F.B.; Morgan, P.W. Ethylene in Plant Biology, 2nd ed.; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Mizoguchi, T.; Ichimura, K. Environmental stress response in plants: The role of mitogen-activated protein kinases. Trends Biotechnol. 1997, 15, 15–19. [Google Scholar] [CrossRef]
- Zhang, S.; Klessig, D.F. MAPK cascades in plant defense signaling. Trends Plant Sci. 2001, 6, 520–527. [Google Scholar] [CrossRef]
- Boller, T. Ethylene in pathogenesis and disease resistance. In The Plant Hormone Ethylene; Mattoo, A.K., Suttle, J.C., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 293–314. [Google Scholar]
- Hoffman, R.; Roebroeck, E. Effects of ethylene biosynthesis in carrot root slices on 6-methoxymellein accumulation and resistance to Botrytis cinerea. Physiol. Plant 1988, 73, 71–76. [Google Scholar] [CrossRef]
- Alonso, J.; Hirayama, T. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 1999, 284, 2148–2152. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; van Wees, S.C. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 1998, 10, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Knoester, M.; Van Loon, L.C. Ethylene-insensitive tobacco lacks non-host resistance against soil-borne fungi. Proc. Natl. Acad. Sci. USA 1998, 95, 1933–1937. [Google Scholar] [CrossRef] [PubMed]
- Dong, X. SA, JA, ethylene, and disease resistance in plants. Curr. Opin. Plant Biol. 1998, 1, 316–323. [Google Scholar] [CrossRef]
- Achuo, E.A.; Audenaert, K. The salicylic acid-dependent defense pathway is effective against different pathogens in tomato and tobacco. Plant Pathol. 2004, 53, 65–72. [Google Scholar] [CrossRef]
- Ryals, J.; Weymann, K. The Arabidopsis NIM1 protein shows homology to the mammalian transcription factor inhibitor IkB. Plant Cell 1997, 9, 425–439. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J.; Rogers, E.E. Isolation of Arabidopsis mutants with enhanced disease susceptibility by direct screening. Genetics 1996, 143, 973–982. [Google Scholar] [PubMed]
- Schenk, P.M.; Kazan, K. Coordinated plant defense responses in Arabidopsis revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2000, 97, 11655–11660. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Eggermont, K. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed]
- Hoffland, E.; Pieterse, C.M.J. Induced systemic resistance in radish is not associated with accumulation of pathogenesis-related proteins. Physiol. Mol. Plant Pathol. 1995, 46, 309–320. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Wees, S.C.M. Systemic resistance in Arabidopsis induced by biocontrol bacteria is independent of salicylic acid accumulation and pathogenesis-related gene expression. Plant Cell 1996, 8, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Kroes, G.M.L.W.; Sommers, E. Two in vitro assays to evaluate resistance in Linum usitatissimum to Fusarium wilt disease. Eur. J. Plant Pathol. 1998, 104, 561–568. [Google Scholar] [CrossRef]
- Kim, C.Y.; Liu, Y. Activation of a stress-responsive mitogen-activated protein kinase cascade induces the biosynthesis of ethylene in plants. Plant Cell 2003, 15, 2707–2718. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Hayakawa, Y. Wound-induced ethylene synthesis and expression and formation of 1-aminocyclopropane-1-carboxylate (ACC) synthase, ACC oxidase, phenylalaninne ammonia lyase and peroxidase in wounded mesocarp tissue of Cucurbita maxima. Plant Cell Physiol. 2000, 41, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Moya-León, M.A.; John, P. Activity of 1-aminocyclopropane-1-carboxylate (ACC) oxidase (ethylene-forming enzyme) in the pulp and peel of ripening bananas. J. Hortic. Sci. 1994, 69, 243–250. [Google Scholar]
- Grabherr, M.G.; Haas, B.J. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Altschul, S.F.; Madden, T.L. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total raw reads | 954,985,454 |
| Total clean reads | 921,578,368 |
| GC content (%) | 54.59 |
| Q20 bases (%) | 97.41 |
| Total transcripts | 338,453 |
| Total length of transcripts (bp) | 227,642,178 |
| Transcript mean length (bp) | 673 |
| Transcripts with N50 length (bp) | 1024 |
| Total unigenes | 208,672 |
| Total length of unigenes (bp) | 116,222,950 |
| Unigenes mean length (bp) | 557 |
| Unigenes with N50 length (bp) | 775 |
| Number of Unigenes | Percentage (%) | |
|---|---|---|
| Annotated in NR | 71,824 | 34.41 |
| Annotated in NT | 46,867 | 22.45 |
| Annotated in KO | 15,903 | 7.62 |
| Annotated in SwissProt | 39,602 | 18.97 |
| Annotated in PFAM | 50,138 | 24.02 |
| Annotated in GO | 57,685 | 27.64 |
| Annotated in KOG | 16,393 | 7.85 |
| Annotated in all Databases | 6264 | 3 |
| Annotated in at least one Database | 90,004 | 43.13 |
| Total Unigenes | 208,672 | 100 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.-Y.; Zhang, J.-L.; Yang, J.-W.; Ma, H.-L. Transcript Profiling and Gene Identification Involved in the Ethylene Signal Transduction Pathways of Creeping Bentgrass (Agrostis stolonifera) during ISR Response Induced by Butanediol. Molecules 2018, 23, 706. https://doi.org/10.3390/molecules23030706
Jiang H-Y, Zhang J-L, Yang J-W, Ma H-L. Transcript Profiling and Gene Identification Involved in the Ethylene Signal Transduction Pathways of Creeping Bentgrass (Agrostis stolonifera) during ISR Response Induced by Butanediol. Molecules. 2018; 23(3):706. https://doi.org/10.3390/molecules23030706
Chicago/Turabian StyleJiang, Han-Yu, Jin-Lin Zhang, Jiang-Wei Yang, and Hui-Ling Ma. 2018. "Transcript Profiling and Gene Identification Involved in the Ethylene Signal Transduction Pathways of Creeping Bentgrass (Agrostis stolonifera) during ISR Response Induced by Butanediol" Molecules 23, no. 3: 706. https://doi.org/10.3390/molecules23030706
APA StyleJiang, H.-Y., Zhang, J.-L., Yang, J.-W., & Ma, H.-L. (2018). Transcript Profiling and Gene Identification Involved in the Ethylene Signal Transduction Pathways of Creeping Bentgrass (Agrostis stolonifera) during ISR Response Induced by Butanediol. Molecules, 23(3), 706. https://doi.org/10.3390/molecules23030706