Formation of Sulforaphane and Iberin Products from Thai Cabbage Fermented by Myrosinase-Positive Bacteria

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. GSL-Metabolizing Bacteria from Thai Fermented Foods and Drinks
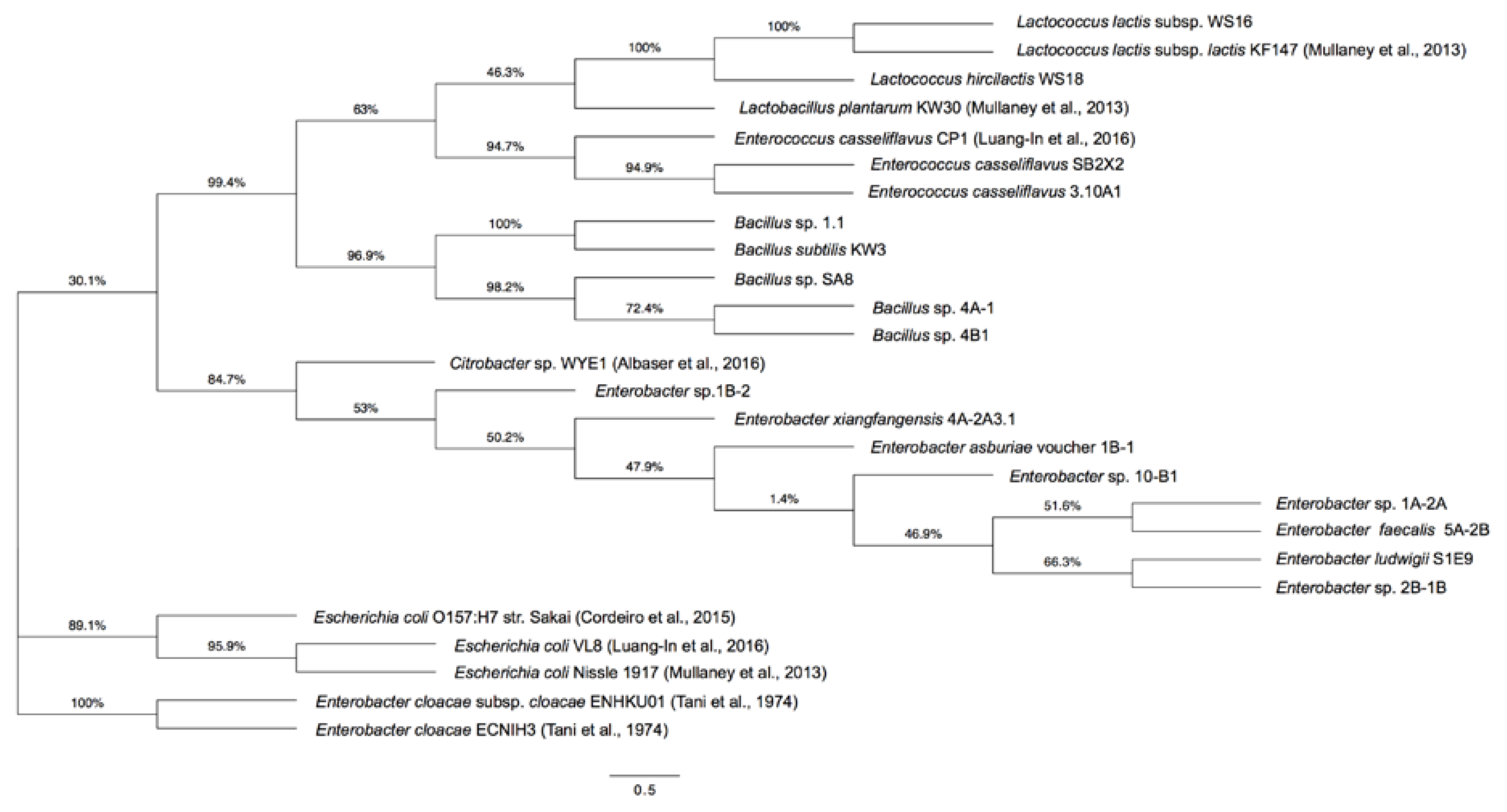
2.2. Phylogenetic Tree of GSL-Metabolizing Bacteria
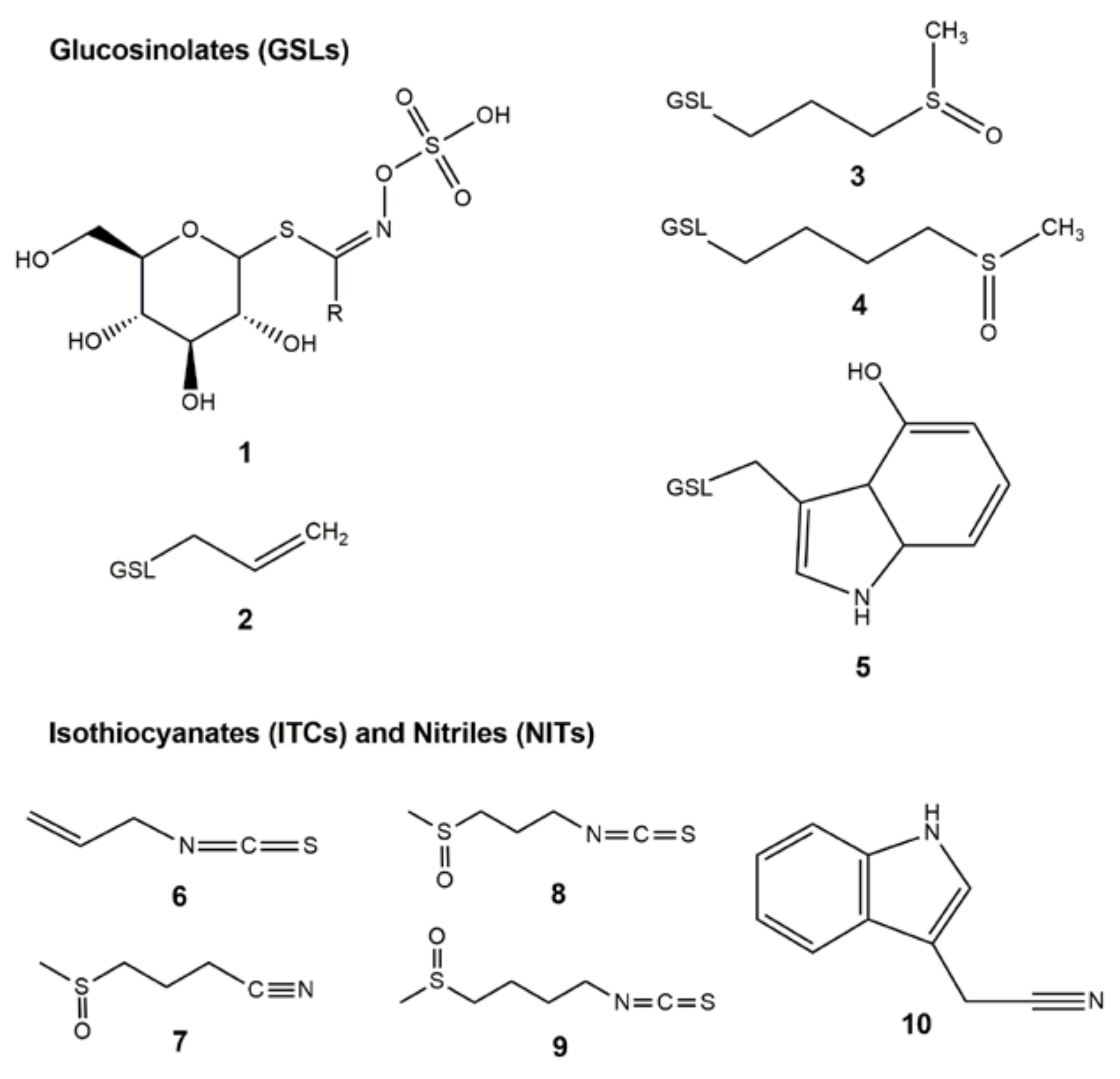
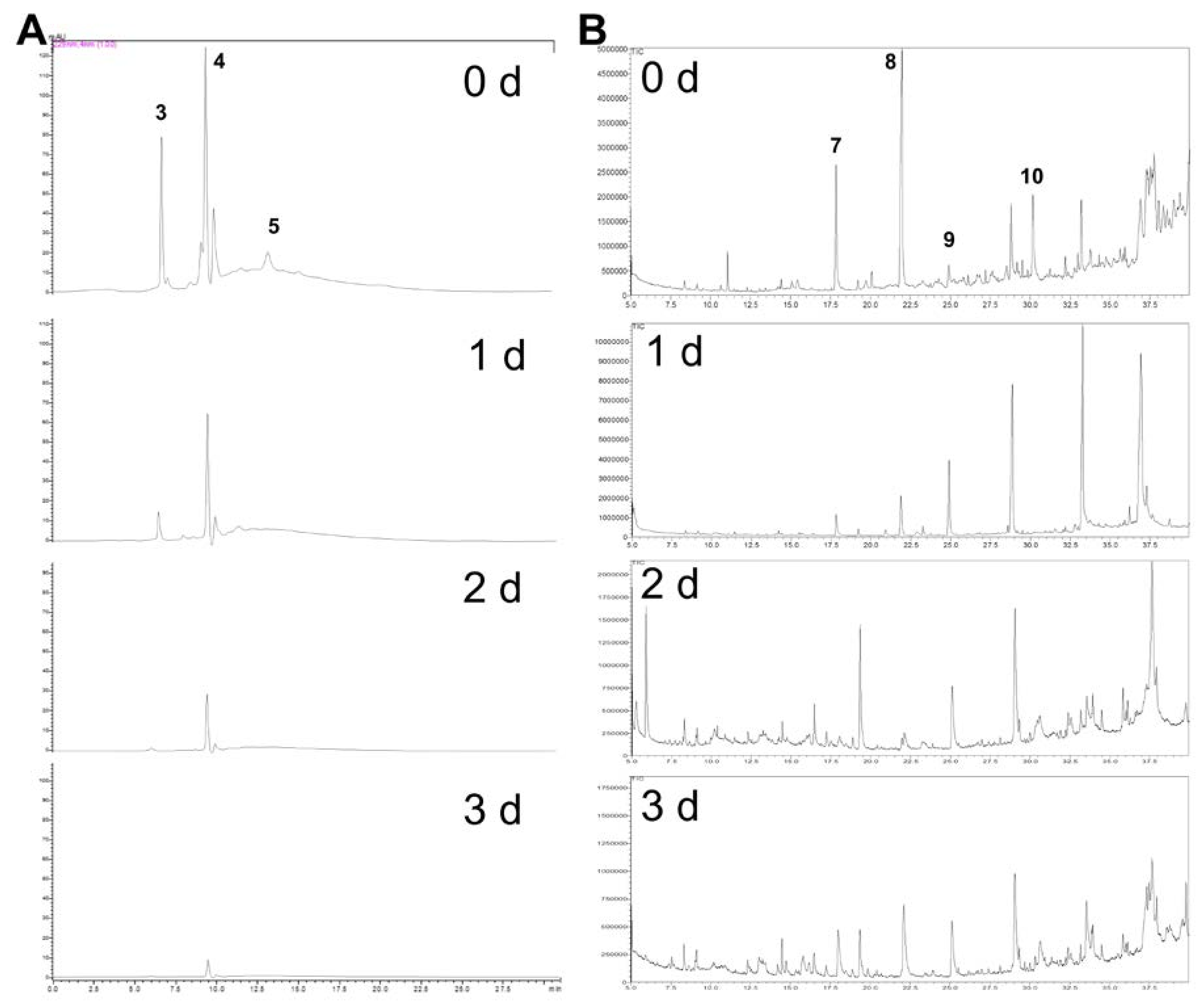
2.3. ITC Products from Fermented Cabbage
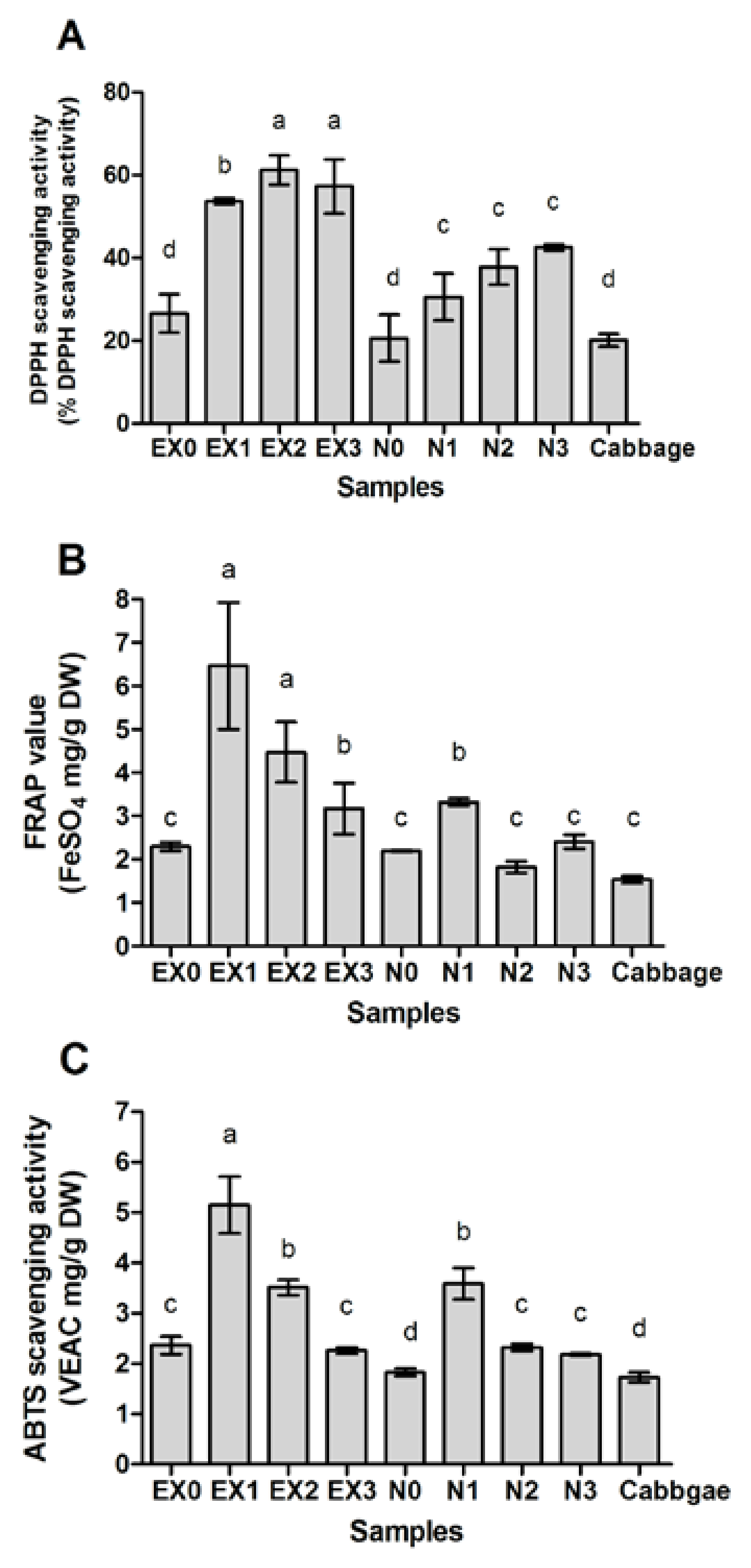
2.4. Antioxidant Activity of Fermented Cabbage
3. Materials and Methods
3.1. Sample Collection
3.2. Isolation of GSL-Metabolizing Microbes
3.3. In Vitro Sinigrin Incubation
3.4. Genomic DNA Isolation and 16S rRNA Gene Analysis
3.5. Starter Culture Preparation
3.6. Cabbage Fermentations
3.7. Sampling and Extraction of Fermented Cabbage
3.8. Sample Preparation and HPLC Analysis to Detect GSLs
3.9. Sample Preparation and GC-MS Analysis to Detect Degradation Products
3.10. Antioxidant Activity of Fermented Cabbage
3.11. Statistical Analyses
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kaufman, N.D.; Chasombat, S.; Tanomsingh, S.; Rajataramya, B.; Potempa, K. Public health in Thailand: Emerging focus on non-communicable diseases. Int. J. Health Plan. Manag. 2011, 26, e197–e212. [Google Scholar] [CrossRef] [PubMed]
- Xanthos, D. Country in focus: Economic transition and non-communicable diseases in Thailand. Lancet Diabetes Endocrinol. 2015, 3, 684–685. [Google Scholar] [CrossRef]
- Country Statistics and Global Health Estimates by WHO and UN Partners. Available online: http://www.who.int/gho/en/ (accessed on 31 March 2018).
- Camps, J.; García-Heredia, A. Introduction: Oxidation and inflammation, a molecular link between non-communicable diseases. Adv. Exp. Med. Biol. 2014, 824, 1–4. [Google Scholar] [PubMed]
- Sun, Y.P.; Chou, C.C.; Yu, R.C. Antioxidant activity of lactic-fermented Chinese cabbage. Food Chem. 2009, 115, 912–917. [Google Scholar] [CrossRef]
- Park, J.M.; Shin, J.H.; Gu, J.G.; Yoon, S.J.; Song, J.C.; Jeon, W.M.; Suh, H.J.; Chang, U.J.; Yang, C.Y.; Kim, J.M. Effect of antioxidant activity in kimchi during a short-term and over-ripening fermentation period. J. Biosci. Bioeng. 2011, 122, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shao, Y. Effects of microbial diversity on nitrite concentration in pao cai, a naturally fermented cabbage product from China. Food Microbiol. 2018, 72, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Parveen Rani, R. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014, 2014, 250424. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Das, G.; Paramithiotis, S.; Shin, H.S. Kimchi and other widely consumed traditional fermented foods of Korea: A review. Front. Microbiol. 2016, 7, 1493. [Google Scholar] [CrossRef] [PubMed]
- Beganović, J.; Kos, B.; Pavunc, A.L.; Uroić, K.; Jokić, M.; Šušković, J. Traditionally produced sauerkraut as source of autochthonous functional starter cultures. Microbiol. Res. 2014, 169, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Verkerk, R.; Schreiner, M.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; De Schrijver, R.; Hansen, M.; Gerhäuser, C.; Mithen, R.; et al. Glucosinolates in Brassica vegetables: The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, S219. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Kelleher, M.O.; Eggleston, I.M. The cancer chemopreventive actions of phytochemicals derived from glucosinolates. Eur. J. Nutr. 2008, 47, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Kusznierewicz, B.; Śmiechowska, A.; Bartoszek, J.; Namieśnik, J. The effect of heating and fermenting on antioxidant properties of white cabbage. Food Chem. 2008, 108, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.L.; Hashimoto, K.; Uda, Y. In vitro digestion of sinigrin and glucotropaeolin by single strains of Bifidobacterium and identification of the digestive products. Food Chem. Toxicol. 2004, 42, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Luang-In, V.; Albaser, A.A.; Nueno-Palop, C.; Narbad, A.; Bennett, M.; Rossiter, J.R. Metabolism of glucosinolates and desulfo-glucosinolates by selected human gut bacteria. Curr. Microbiol. 2016, 73, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Albaser, A.; Kazana, E.; Bennett, M.; Cebeci, F.; Luang-In, V.; Spanu, P.D.; Rossiter, J.T. Discovery of a bacterial glycoside hydrolase family 3 (GH3) β-glucosidase with myrosinase activity from a Citrobacter strain isolated from soil. J. Agric. Food Chem. 2016, 64, 1520–1527. [Google Scholar] [CrossRef] [PubMed]
- Luciano, F.B.; Holley, R.A. Bacterial degradation of glucosinolates and its influence on the growth of E. coli O157:H7. Fleischwirtsch. Int. 2011, 1, 78–81. [Google Scholar]
- Tani, N.; Ohtsuru, M.; Hata, T. Isolation of myrosinase producing microorganism. Agric. Biol. Chem. 1974, 38, 1617–1622. [Google Scholar] [CrossRef]
- Luang-In, V.; Rossiter, J.T. Stability studies of isothiocyanates and nitriles in aqueous media. Songklanakarin J. Sci. Technol. 2015, 37, 625–630. [Google Scholar]
- El-Shora, H.M.; El-Shobaky, A.M.; El-Atrozy, M.M. Activity of purified bacterial myrosinase and its essential residues. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 567–578. [Google Scholar] [CrossRef]
- Madhuri, R.J.; Amerada, M. Production of myrosinase enzyme by Actinomycetes isolated from cotton soil. Int. J. Chem. Environ. Biol. Sci. 2015, 3, 397–403. [Google Scholar]
- Luang-In, V.; Narbad, A.; Nueno-Palop, C.; Mithen, R.; Bennett, M.; Rossiter, J.T. The metabolism of methylsulfinylalkyl- and methylthioalkyl-glucosinolates by a selection of human gut bacteria. Mol. Nutr. Food Res. 2014, 58, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Mullaney, J.A.; Kelly, W.; Mcghie, T.K.; Ansell, J.; Heyes, J.A. Lactic acid bacteria convert glucosinolates to nitriles efficiently yet differently to Enterobacteriaceae. J. Agric. Food Chem. 2013, 61, 3039–3046. [Google Scholar] [CrossRef] [PubMed]
- Llanos-Palop, M.; Smiths, J.P.; Brink, B.T. Degradation of sinigrin by Lactobacillus agilis strain R16. Int. J. Food Microbiol. 1995, 26, 219–229. [Google Scholar] [CrossRef]
- Gu, C.T.; Li, C.Y.; Yang, L.J.; Huo, C.G. Enterobacter xiangfangensis sp. nov., isolated from Chinese traditional sourdough, and reclassification of Enterobacter sacchari Zhu et al. 2013 as Kosakonia sacchari comb. Nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 2650–2656. [Google Scholar] [CrossRef] [PubMed]
- Paramithiotis, S.; Hondrodimou, O.L.; Drosinos, E.H. Development of the microbial community during spontaneous cauliflower fermentation. Food Res. Int. 2013, 43, 1098–1103. [Google Scholar] [CrossRef]
- Cordeiro, R.P.; Doria, J.H.; Zhanel, G.G.; Sparling, R.; Holley, R.A. Role of glycoside hydrolase genes in sinigrin degradation by E. coli O157:H7. Int. J. Food Microbiol. 2015, 205, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, M.J.; Jeong, M.H.; Kim, J.E. Identification and quantification of glucosinolates in kimchi by liquid chromatography-electrospray tandem mass spectrometry. Int. J. Anal. Chem. 2017, 2017, 6753481. [Google Scholar] [CrossRef] [PubMed]
- Bohinc, T.; Trdan, S. Environmental factors affecting the glucosinolate content in Brassicaceae. J. Food Agric. Environ. 2012, 10, 357–360. [Google Scholar]
- Peñas, E.; Pihlava, J.M.; Vidal-Valverde, C.; Frias, J. Influence of fermentation conditions of Brassica oleracea L. var. capitata on the volatile glucosinolate hydrolysis compounds of sauerkrauts. LWT Food Sci. Technol. 2012, 48, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Tolonen, M.; Taipale, M.; Viander, B.; Pihlava, J.M.; Korhonen, H.; Ryhänen, E.L. Plant-derived biomolecules in fermented cabbage. J. Agric. Food Chem. 2002, 50, 6798–6803. [Google Scholar] [CrossRef] [PubMed]
- Palani, K.; Harbaum-Piayda, B.; Meske, D.; Keppler, J.K.; Bockelmann, W.; Heller, K.J.; Schwarz, K. Influence of fermentation on glucosinolates and glucobrassicin degradation products in sauerkraut. Food Chem. 2016, 190, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.R.; Lee, K.J.; Kim, H.Y.; Kim, J.H.; Kim, Y.B.; Sok, D.E. Effect of various kimchi extracts on the hepatic glutathione S-transferase activiy of mice. Food Res. Int. 1999, 31, 389–394. [Google Scholar] [CrossRef]
- Hong, E.; Kim, G.H. GC-MS analysis of the extracts from Korean cabbage (Brassica campestris L. ssp. pekinensis) and its seed. Prev. Nutr. Food Sci. 2013, 18, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Min, J.H.; Lee, J.H.; Ji, G.E. Growth of lactic acid bacteria and bifidobacteria in natural media using vegetables, seaweeds, grains and potatoes. Food Sci. Biotechnol. 2000, 9, 322–324. [Google Scholar]
- Singh, P.; Saini, P.; Puranik, V.; Gupta, S.; Dubey, S. Optimization and characterization of exopolysaccharides produced by lactobacillus strains isolated from cabbage and cucumber. J. Microbiol. Biotechnol. Res. 2016, 6, 27–35. [Google Scholar]
- Bhat, B.; Bajaj, B.K. Hypocholesterolemic and bioactive potential of exopolysaccharide from a probiotic Enterococcus faecium K1 isolated from kalarei. Bioresour. Technol. 2018, 254, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, K.; Devasya, R.P.; Bhagwath, A.A. Exopolysaccharide produced by Enterobacter sp. YG4 reduced uranium induced nephrotoxicity. Int. J. Biol. Macromol. 2016, 82, 557–561. [Google Scholar]
- Sakanaka, S.; Tachibana, Y.; Okada, Y. Preparation and antioxidant properties of extracts of Japanese persimmon leaf tea (kakinoha-cha). Food Chem. 2005, 89, 569–575. [Google Scholar] [CrossRef]
- Hunaefi, D.; Gruda, N.; Riedel, H.; Akumo, N.D.; Thaw Saw, N.M.M.; Smetanska, I. Improvement of antioxidant activities in red cabbage sprouts by lactic acid bacterial fermentation. Food Biotechnol. 2013, 27, 279–302. [Google Scholar] [CrossRef]
- Vicaş, S.; Rugină, D.; Leopold, L.; Pintea, A.; Socaciu, C. HPLC fingerprint of bioactive compounds and antioxidant activities of Viscum albumfrom different host trees. Notulae Botanicae 2011, 39, 48–57. [Google Scholar]
- Akowuah, G.A.; Ismail, Z.; Norhayati, I.; Sadikun, A. The effects of different extraction solvents of varying polarities of polyphenols of Orthosiphon stamineus and evaluation of the free radical-scavenging activity. Food Chem. 2005, 93, 311–317. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Adams, L.S.; Zhang, Y.; Lee, R.; Sand, D.; Scheuller, H.S.; Heber, D. Blackberry, black raspberry, blueberry, cranberry, red raspberry, and strawberry extracts inhibit growth and stimulate apoptosis of human cancer cells in vitro. J. Agric. Food Chem. 2006, 54, 9329–9339. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of compounds 1–10 are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession no. a | Species | Closest Relative Species b (% Identity)/Accession no. c/Origin of Isolate d | Sinigrin Degradation (nmol) | AITC Product (nmol) | % Product Formation e |
|---|---|---|---|---|---|---|
| 1. Fermented cabbage pH 3.87 | ||||||
| 1 | LC342980.1 | Enterobacter sp. 1A-1A | Enterobacter sp. Md1-53 (92%) MF581459.1 Paeonia ostii root, China | 73 ± 8 | 30 ± 5 | 41 ± 4 |
| 2 | LC342981.1 | Enterobacter faecalis 5A-2B | Enterococcus faecalis NW A20 (99%) MG543833.1 Raw meat, South Africa | 78 ± 7 | 39 ± 11 | 50 ± 14 |
| 3 | LC342982.1 | Enterobacter asburiae 1B-1 | Enterobacter asburiae voucher ST56 (100%) KT287073.1 Rumen, China | 75 ± 11 | 33 ± 8 | 44 ± 6 |
| 4 | LC342983.1 | Enterobacter sp. 1B-2 | Enterobacter sp. NU33 (96%) MG459258.1 Plant growth-promoting bacteria in sugarcane, Brazil | 79 ± 5 | 41 ± 7 | 52 ± 8 |
| 5 | LC342984.1 | Enterobacter ludwigii S1E9 | Enterobacter ludwigii HTP04 (100%) KX024731.1 Shrimp gut, India | 90 ± 8 | 50 ± 9 | 56 ± 7 |
| 6 | LC342985.1 | Enterococcus casseliflavus SB2X2 | Enterococcus casseliflavus HMF4406 (98%) KT984002.1 Jeotgal (salted fermented food), Korea | 100 ± 0 | 61 ± 4 | 61 ± 6 |
| 7 | LC342986.1 | Bacillus sp. SA8 | Bacillus sp. SK123 (97%) KU060226.1 Honey bee apiary, Thailand | 79 ± 8 | 39 ± 7 | 49 ± 6 |
| 8 | LC342987.1 | Bacillus sp. 1.1 | Bacillus sp. BDU13 (96%) JX847614.1 Fermented fish, India | 87 ± 10 | 42 ± 11 | 48 ± 10 |
| 2. Pickled onion pH 4.81 | ||||||
| 9 | LC342980.1 | Enterobacter sp. 1A-1A | Enterobacter sp. Md1-53 (92%) MF581459.1 Paeonia ostii root, China | 73 ± 5 | 40 ± 5 | 55 ± 4 |
| 10 | LC342988.1 | Enterobacter sp. 2B-1B | Enterobacter sp. SR19 (100%) KF896099.1 Seawater sediment, Belgium | 71 ± 0 | 39 ± 3 | 55 ± 6 |
| 3. Fermented fish pH 4.60 | ||||||
| 11 | LC342989.1 | Enterobacter xiangfangensis 4A-2A3.1 | Enterobacter xiangfangensis W31 (100%) KP813789.1 Storm water bacteria in two urban lakes, China | 100 ± 0 | 65 ± 3 | 65 ± 4 |
| 12 | LC342990.1 | Bacillus sp. 4A-1 | Bacillus sp. S42 (100%) JX293317.1 Crystal tuff, China | 71 ± 6 | 40 ± 4 | 56 ± 1 |
| 13 | LC342991.1 | Bacillus sp. 4B1 | Bacillus sp. SO5.17 (97%) KC867296.1 Mine drainage, Brazil | 73 ± 4 | 42 ± 9 | 58 ± 13 |
| 4. Fermented pork pH 4.73 | ||||||
| 14 | LC342992.1 | Enterococcus casseliflavus 3.10A1 | Enterococcus casseliflavus JFL12 (100%) KT343156.1 Fiber-degrading bacteria in rumen of Tibetan yak, China | 74 ± 13 | 35 ± 8 | 47 ± 4 |
| 5. Fermented herbal drink pH 2.80 | ||||||
| 15 | LC342981.1 | Enterobacter faecalis 5A-2B | Enterococcus faecalis NW A20 (99%) MG543833.1 Raw meat, South Africa | 78 ± 7 | 35 ± 6 | 45 ± 5 |
| 6. Fermented juice pH 2.93 | ||||||
| 16 | LC342993.1 | Enterobacter sp. 10-B1 | Enterobacter sp. DBM3 (97%) KT957440.1 Plutella xylostella larval gut, China | 77 ± 5 | 34 ± 11 | 42 ± 16 |
| 17 | LC342980.1 | Enterobacter sp. 1A-1A | Enterobacter sp. Md1-53 (92%) MF581459.1 Paeonia ostii root, China | 73 ± 10 | 34 ± 8 | 47 ± 7 |
| 7. Water kefir from Nakhon Ratchasima pH 5.94 | ||||||
| 18 | LC336444.1 | Lactococcus hircilactis WS16 | Lactococcus hircilactis DSM 28960 (100%) KJ201026.1 Goat milk, Italy | 77 ± 4 | Nd | na |
| 19 | LC336446.1 | Lactococcus lactis WS18 | Lactococcus lactis RCB787 (98%) KT260999.1 Bat guano, India | 78 ± 3 | Nd | na |
| 8. Milk kefir from Kamphaeng Phet pH 5.23 | ||||||
| 20 | LC342994.1 | Bacillus subtilis KW3 | Bacillus subtilis MA-48 (93%) KX426648.1 Rhizospheric soil in desert, Pakistan | 80 ± 9 | nd | na |
| Samples | Remaining GSLs (µmol/100 g Dry Weight) | Products (µmol/100 g Dry Weight) | |||||
|---|---|---|---|---|---|---|---|
| GIB | GRP | GBS | IBN | IBR | SFN | IAN | |
| Cabbage | 430.5 ± 34.1aB | 615.1 ± 30.0aA | 108.5 ± 19.1aC | 75.1 ± 26.4aC | 13.3 ± 10.0bD | 39.5 ± 16.3dC | 49.1 ± 29.8abC |
| N0 | 273.8 ± 15.8bB | 534.4 ± 30.4abA | 30.2 ± 4.4bC | 61.82 ± 28.4abC | 11.2 ± 6.0bD | 56.2 ± 26.0dC | 52.4 ± 19.1abC |
| N1 | 103.8 ± 19.5cB | 419.0 ± 26.5cA | 0.0 + 0.0cE | 43.3 ± 23.2abC | 32.1 ± 3.9bD | 135.2 ± 42.0abB | 22.9 ± 4.7abC |
| N2 | 20.9 ± 12.0dB | 217.5 ± 14.8deA | 0.0 + 0.0cC | 35.2 ± 15.4abB | 11.57 ± 5.8bD | 127.9 ± 60.0abA | 15.8 ± 2.0bB |
| N3 | 2.9 ± 1.1dC | 146.3 ± 27.7deA | 0.0 + 0.0cD | 21.6 ± 11.0bcB | 8.6 ± 3.9bB | 112.2 ± 52.0abA | 10.7 ± 3.1bB |
| EC0 | 305.7 ± 29.8bB | 514.2 ± 42.6bA | 27.4 ± 6.9bD | 64.3 ± 23.3abC | 15.4 ± 5.9bD | 45.6 ± 22.0dC | 60.5 ± 21.8aC |
| EC1 | 91.8 ± 17.3cC | 359.9 ± 31.8cdA | 0.0 + 0.0cE | 25.6 ± 12.6abD | 35.2 ± 20.0bD | 177.8 ± 42.0abB | 22.5 ± 14.6abD |
| EC2 | 21.0 ± 9.3dB | 211.0 ± 27.7deA | 0.0 + 0.0cD | 6.4 ± 2.1cC | 51.7 ± 35.9abB | 242.6 ± 40.0aA | 14.7 ± 11.9abB |
| EC3 | 3.1 ± 0.7dD | 111 ± 20.2fB | 0.0 + 0.0cE | 2.9 ± 0.8cD | 33.6 ± 22.0bC | 222.4 ± 42.0aA | 16.4 ± 7.2abC |
| EX0 | 305.2 ± 13.0bB | 536.6 ± 29.1abA | 31.6 ± 7.8aC | 65.2 ± 20.2abC | 12.5 ± 4.0bD | 48.6 ± 30.0dC | 55.3 ± 20.2abC |
| EX1 | 74.3 ± 14.6cC | 275.9 ± 19.deA | 0.0 + 0.0cF | 26.6 ± 12.3abD | 117.4 ± 41.9aB | 294.1 ± 44.0aA | 7.7 ± 3.7bE |
| EX2 | 11.1 ± 6.4dD | 160.8 ± 29.0eB | 0.0 + 0.0cE | 6.5 ± 3.8cD | 78.2 ± 38.0abC | 252.6 ± 46.0aA | 16.3 ± 7.4abD |
| EX3 | 2.4 ± 0.6dD | 85.3 ± 24.7fB | 0.0 + 0.0cE | 13.8 ± 3.1bcC | 70.5 ± 39.9abB | 244.7 ± 70.0aA | 20.9 ± 8.8abC |
| EC2 solid | 5.2 ± 1.6dC | 93.3 ± 23.6fA | 0.0 + 0.0cD | 3.6 ± 2.3cC | 19.2 ± 10.0bB | 69.7 ± 34.0cA | 5.9 ± 1.2bB |
| EC2 liquid | 16.3 ± 7.5dC | 117.7 ± 34.4fB | 0.0 + 0.0cF | 3.5 ± 0.7cE | 30.8 ± 22.0bC | 173.3 ± 48.0abA | 10.4 ± 1.3bD |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luang-In, V.; Deeseenthum, S.; Udomwong, P.; Saengha, W.; Gregori, M. Formation of Sulforaphane and Iberin Products from Thai Cabbage Fermented by Myrosinase-Positive Bacteria. Molecules 2018, 23, 955. https://doi.org/10.3390/molecules23040955
Luang-In V, Deeseenthum S, Udomwong P, Saengha W, Gregori M. Formation of Sulforaphane and Iberin Products from Thai Cabbage Fermented by Myrosinase-Positive Bacteria. Molecules. 2018; 23(4):955. https://doi.org/10.3390/molecules23040955
Chicago/Turabian StyleLuang-In, Vijitra, Sirirat Deeseenthum, Piyachat Udomwong, Worachot Saengha, and Matteo Gregori. 2018. "Formation of Sulforaphane and Iberin Products from Thai Cabbage Fermented by Myrosinase-Positive Bacteria" Molecules 23, no. 4: 955. https://doi.org/10.3390/molecules23040955
APA StyleLuang-In, V., Deeseenthum, S., Udomwong, P., Saengha, W., & Gregori, M. (2018). Formation of Sulforaphane and Iberin Products from Thai Cabbage Fermented by Myrosinase-Positive Bacteria. Molecules, 23(4), 955. https://doi.org/10.3390/molecules23040955