An Overview of Biotransformation and Toxicity of Diterpenes


Abstract
:1. Introduction
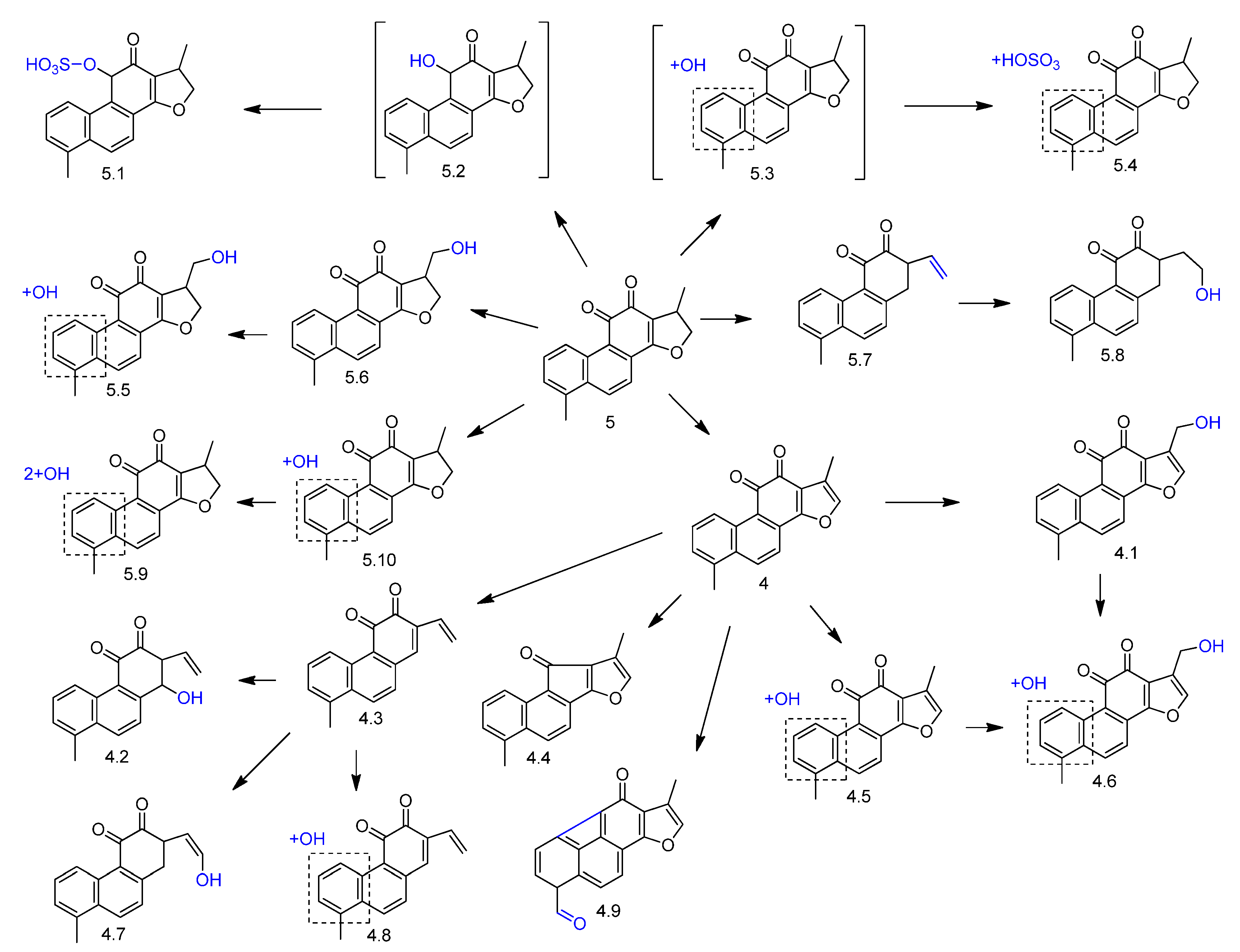
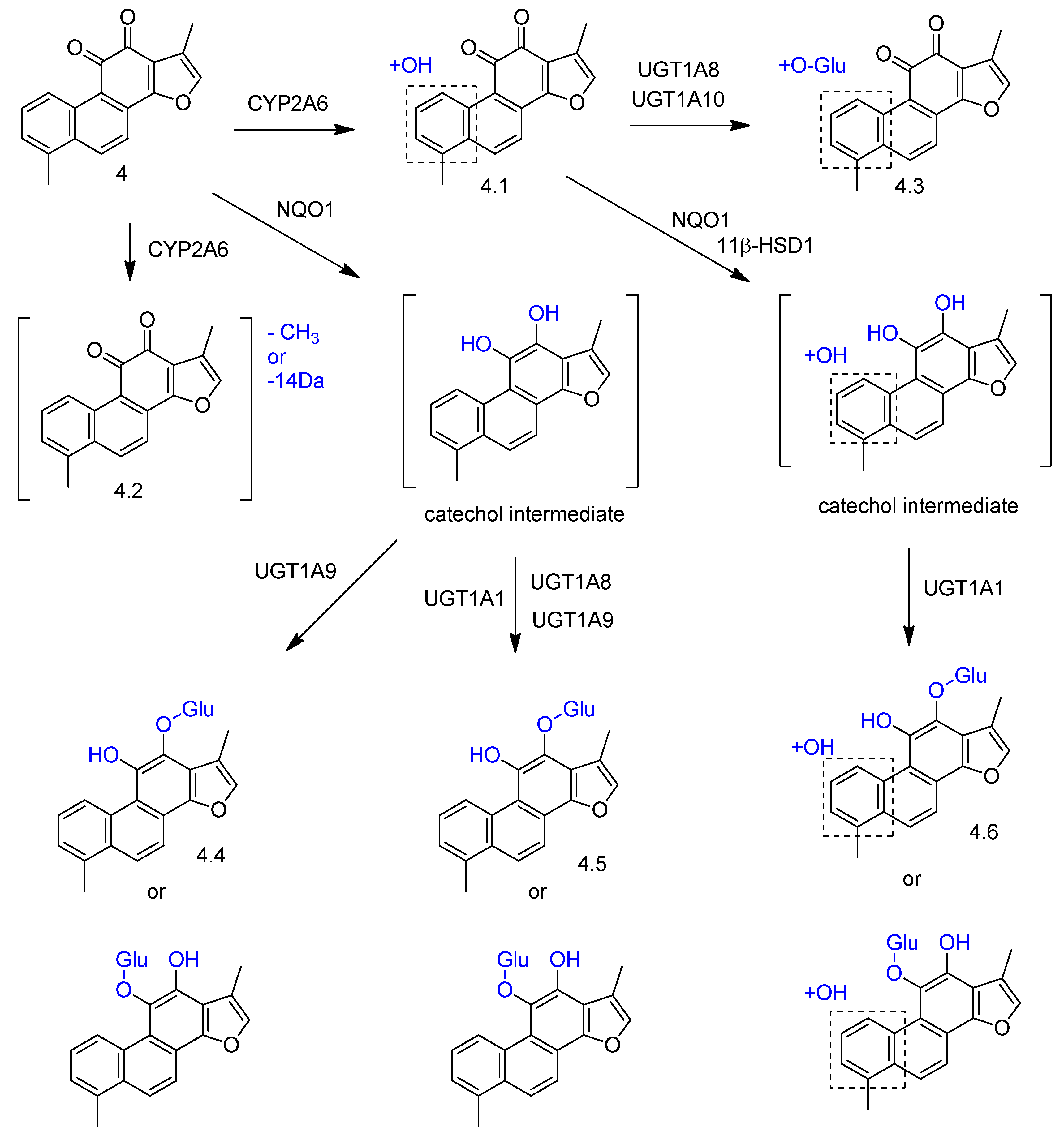
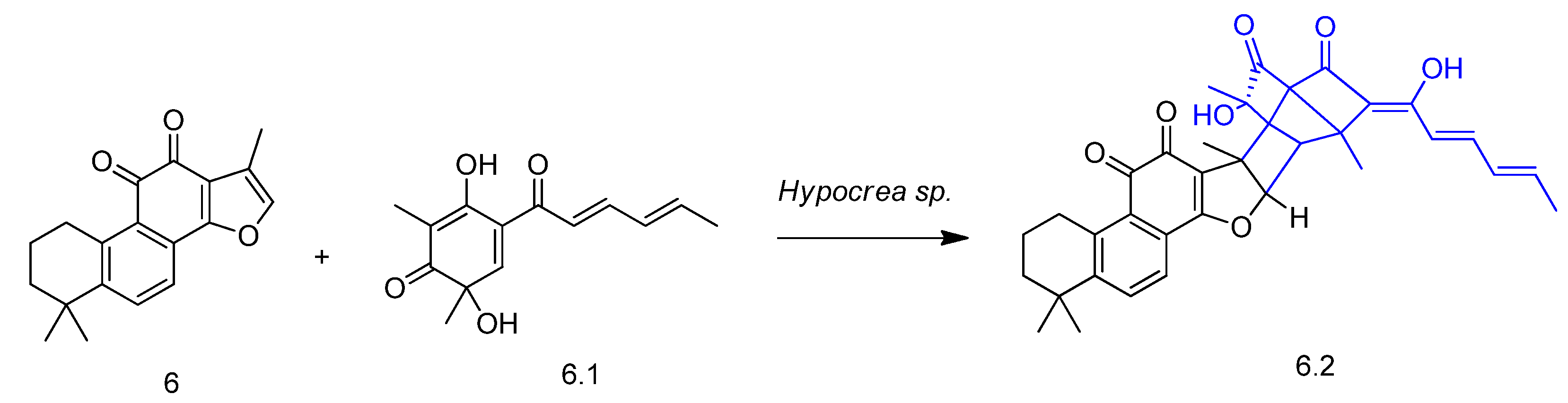
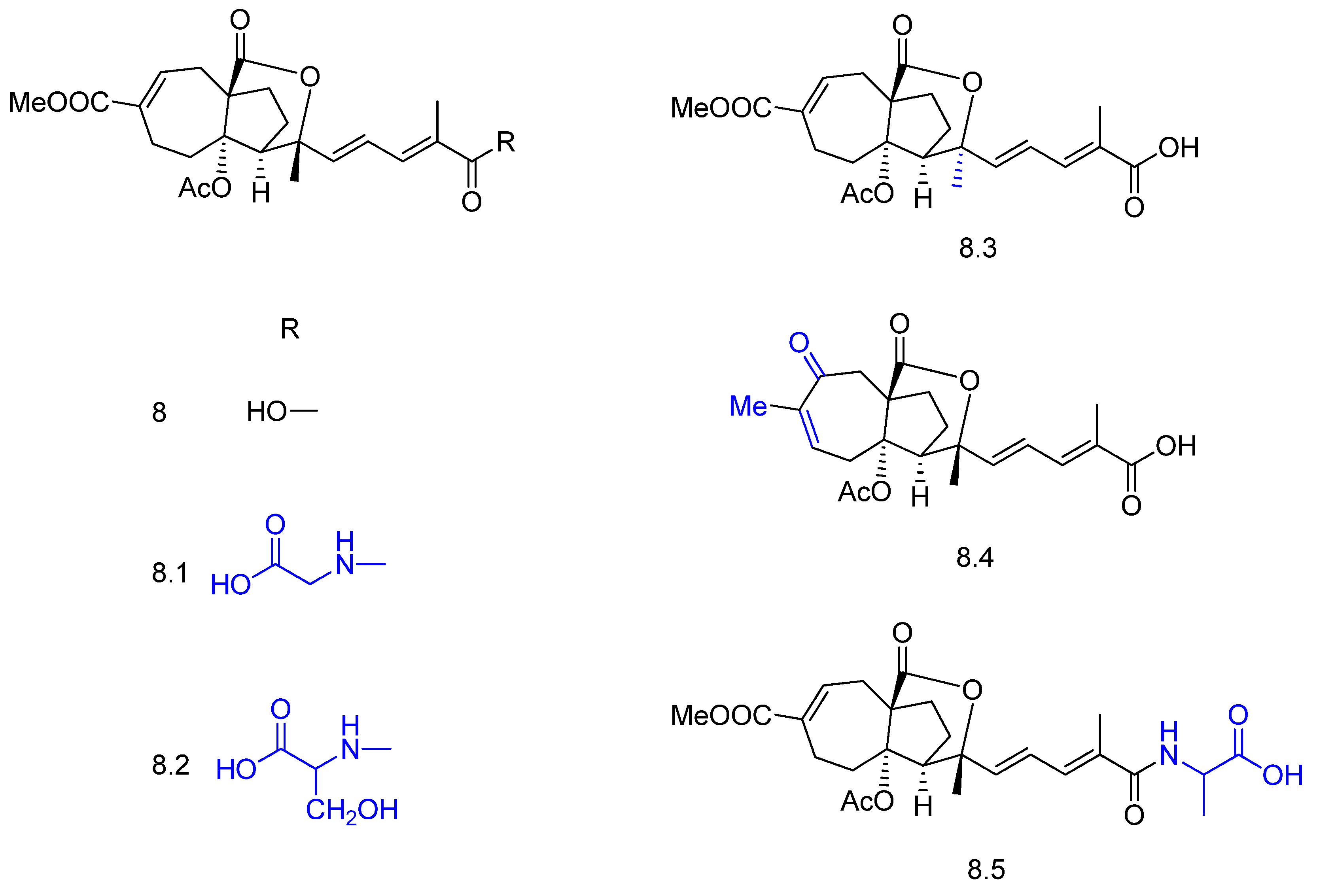
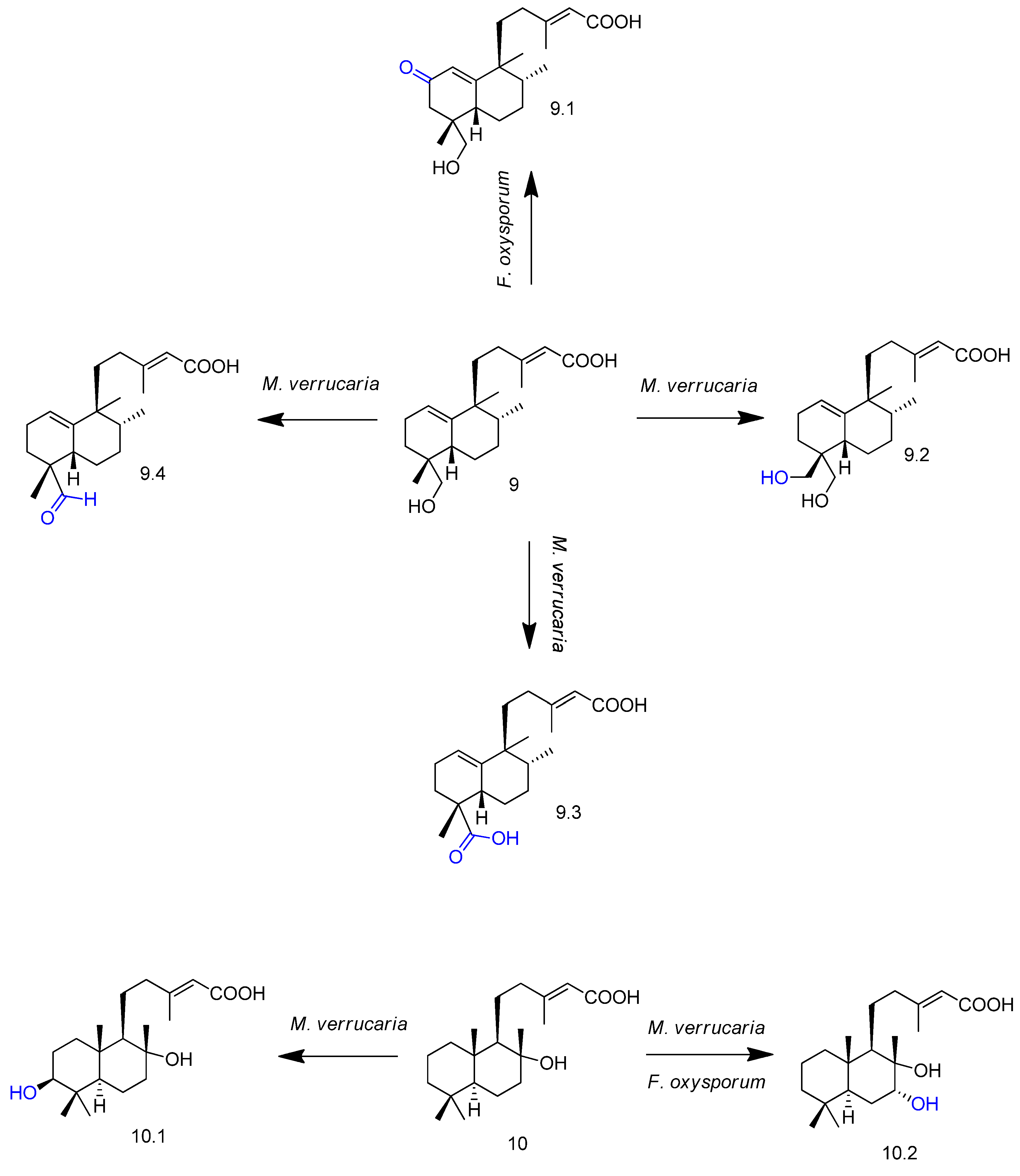
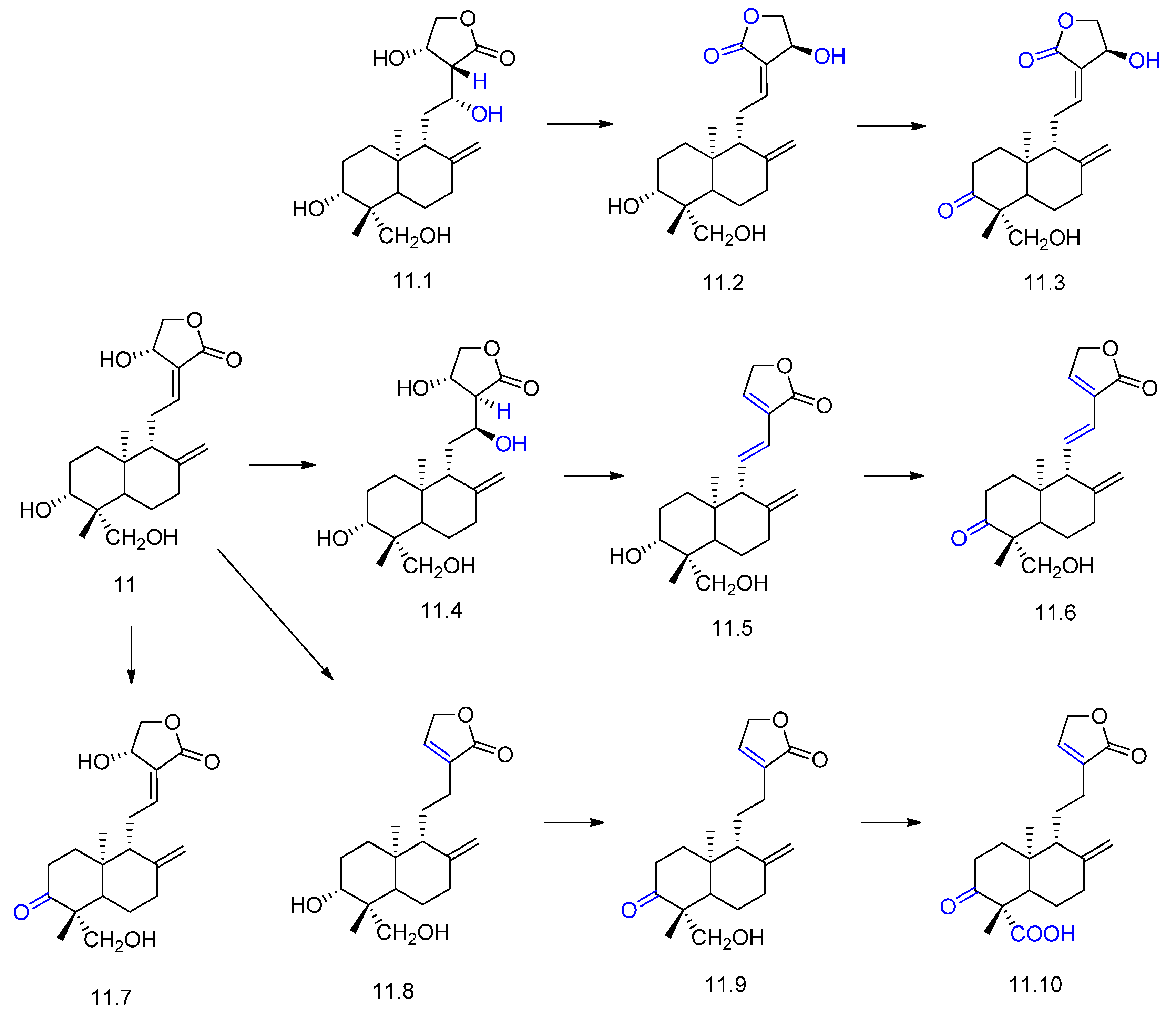
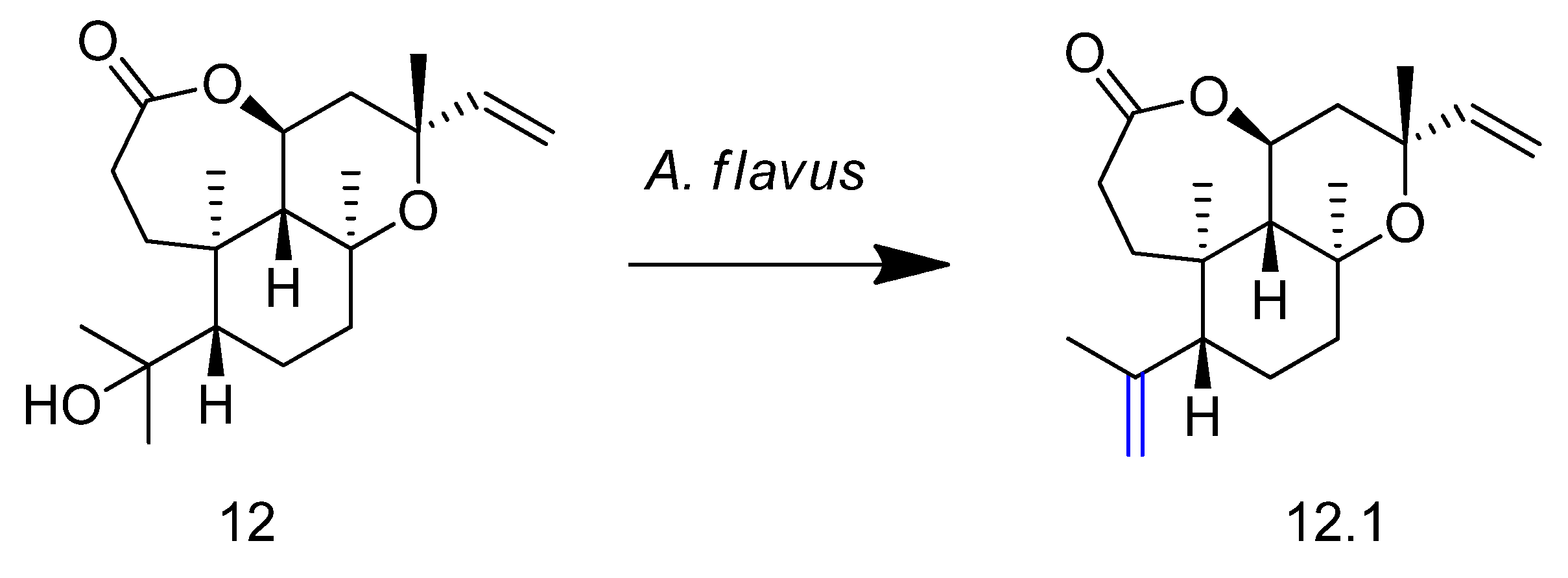
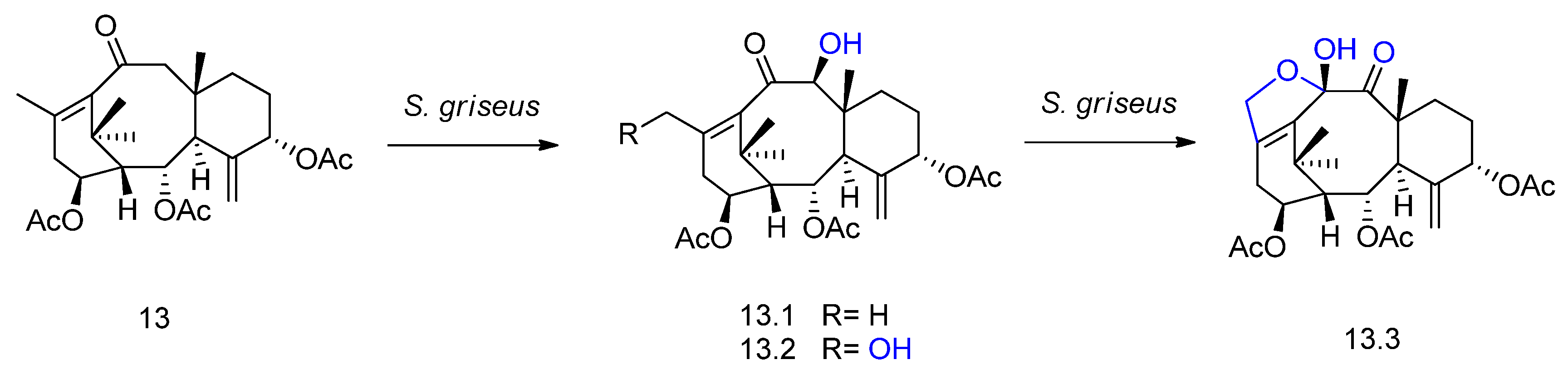
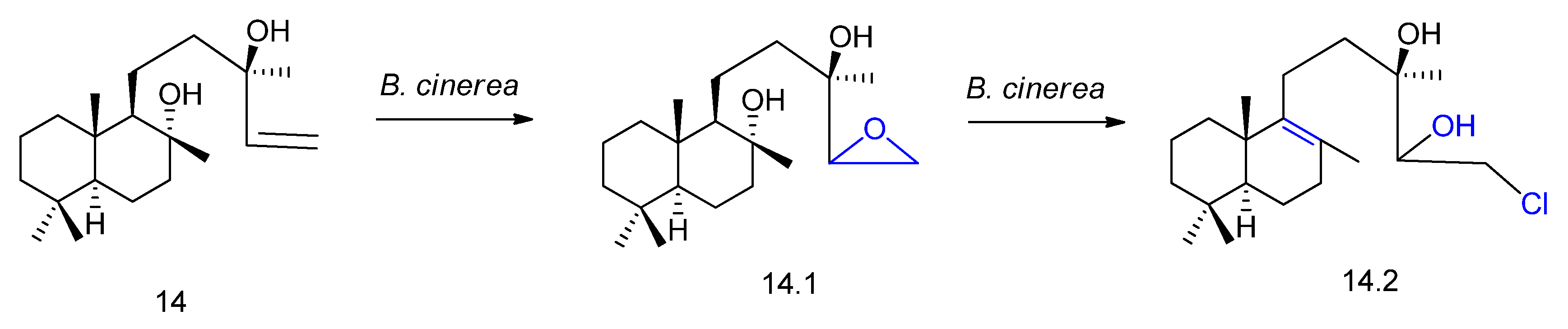
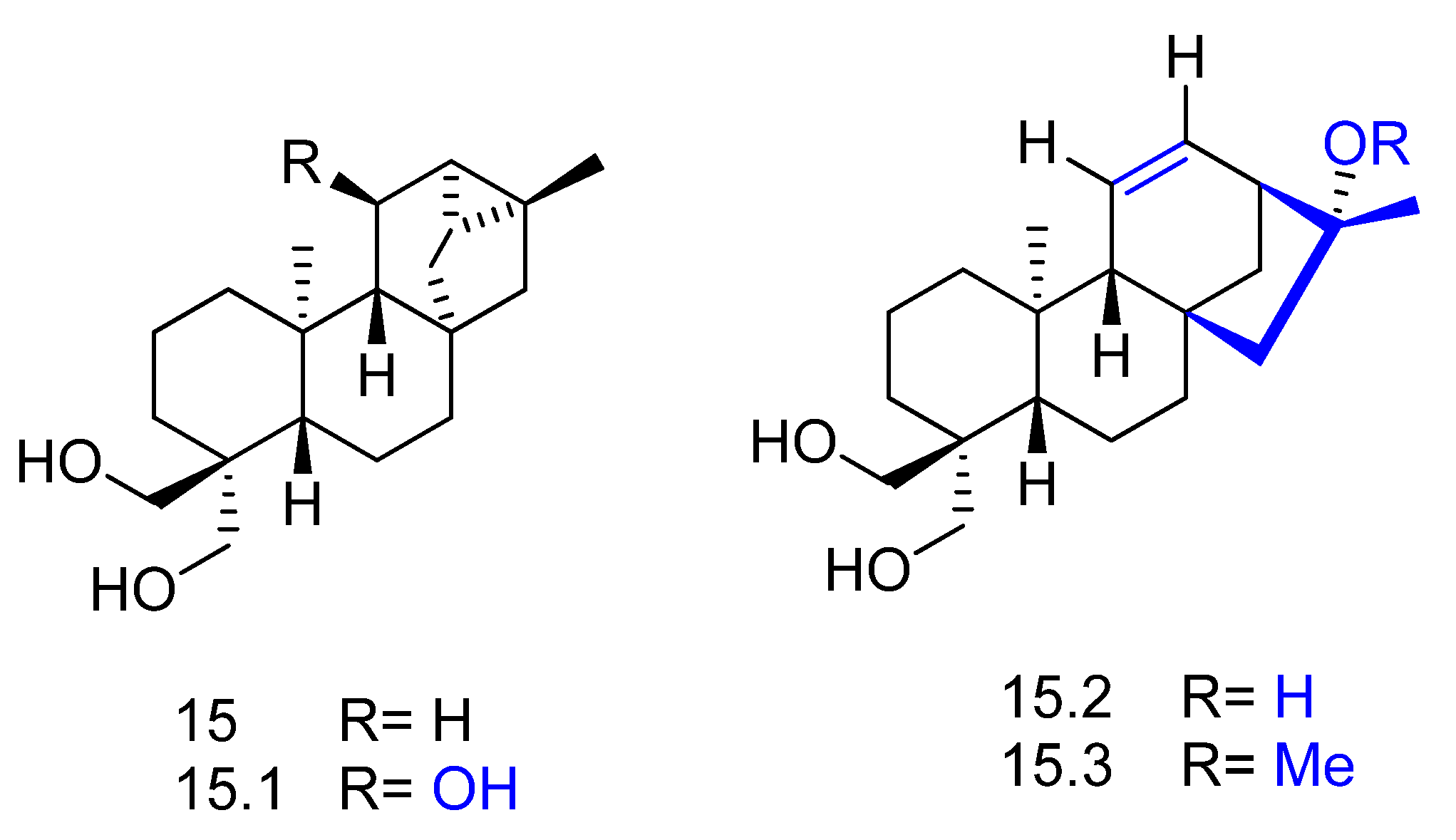
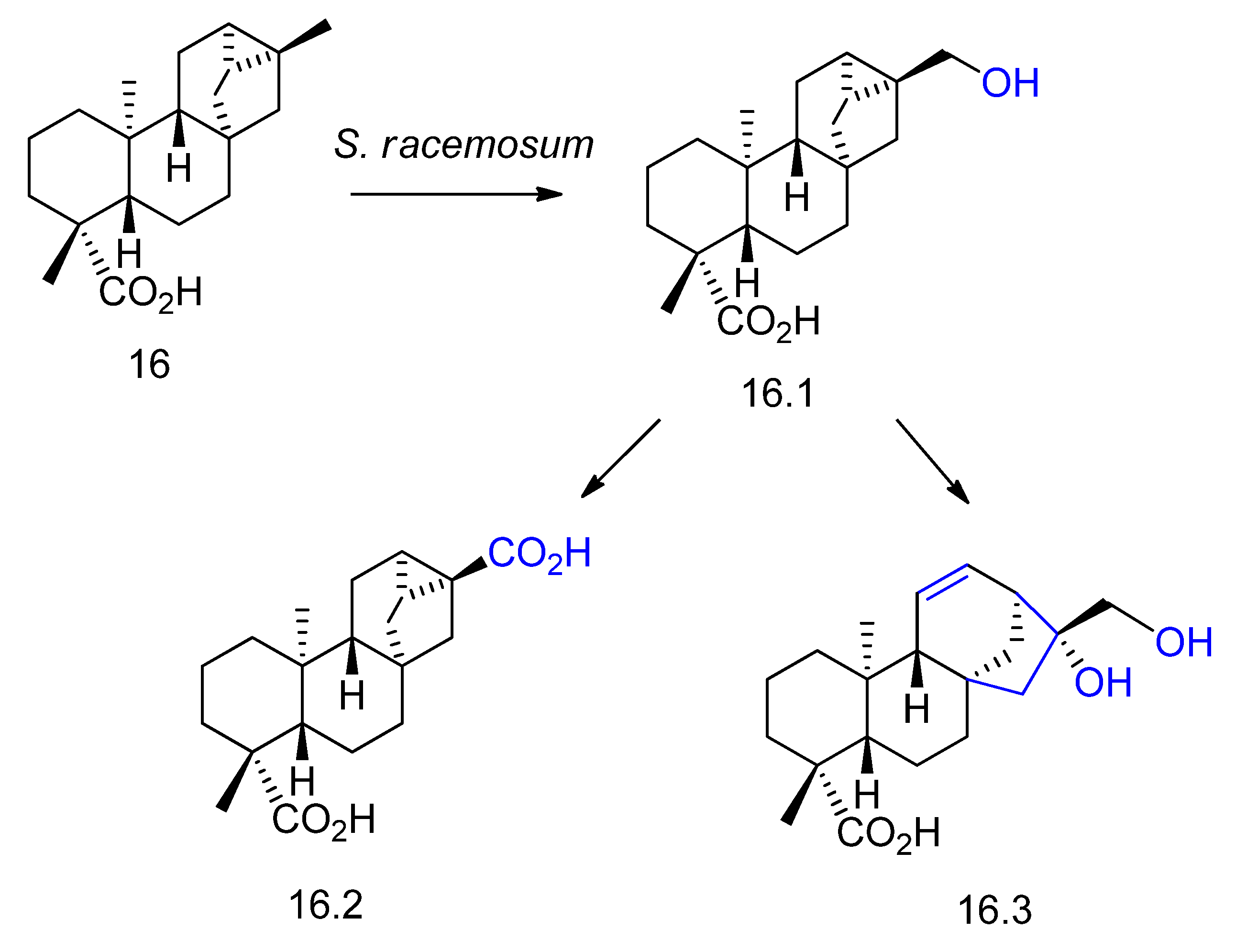
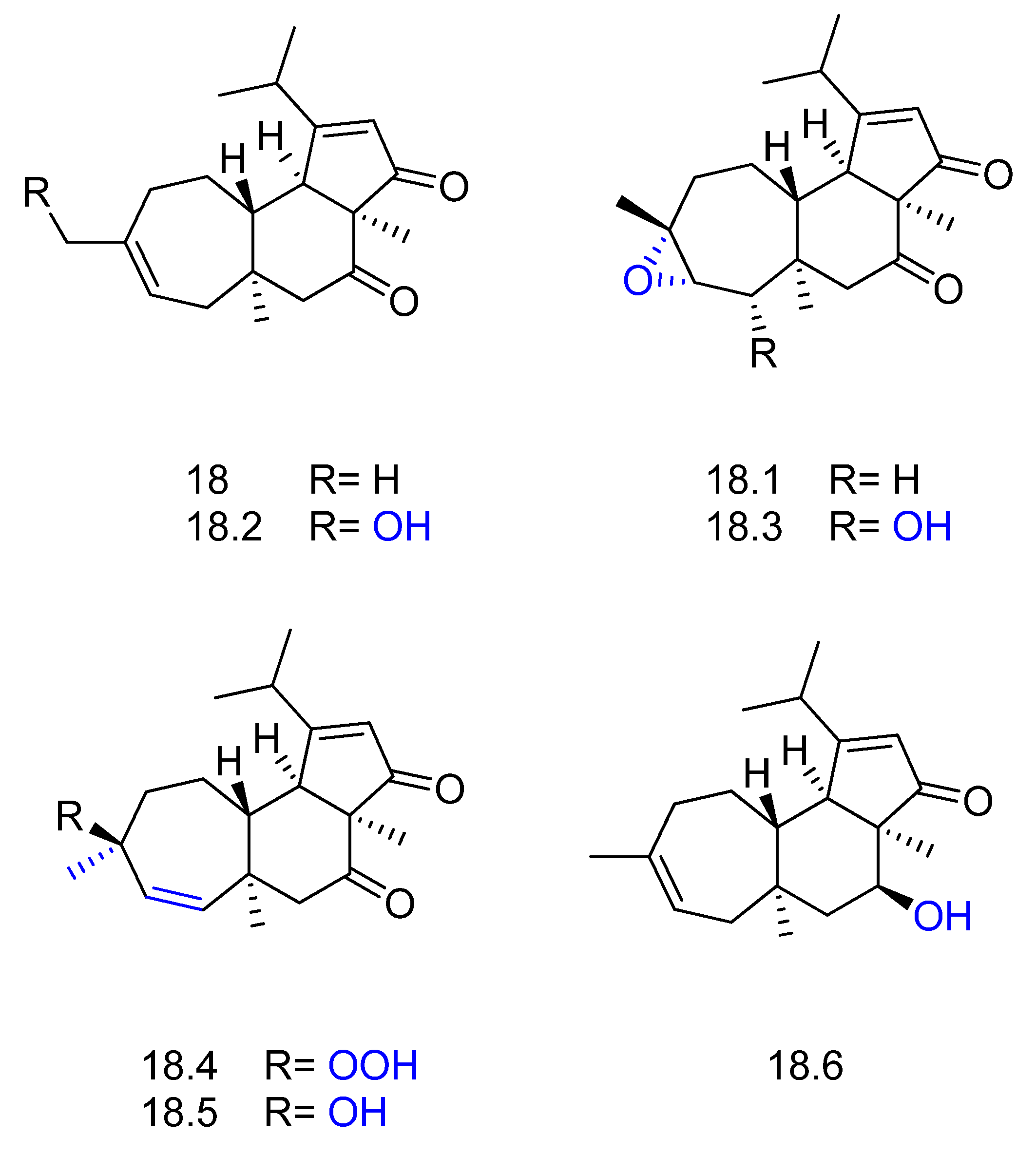
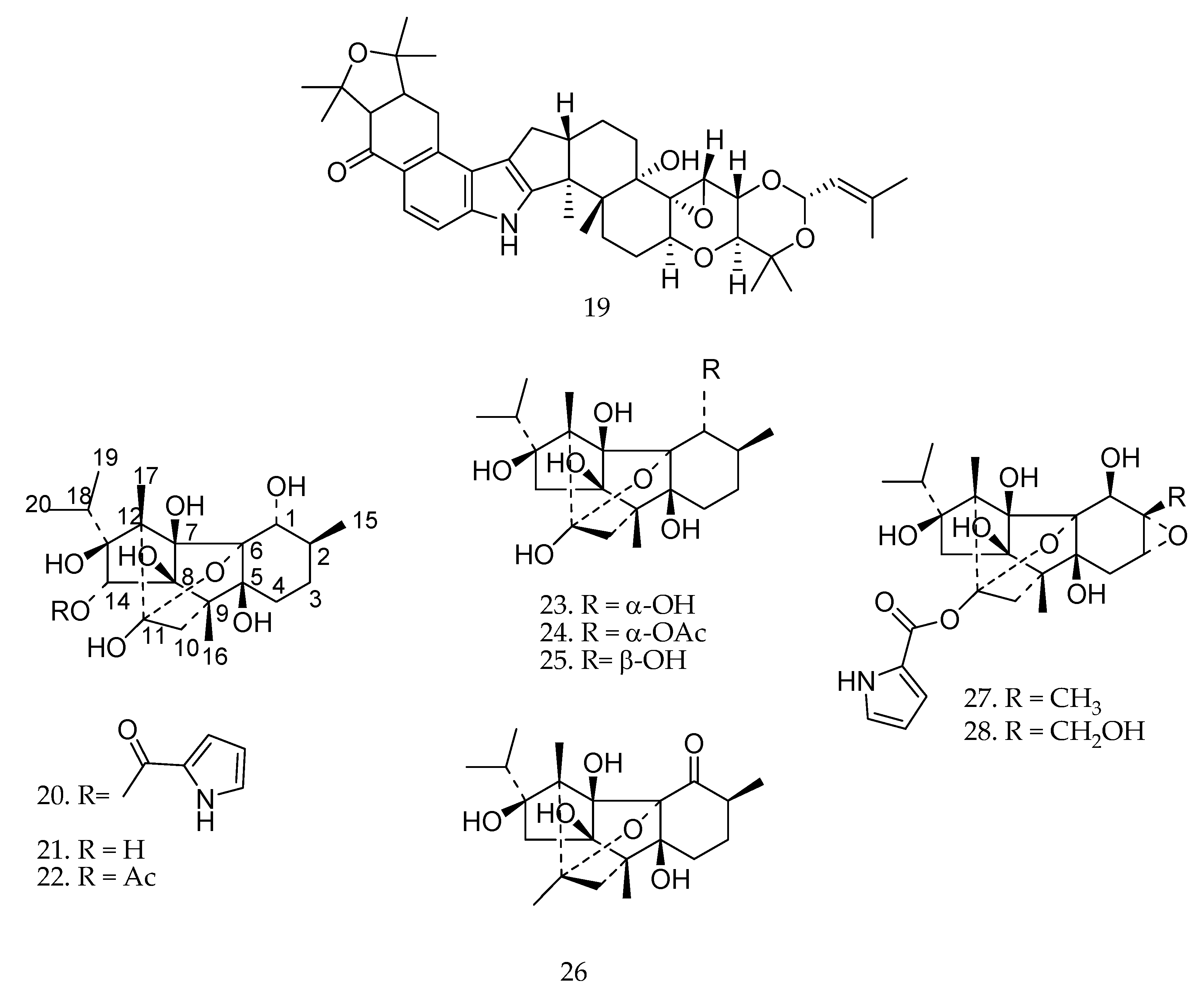
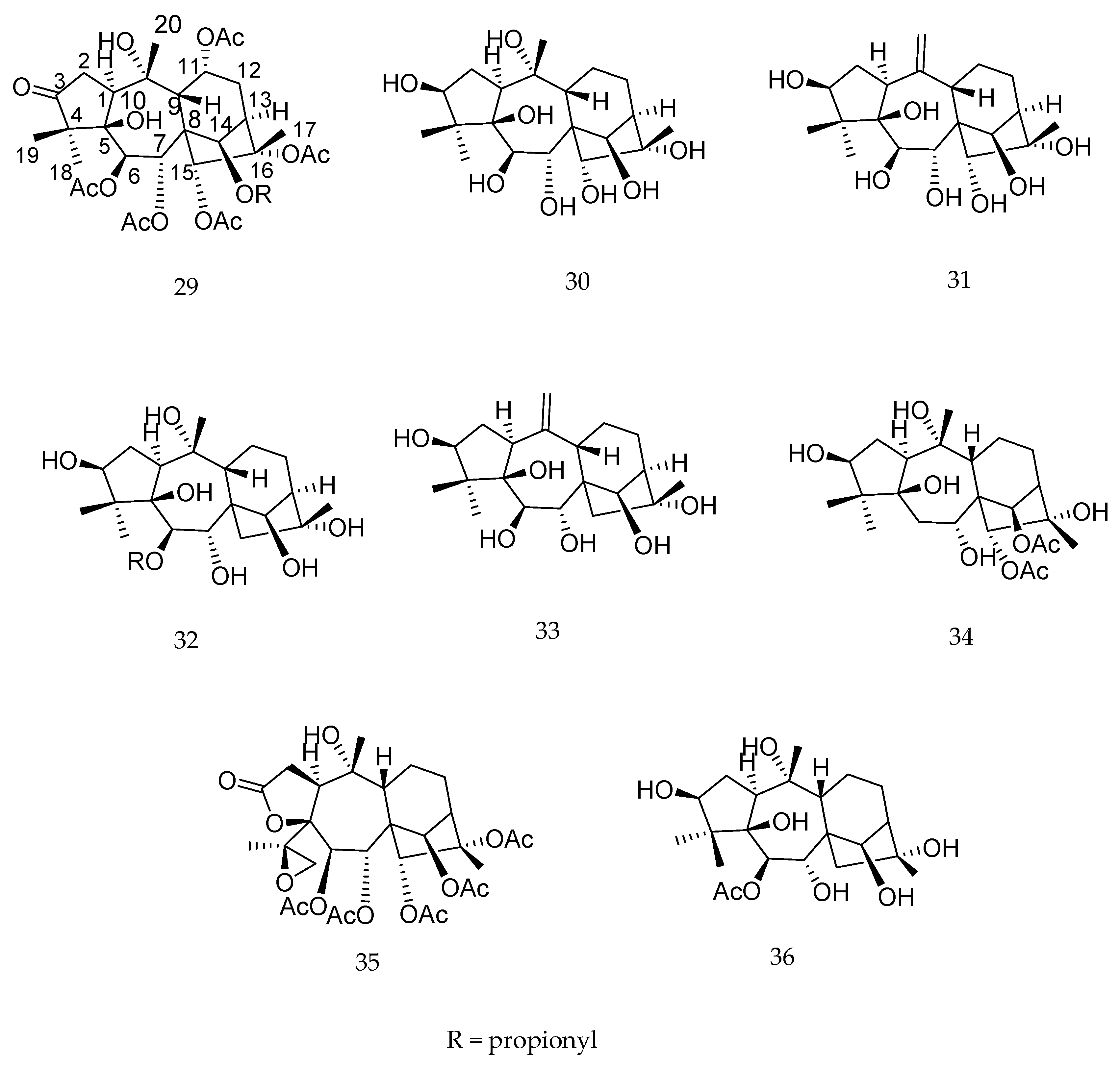
2. Biotransformation of Diterpenes
3. Toxicity of Diterpenes
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Hanson, J.R. Diterpenoids of terrestrial origin. Nat. Prod. Rep. 2017, 34, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Morris-Natschke, S.L.; Lee, K.H. Clerodane diterpenes: Sources, structures and biological activities. Nat. Prod. Rep. 2016, 33, 1166–1226. [Google Scholar] [CrossRef] [PubMed]
- Lin-Gen, L.; Ung, C.O.L.; Feng, Z.L.; Huang, L.; Hu, H. Naturally occurring diterpenoids dimers: Source, biosynthesis, chemistry and bioactivities. Planta Med. 2016, 82, 1309–1328. [Google Scholar] [CrossRef]
- Mafu, S.; Zerbe, P. Plant diterpenoid metabolism for manufacturing the biopharmaceuticals of tomorrow: Prospects and challenges. Phytochem. Rev. 2018, 17, 113–130. [Google Scholar] [CrossRef]
- Wang, X.; Yo, H.; Zhang, Y.; Lu, X.; Wang, B.; Liu, X. Bioactive pimarane–type diterpenes from marine organisms. Chem. Biodivers. 2018, 15, e1700276. [Google Scholar] [CrossRef] [PubMed]
- Dickschat, J.S. Isoprenoids in three-dimensional space: The stereochemistry of terpene biosynthesis. Nat. Prod. Rep. 2011, 28, 1917–1936. [Google Scholar] [CrossRef] [PubMed]
- Dewinck, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed.; John Wiley & Sons: Notthingham, UK, 2001; 425p, ISBN 0471-49641-3. [Google Scholar]
- Hemmelin, A.; Harwood, J.L.; Bach, T.J. A raison d’être two distinct pathways in the early steps of plants isoprenoids biosynthesis? Prog. Lipid Res. 2012, 51, 95–148. [Google Scholar] [CrossRef] [PubMed]
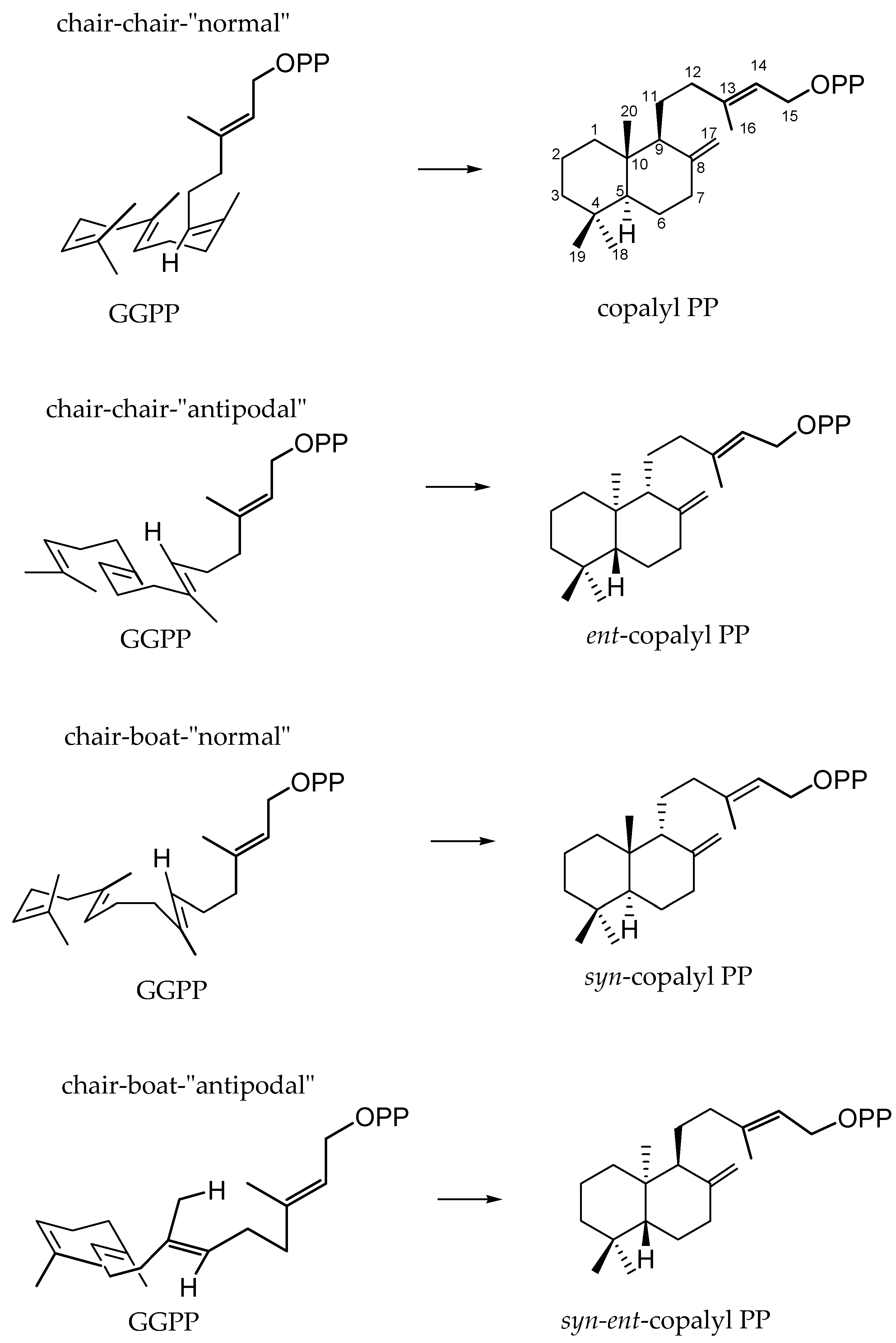
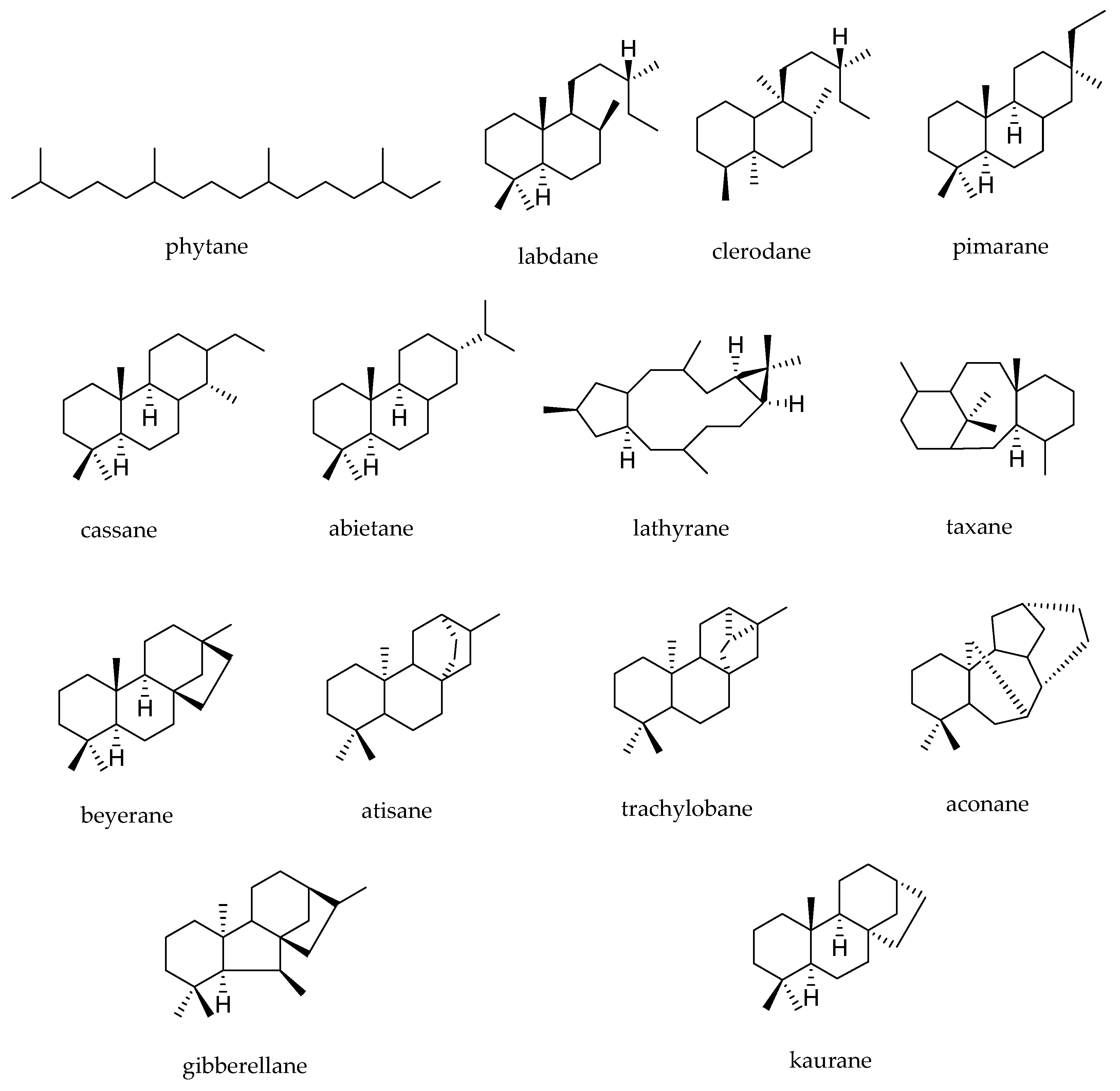
- Peters, R.J. Two rings in them all: The labdane-related diterpenoids. Nat. Prod. Rep. 2010, 27, 1521–1530. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.A.; Oliveira, A.B.; Batista, R. Occurrence, biological activities and synthesis of kaurane diterpenes and their glycosides. Molecules 2007, 12, 455–483. [Google Scholar] [CrossRef] [PubMed]
- Man, S.; Gao, W.; Wei, C.; Liu, C. Anticancer drugs from traditional toxic Chinese medicine. Phytochem. Res. 2012, 26, 1449–1465. [Google Scholar] [CrossRef] [PubMed]
- Vargas, F.S.; Almeida, P.D.O.; Aranha, E.S.P.; Boleti, A.P.A.; Newton, P.; Vasconcellos, M.C.; Veiga Junior, V.F.; Lima, E.S. Biological activities and cytotoxicity of diterpenes from Copaifera ssp. oleorresins. Molecules 2015, 20, 6194–6210. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, T.; Zhao, D.; Zheng, J.; Liu, Z. Ginkgolide B exerts cardioprotective properties against doxorubicin-induced cardiotoxicity by regulating reactive oxygen species, Akt and calcium signaling pathways in vitro and in vivo. PLoS ONE 2016, 11, e0168219. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Sun, H.X.; Xiao, H.; Cui, G.; Hillwig, M.L.; Jackson, A.; Wang, X.; Shen, Y.; Zhao, N.; Zhang, L.; et al. Combining metabolomics and transcriptomics to characterize tanshinone biosynthesis in Salvia miltiorrhiza. BMC Genom. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.K.; Laya, M.S.; Mora, H.R.; Cabrera, E.V. ChemInform abstract: The chemistry of bioactive diterpenes. Curr. Org. Chem. 2008, 12, 1050–1070. [Google Scholar] [CrossRef]
- Pal, M.; Mishra, T.; Kumar, A.; Tewari, S.K. Biological evaluation of terrestrial and marine plant originated labdane diterpene. Pharm. Chem. J. 2016, 50, 558–567. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, Q.; Ye, Y.; Lin, L. Naturally occurring furanoditerpenoids: Distribution, chemistry and their pharmacological activities. Phytochem. Rev. 2017, 16, 235–270. [Google Scholar] [CrossRef]
- Li, C.; Yan, X.; Zhang, A.L.; Gao, J.M. Structural diversity and biological activity of the genus Pieris terpenoids. J. Agric. Food Chem. 2017, 65, 9934–9949. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.O.; Furtado, N.A.J.C.; Aleu, J.; Collado, G. Non-terpenoid biotransformation by Mucor species. Phytochem. Rev. 2015, 14, 745–764. [Google Scholar] [CrossRef]
- Kebano, S.; Tesema, S.; Geleta, B. The role biotransformation in drug discovery and development. J. Drug Metab. Toxicol. 2015, 6, 2157–7609. [Google Scholar] [CrossRef]
- Roh, C.; Kang, C. Production of anti-cancer agent using microbial biotransformation. Molecules 2014, 19, 16684–16692. [Google Scholar] [CrossRef] [PubMed]
- Paludo, C.R.; Silva-Junior, E.A.; Silva, E.O.; Vessecchi, R.; Lopes, N.P.; Pupo, M.T.; Emery, F.S.; Gonçalves, N.S.; Santos, R.A.; Furtado, N.A.J.C. Inactivation of β-lapachol cytotoxicity by filamentous fungi that mimic the human blood metabolism. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.O.; De Carvalho, T.C.; Parshikov, I.A.; Dos Santos, R.A.; Emery, F.S.; Furtado, N.A.J.C. Cytotoxicity of lapachol metabolites produced by probiotics. Lett. Appl. Microbiol. 2014, 59, 108–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, E.O.; Gonçalves, N.S.; Santos, R.A.; Furtado, N.A.J.C. Microbial metabolism of atovaquone and cytotoxicity of the produced phase I metabolite. Eur. J. Drug Metab. Pharmacokinet. 2016, 41, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Wermuth, C.G.; Ganellin, C.R.; Lindberg, P.; Mitscher, L.A. Glossary of terms used in medicinal chemistry (IUPAC Recommendations 1998—online). Pure Appl. Chem. 2009, 70, 1129–1143. [Google Scholar] [CrossRef]
- Parkinson, A.; Ogilvie, B.W. Biotransformation of xenobiotics. In Cassarett & Doull’s Essentials of Toxicology, 2nd ed.; Klaassen, C.D., Watkins, J.B., III, Eds.; McGraw Hill: New York, NY, USA, 2010; 472p, ISBN 0071622403. [Google Scholar]
- Silverman, R.B.; Holladay, M.W. Drug Metabolism. In The Organic Chemistry of Drug Design and Drug Action, 3rd ed.; Academic Press: San Diego, CA, USA, 2014; 517p, ISBN 978-0-12-382030-3. [Google Scholar]
- Hudlicky, T.; Reed, J.W. Applications of biotransformations and biocatalysis to complexity generation in organic synthesis. Chem. Soc. Rev. 2009, 38, 3117–3132. [Google Scholar] [CrossRef] [PubMed]
- Hanson, J.R. An Introduction to Biotransformations in Organic Chemistry, 1st ed.; W.H. Freeman Spektrum: Oxford, UK, 1995; 92p, ISBN 0-7167-4541-0. [Google Scholar]
- Gonzalez, F.J.; Coughtrie, M.; Tukey, R.H. Drug Metabolism. In Goodman and Gilman’s—The Pharmacological Basis of Therapeutics, 12th ed.; Brunton, L.L., Chabner, B.A., Knollmann, B.C., Eds.; McGraw Hill: New York, NY, USA, 2011; 2084p, ISBN 978-0-07-162442-8. [Google Scholar]
- Argikar, U.A.; Dumouchel, J.L.; Dunne, C.E.; Saran, C.; Cirello, A.L.; Gunduz, M. Ocular metabolism of levobunolol; historic and emerging metabolic pathways. Drug Metab. Dispos. 2016, 44, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- Koppel, N.; Rekdal, V.M.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356. [Google Scholar] [CrossRef] [PubMed]
- Fasinu, P.; Bouic, P.J.; Rosenkranz, B. Liver-based in vitro technologies for drug biotransformation studies—A review. Curr. Drug MeTab. 2012, 13, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.C.C.R.; Fonseca, M.M.R. Biotransformation of terpenes. Biotechnol. Adv. 2006, 24, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Orbach, S.M.; Less, R.R.; Kothari, A.; Rajagopalan, P. In vitro intestinal and liver models for toxicity testing. ACS Biomater. Sci. Eng. 2017, 3, 1898–1910. [Google Scholar] [CrossRef]
- Salerno, S.; Bartolo, L. Biohybrid membrane systems and bioreactors as tools for in vitro drug testing. Curr. Pharm. Des. 2017, 23, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Gröger, M.; Dinger, J.; Kiehntopf, M.; Peters, F.T.; Rauen, U.; Mosig, A.S. Preservation of cell structure, metabolism, and biotransformation activity of liver-on-chip organ models by hypothermic storage. Adv. Healthc. Mater. 2017, 7, e1700616. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yang, S.; Zhang, H.; Zhou, J.; Li, Y.; Zhang, J.; Jin, Y.; Wang, Z.; Li, Y.; Shen, J.; et al. Comprehensive analysis of tiamulin metabolites in various species of farm animals using ultra-high-performance liquid chromatography coupled to quadrupole/time-of-flight. J. Agric. Food Chem. 2017, 65, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Husser, C.; Pahler, A.; Seymour, M.; Kuhlmann, O.; Schadt, S.; Zell, M. Profiling of dalcetrapib metabolites in human plasma by accelerator mass spectrometry and investigation of the free phenothiol by derivatisation with methylacrylate. J. Pharm. Biomed. Anal. 2018, 152, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Wakai, S.; Arazoe, T.; Ogino, C.; Kondo, A. Future insights in fungal metabolic engineering. Bioresour. Technol. 2017, 245, 1314–1326. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, S. In silico ADME-Tox modeling: Progress and prospects. Expert Opin. Drug Metab. Toxicol. 2017, 13, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Sultana, N.; Saify, Z.S. Enzymatic biotransformation of terpenes as bioactive agents. J. Enzyme Inhib. Med. Chem. 2013, 28, 1113–1128. [Google Scholar] [CrossRef] [PubMed]
- García-Granados, A.; Martínez, A.; Parra, A.; Rivas, F. Manoyl-oxide biotransformations with filamentous fungi. Curr. Org. Chem. 2007, 11, 679–692. [Google Scholar] [CrossRef]
- Bhatti, H.N.; Khera, R.A. Biotransformations of diterpenoids and triterpenoids: A review. J. Asian Nat. Prod. Res. 2014, 16, 70–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Dai, H.; Li, X.; Li, Y.; Wang, L.; Xue, M. Structural elucidation of metabolites of tanshinone I and its analogue dihydrotanshinone I in rats by HPLC-ESI-MSn. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ye, M.; Gong, Y.; Feng, L.; Tao, S.; Yin, J.; Guo, D. Biotransformation of pseudolaric acid B by Chaetomium globosum. Process Biochem. 2011, 46, 2064–2067. [Google Scholar] [CrossRef]
- Carneiro, L.J.; Bianchi, T.C.; da Silva, J.J.M.; Oliveira, L.C.; Borges, C.H.G.; Lemes, D.C.; Bastos, J.K.; Veneziani, R.C.S.; Ambrósio, S.R. Development and validation of a rapid and reliable RP-HPLC-PDA method for the quantification of six diterpenes in Copaifera duckei, Copaifera reticulata and Copaifera multijuga oleoresins. J. Braz. Chem. Soc. 2018, 29, 729–737. [Google Scholar] [CrossRef]
- González-Burgos, E.; Carretero, M.E.; Gómez-Serranillos, M.P. Sideritis spp.: Uses, chemical composition and pharmacological activities—A review. J. Ethnopharmacol. 2011, 135, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guo, J.; Tang, Y.; Wu, L.; Tao, W.; Qian, Y.; Duan, J. Pharmacokinetic profile and metabolite identification of yuanhuapine, a bioactive component in Daphne genkwa by ultra-high-performance liquid chromatography coupled with tandem mass spectrometry. J. Pharm. Biomed. Anal. 2015, 112, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Jin, Y.; Ma, Y.; Xie, W.; Xu, H.; Zhang, K.; Zhang, L.; Du, Y. Identification of metabolites of oridonin in rats with a single run on UPLC-Triple-TOF-MS/MS system based on multiple mass defect filter data acquisition and multiple data processing techniques. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 1006, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, J.; Wang, X.; Cai, Z. Study of the phase I and phase II metabolism of a mixture containing multiple tanshinones using liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 2992–2998. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, Y.; Su, H.; Wang, Q.; Li, G.; Hu, Y.; Jiang, J.; Tan, B.; Qiu, F. Metabolic characteristics of tanshinone I in human liver microsomes and S9 subcellular fractions. Xenobiotica 2018. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, M.; Li, X.; Zhang, X.; Abdel-Mageed, W.M.; Li, L.; Wang, W.; Zhang, J.; Han, J.; Dai, H.; et al. Fungal biotransformation of tanshinone results in [4+2] cycloaddition with sorbicillinol: Evidence for enzyme catalysis and increased antibacterial activity. Appl. Microbiol. Biotechnol. 2016, 100, 8349–8357. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Li, Y.; Qin, Y.; Tong, X.; Song, Z.; Zhao, Y.; Wei, R.; Li, L.; Dai, H.; Wang, W.; et al. New cryptotanshinone derivatives with anti-influenza A virus activities obtained via biotransformation by Mucor rouxii. Appl. Microbiol. Biotechnol. 2017, 101, 6365–6374. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, A.F.; Seidl, C.; Severino, V.G.P.; Cardoso, C.L.; Castro-Gamboa, I. Biotransformation of labdane and halimane diterpenoids by two filamentous fungi strains. R. Soc. Open Sci. 2017, 4, 170854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Wang, Y.; Hu, H.; Wu, Y.; Zeng, X. Novel bioconversion products of andrographolide by Aspergillus ochraceus and their cytotoxic activities against human tumor cell lines. J. Mol. Catal. B Enzym. 2011. [Google Scholar] [CrossRef]
- He, X.; Zeng, X.; Hu, H.; Wu, Y. Cytotoxic biotransformed products from andrographolide by Rhizopus stolonifer ATCC 12939. J. Mol. Catal. B Enzym. 2010, 62, 242–247. [Google Scholar] [CrossRef]
- Sura, M.B.; Ankireddy, M.; Ponnapalli, M.G. Biotransformation of agallochaexcoerin A by Aspergillus flavus. Nat. Prod. Res. 2015, 29, 838–841. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xie, D.; Chen, R.; Mei, M.; Zou, J.; Chen, X.; Dai, J. A furantaxane with an unusual 6/8/6/5 ring system and potent tumor MDR reversal activity obtained via microbial transformation. Org. Lett. 2012, 14, 4106–4109. [Google Scholar] [CrossRef] [PubMed]
- Farooq, A.; Tahara, S. Biotransformation of two cytotoxic terpenes, α-santonin and sclareol by Botrytis cinerea. Z. Naturforsch. C 2000, 55c, 713–717. [Google Scholar] [CrossRef]
- Frija, L.M.T.; Frade, R.F.M.; Afonso, C.A.M. Isolation, chemical, and biotransformation routes of labdane-type diterpenes. Chem. Rev. 2011, 111, 4418–4452. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.P.; Ouazzani, J.; Arcile, G.; Jeller, A.H.; Lima, J.P.F.; Seleghim, M.H.R.; Oliveira, A.L.L.; Debonsi, H.M.; Venâncio, T.; Yokoya, N.S.; et al. Biohydroxylation of (−)-Ambrox®, (−)-sclareol, and (+)-sclareolide by whole cells of Brazilian marine-derived fungi. Mar. Biotechnol. 2015, 17, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.; Sepúlveda, C.; Melo, R.; Cotoras, M. Characterization of the antifungal activity against Botrytis cinerea of sclareol and 13-epi-sclareol, two labdane-type diterpenoids. J. Chil. Chem. Soc. 2015, 60, 3024–3028. [Google Scholar] [CrossRef]
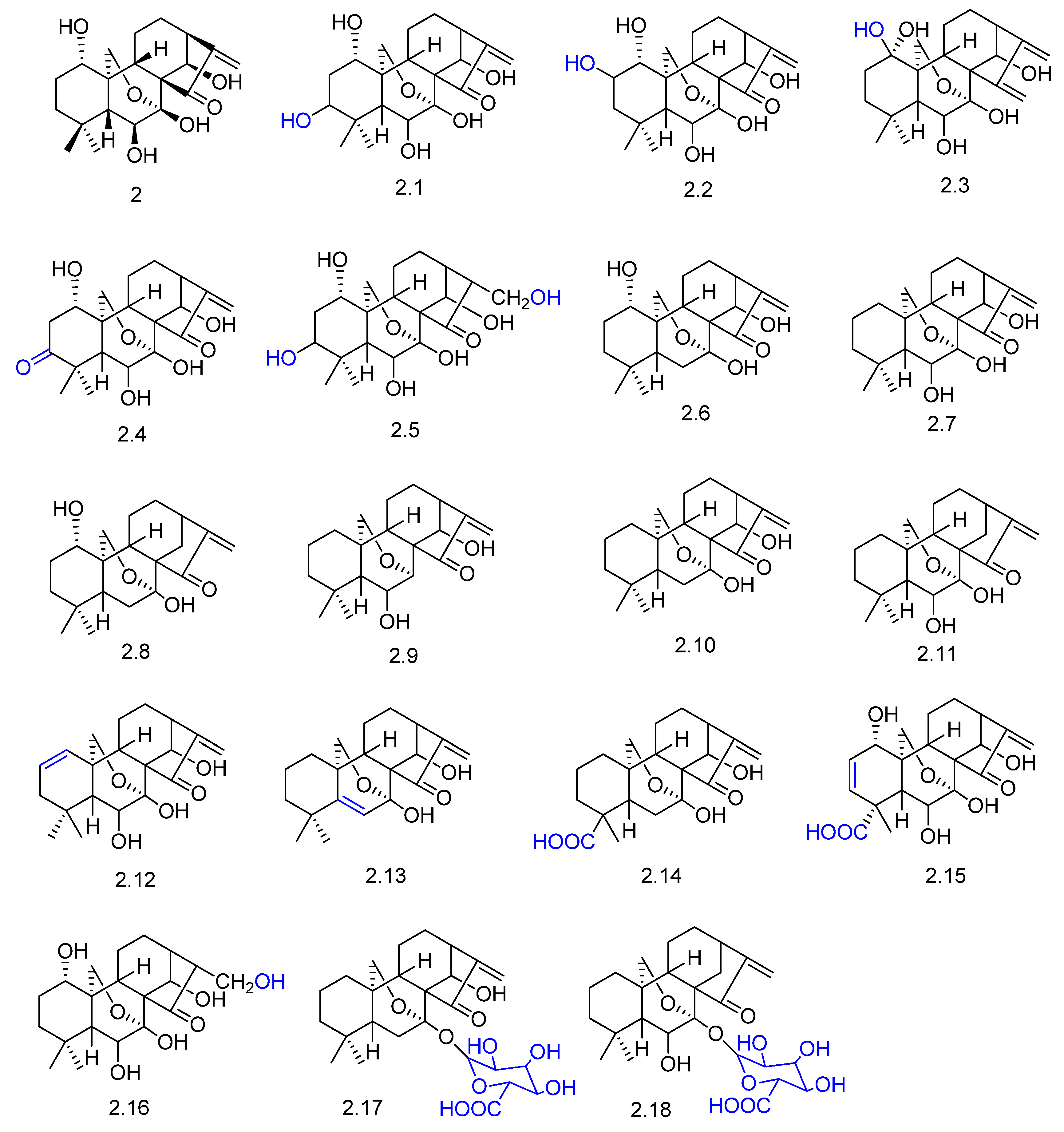
- Vasconcelos, D.H.P.; Mafezoli, J.; Uchôa, P.K.S.; Saraiva, N.N.; Lima, M.A.S.; Júnior, J.N.S.; Barbosa, F.G.; Mattos, M.C.; Oliveira, M.C.F.; Lima, C.S.; et al. Biotransformation of the diterpene ent-18,19-dihydroxytrachylobane by Rhizopus stolonifer. J. Braz. Chem. Soc. 2015, 26, 1043–1047. [Google Scholar] [CrossRef]
- Dos Santos, G.F.; Lima, G.S.; Oliveira, G.P.; Souza Filho, J.D.; Amaral, L.S.; Rodrigues-Filho, E.; Takahashi, J.A. New AChE inhibitors from microbial transformation of trachyloban-19-oic acid by Syncephalastrum racemosum. Bioorg. Chem. 2018, 79, 60–63. [Google Scholar] [CrossRef] [PubMed]
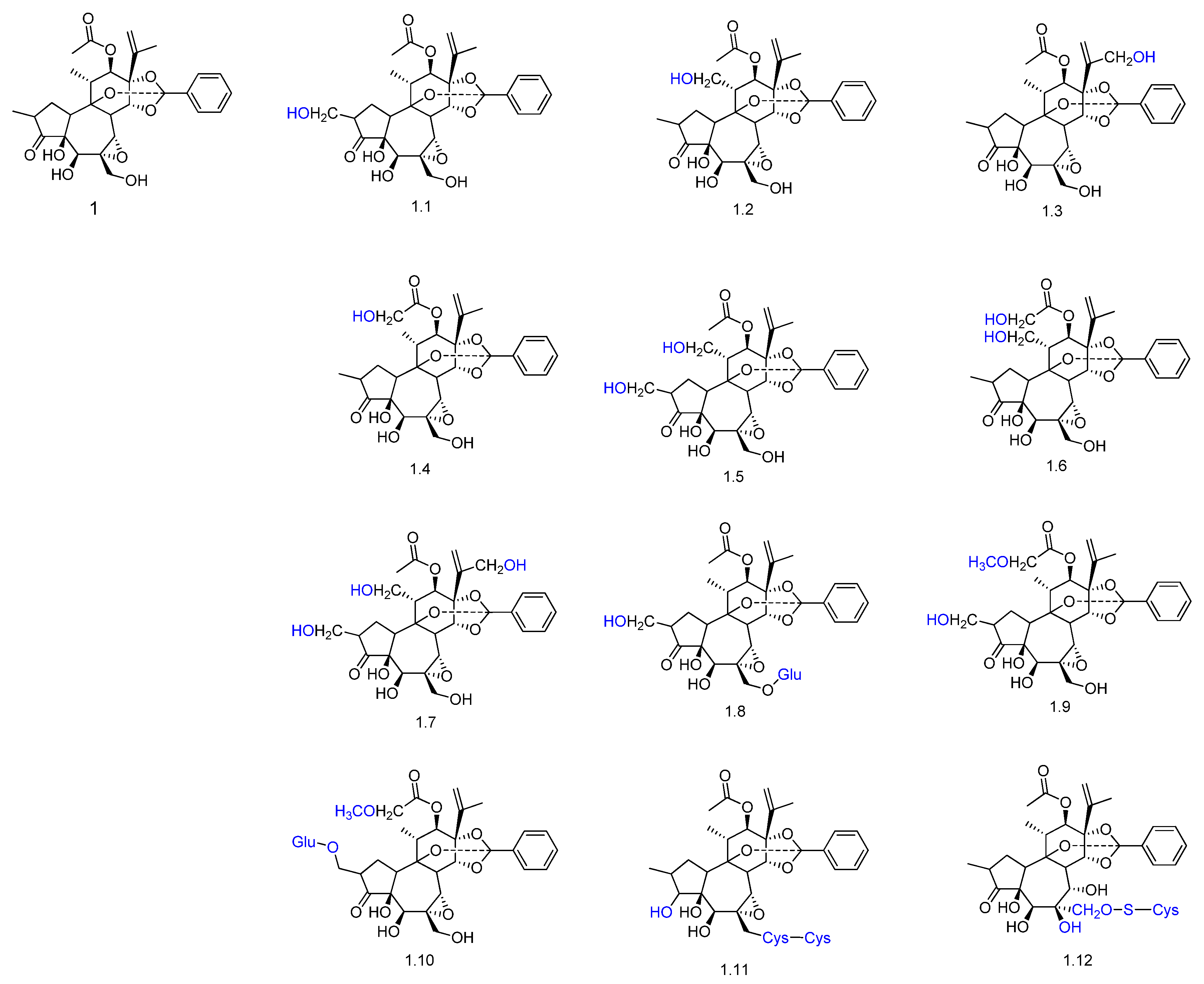
- Teng, R.; McManus, D.; Aylward, J.; Ogbourne, S.; Armstrong, D.; Mau, S.; Johns, J.; Bacic, A. Biotransformation of ingenol-3-angelate in four plant cell suspension cultures. Biocatal. Biotransform. 2009, 27, 186–194. [Google Scholar] [CrossRef]
- Peng, J.; Kasanah, N.; Stanley, C.E.; Chadwick, J.; Fronczek, F.R.; Hamann, M.T. Microbial metabolism studies of cyanthiwigin B and synergic antibiotic effects. J. Nat. Prod. 2006, 69, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Rico-Martínez, M.; Medina, F.G.; Marrero, J.G.; Osegueda-Robles, S. Biotransformation of diterpenes. RSC Adv. 2014, 4, 10627–10647. [Google Scholar] [CrossRef]
- Takahashi, J.A.; Gomes, D.C.; Lyra, F.H.; Santos, G.F.; Martins, L.R. The remarkable structural diversity achieved in ent-kaurane diterpenes by fungal biotransformations. Molecules 2014, 19, 1856–1886. [Google Scholar] [CrossRef] [PubMed]
- Jeschek, M.; Panke, S.; Ward, T.R. Artificial metalloenzymes on the verge of new-to-nature metabolism. Trends Biotechnol. 2018, 36, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, D.; Freundlich, J.S. Antimycobacterial metabolism: Illuminating Mycobacterium tuberculosis biology and drug discovery. Trends Microbiol. 2017, 25, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.; Ramírez-Apan, M.T.; Delgado, G. Biotransformation of sclareolide by filamentous fungi: Cytotoxic evaluation of the derivatives. J. Braz. Chem. Soc. 2011, 22, 1177–1182. [Google Scholar] [CrossRef]
- Rocha, A.D.; Vieira, H.S.; Takahashi, J.A.; Boaventura, M.A.D. Synthesis of a new allelopathic agent from the biotransformation of ent-15α-hydroxy-16-kauren-19-oic acid with Fusarium proliferatum. Nat. Prod. Res. 2017, 31, 2647–2653. [Google Scholar] [CrossRef] [PubMed]
- Porto, T.S.; Simão, M.R.; Carlos, L.Z.; Martins, C.H.G.; Furtado, N.A.J.C.; Said, S.; Arakawa, N.S.; Santos, R.A.; Veneziani, R.C.S.; Ambrósio, S.R. Pimarane-type diterpenes obtained by biotransformation: Antimicrobial properties against clinically isolated Gram-positive multidrug-resistant bacteria. Phytother. Res. 2013, 27, 1502–1507. [Google Scholar] [CrossRef] [PubMed]
- Sepúlveda, B.; Quispe, C.; Simirgiotis, M.; García-Beltrán, O.; Areche, C. Gastroprotective effects of new diterpenoid derivatives from Azorella cuatrecasasii Mathias & Constance obtained using a β-cyclodextrin complex with microbial and chemical transformations. Bioorg. Med. Chem. Lett. 2016, 26, 3220–3222. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chang, S.; Lin, W.; Chou, B.; Wang, L.; Liu, P.; Lin, S. Oxygenated compounds from the bioconversion of isostevic acid and their inhibition of TNF-α and COX-2 expressions in LPS-stimulated RAW 264.7 cells. Phytochemistry 2012, 75, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhang, B.J.; Wang, C.Y.; Tian, Y.; Yao, J.H.; An, L.; Huang, S.S.; Peng, J.Y.; Liu, K.X.; Ma, X.C. Microbial transformation of deoxyandrographolide and their inhibitory activity on LPS-induced NO production in RAW 264.7 macrophages. Bioorg. Med. Chem. Lett. 2012, 22, 1615–1618. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Huang, Y.; Feng, F.; Chen, Z. Efficient enzymatic synthesis and antibacterial activity of andrographolide glycoside. Process Biochem. 2016, 51, 675–680. [Google Scholar] [CrossRef]
- Chen, Z.; Zhu, Q.; Zong, M.; Gu, Z.; Han, Y. Enzymatic synthesis and antibacterial activity of andrographolide derivatives. Process Biochem. 2011, 46, 1649–1653. [Google Scholar] [CrossRef]
- Leverrier, A.; Martin, M.T.; Servy, C.; Ouazzani, J.; Retailleau, P.; Awang, K.; Mukhtar, M.R.; Gueritte, F.; Litaudon, M. Rearranged diterpenoids from the biotransformation of ent-trachyloban-18-oic acid by Rhizopus arrhizus. J. Nat. Prod. 2010, 73, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Cao, Y.; Liu, X.; Cheng, Z.H. Regio- and stereo-selective hydroxylations of ingenane diterpenoids by Mortierella ramanniana and Gibberella fujikuroi. Chin. J. Nat. Med. 2016, 14, 939–945. [Google Scholar] [CrossRef]
- Kumar, R.; Saha, A.; Saha, D. Biotransformation of 16-oxacleroda-3,13(14)E-dien-15-oic acid isolated from Polyalthia longifolia by Rhizopus stolonifera increases its antifungal activity. Biocatal. Biotransform. 2016, 34, 212–218. [Google Scholar] [CrossRef]
- Venisetty, R.K.; Ciddi, V. Application of microbial biotransformation for the new drug discovery using natural drugs as substrates. Curr. Pharm. Biotechnol. 2003, 4, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Fraga, B.M.; González-Vallejo, V.; Guillermo, R. On the biotransformation of ent-trachylobane to ent-kaur-11-ene diterpenes. J. Nat. Prod. 2011, 74, 1985–1989. [Google Scholar] [CrossRef] [PubMed]
- Mafezoli, J.; Oliveira, M.C.F.; Paiva, J.R.; Sousa, A.H.; Lima, M.A.S.; Silva Júnior, J.N.; Barbosa, F.G.; Wijeratne, K.; Gunatilaka, A.A.L. Stereo and regioselective microbial reduction of the clerodane diterpene 3,12-dioxo-15,16-epoxy-4-hydroxycleroda-13(16),14-diene. Nat. Prod. Commun. 2014, 9, 759–762. [Google Scholar] [PubMed]
- Chen, Z.G.; Tan, R.X.; Huang, M. Efficient regioselective acylation of andrographolide catalyzed by immobilized Burkholderia cepacia lipase. Process Biochem. 2010, 45, 415–418. [Google Scholar] [CrossRef]
- Ruan, Y.; Ma, B.J.; Li, L.Z.; Wang, X.L. Specific 12α-hydroxylation of grandiflorenic acid by permeabilised fungus Fusarium graminearum. Nat. Prod. Res. 2014, 28, 677–679. [Google Scholar] [CrossRef] [PubMed]
- De Graan, A.M.; Elens, L.; Sprowl, J.A.; Sparreboom, A.; Friberg, L.E.; Van der Holt, B.; De Raaf, P.J.; De Bruijn, P.; Engels, F.K.; Eskens, F.A.L.M.; et al. CYP3A4*22 genotype and systemic exposure affect paclitaxel-induced neurotoxicity. Clin. Cancer Res. 2013, 19, 3316–3324. [Google Scholar] [CrossRef] [PubMed]
- Yarom, N.; Cyjon, A.; Kovel, S.; Yachin, A.; Sella, A.; Evron, E. Epistaxis during treatment with paclitaxel. Basic Clin. Pharmacol. Toxicol. 2009, 104, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Micali, G.; Lacarruba, F.; Nasca, M.R.; Schwartz, R. Topical pharmacotherapy for skin cancer: Part I. Pharmacology. J. Am. Acad. Dermatol. 2014, 70. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Cheng, F.; Yao, W.; Bao, B.; Zhang, K.; Zhang, L.; Ding, A. Toxicity of pekinenin C from Euphorbia pekinensis radix on rat small intestinal crypt epithelial cell and its apoptotic mechanism. Int. J. Mol. Sci. 2016, 17, 850. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Z.; Zhang, R.; Hou, P.; Bi, K.; Chen, X. Nephrotoxicity evaluation of a new cembrane diterpene from Euphorbiae pekinensis Radix with HEK 293T cells and the toxicokinetics study in rats using a sensitive and reliable UFLC–MS/MS. J. Pharm. Biomed. Anal. 2016, 119, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Zayed, S.M.; Farghaly, M.; Soliman, S.M.; Gotta, H.; Sorg, B.; Hecker, E. Dietary cancer risk from conditional cancerogens (tumor promoters) in produce of livestock fed on species of spurge (Euphorbiaceae). V. Skin irriitant and tumor-promoting diterpene ester toxins of the tigliane and ingenane type in the herbs Euphorbia nubica and Euphorbia helioscopia contaminating fodder of livestock. J. Cancer Res. Clin. Oncol. 2001, 127, 40–47. [Google Scholar] [PubMed]
- Zayed, S.M.; Farghaly, M.; Taha, H.; Gotta, H.; Hecker, E. Dietary cancer risk from conditional cancerogens in produce of livestock fed on species of spurge (Euphorbiaceae) I. Skin irritant and tumor-promoting ingenane-type diterpene esters in E. peplus, one of several herbaceous Euphorbia species contaminating fodder of livestock. J. Cancer Res. Clin. Oncol. 1998, 124, 131–140. [Google Scholar] [CrossRef]
- Philippe, G. Lolitrem B and indole diterpene alkaloids produced by endophytic fungi of the genus Epichloë and their toxic effects in livestock. Toxins 2016, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Imlach, W.L.; Finch, S.C.; Dunlop, J.; Meredith, A.L.; Aldrich, R.W.; Dalziel, J.E. The molecular mechanism of “ryegrass staggers,” a neurological disorder of K+ channels. J. Pharmacol. Exp. Ther. 2008, 327, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kang, J.; Cao, X.; Sun, X.; Yu, S.; Zhang, X.; Sun, H.; Guo, Y. Characterization of diterpenes from Euphorbia prolifera and their antifungal activities against phytopathogenic fungi. J. Agric. Food Chem. 2015, 63, 5902–5910. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Luo, S.H.; Li, S.H.; Gao, J.M. New antifeedant grayanane diterpenoids from the flowers of Pieris formosa. Molecules 2017, 22, e1431. [Google Scholar] [CrossRef] [PubMed]
- Thacker, J.R.M. An Introduction to Arthropod Pest Control; Cambridge University Press: London, UK, 2002; 380p, ISBN 978-0521561068. [Google Scholar]
- González-Coloma, A.; Terrero, D.; Perales, A.; Escoubas, P.; Fraga, B.M. Insect antifeedant ryanodane diterpenes from Persea indica. J. Agric. Food Chem. 1996, 4, 296–300. [Google Scholar] [CrossRef]
- González-Coloma, A.; Reina, M.; Gutiérrez, C.; Fraga, B.M. Natural insecticides: Structure diversity, effects and structure-activity relationships. A case study. Stud. Nat. Prod. Chem. 2002, 26, 849–879. [Google Scholar] [CrossRef]
- Niu, C.S.; Li, Y.; Liu, Y.B.; Ma, S.G.; Liu, F.; Cui, L.; Yu, H.B.; Wang, X.J.; Qu, J.; Yu, S.S. Grayanane diterpenoids with diverse bioactivities from the roots of Pieris formosa. Tetrahedron 2018, 74, 375–382. [Google Scholar] [CrossRef]
- Chen, X.Q.; Gao, L.H.; Li, Y.P.; Li, H.M.; Liu, D.; Liao, X.L.; Li, R.T. Highly oxygenated grayanane diterpenoids from flowers of Pieris japonica and structure−activity relationships of antifeedant activity against Pieris brassicae. J. Agric. Food Chem. 2017, 65, 4456–4463. [Google Scholar] [CrossRef] [PubMed]
- Coll, J.; Tandrón, Y.A. Neo-clerodane diterpenoids from Ajuga: Structural elucidation and biological activity. Phytochem. Rev. 2008, 7, 25–49. [Google Scholar] [CrossRef]
- Sivasubramanian, A.; Narasimha, K.K.G.; Rathnasamy, R.; Campos, A.M. A new antifeedant clerodane diterpenoid from Tinospora cordifolia. Nat. Prod. Res. 2013, 27, 1431–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozov, P.I.; Vasileva, T.A.; Iliev, I.N. Structure and antifeedant activity relationship of neo-clerodane diterpenes against colorado potato beetle larvae. Chem. Nat. Compd. 2014, 50, 762–764. [Google Scholar] [CrossRef]
- Gebbinck, E.A.K.; Jansen, B.J.; De Groot, A. Insect antifeedant activity of clerodane diterpenes and related model compounds. Phytochemistry 2002, 61, 737–770. [Google Scholar] [CrossRef]
- Tang, W.; Wei, X.; Xu, H.; Zeng, D.; Long, L. 13-Deoxyitol A, a new insecticidal isoryanodane diterpene from the seeds of Itoa orientalis. Fitoterapia 2009, 80, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Vattikonda, S.R.; Sangam, S.R. Effect of forskolin on the growth and differentiation of the ovary of Papilio demoleus L. (Lepidoptera: Papilionidae). Int. Res. J. Environ. Sci. 2017, 6, 13–17. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Avolio, F.; Santini, A.; Tuzi, A.; Berestetskyi, A.; Vurro, M.; Evidente, A. Chenopodolin: A phytotoxic unrearranged ent-pimaradiene diterpene produced by Phoma chenopodicola, a fungal pathogen for Chenopodium album biocontrol. J. Nat. Prod. 2013, 76, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.; Espinoza, P.; Urzua, A.; Vivanco, M.; Cotoras, M. In vitro antifungal activity of the diterpenoid 7α-hydroxy-8(17)- labden-15-oic acid and its derivatives against Botrytis cinerea. Molecules 2009, 14, 1966–1979. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Xu, Q.-L.; He, C.-M.; Zeng, L.; Wang, H.-F. Two new anti-fungal diterpenoids from the husks of Oryza sativa. Phytochem. Lett. 2014, 10, 309–312. [Google Scholar] [CrossRef]
- Shah, A.N.; Iqbal, J.; Ullah, A.; Yang, G.; Yousaf, M.; Fahad, S.; Tanveer, M.; Hassan, W.; Tung, S.A.; Wang, L.; et al. Allelopathic potential of oil seed crops in production of crops: A review. Environ. Sci. Pollut. Res. Int. 2016, 23, 14854–14867. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |




























{kind=link}
{kind=link}
{kind=link}
{kind=link}
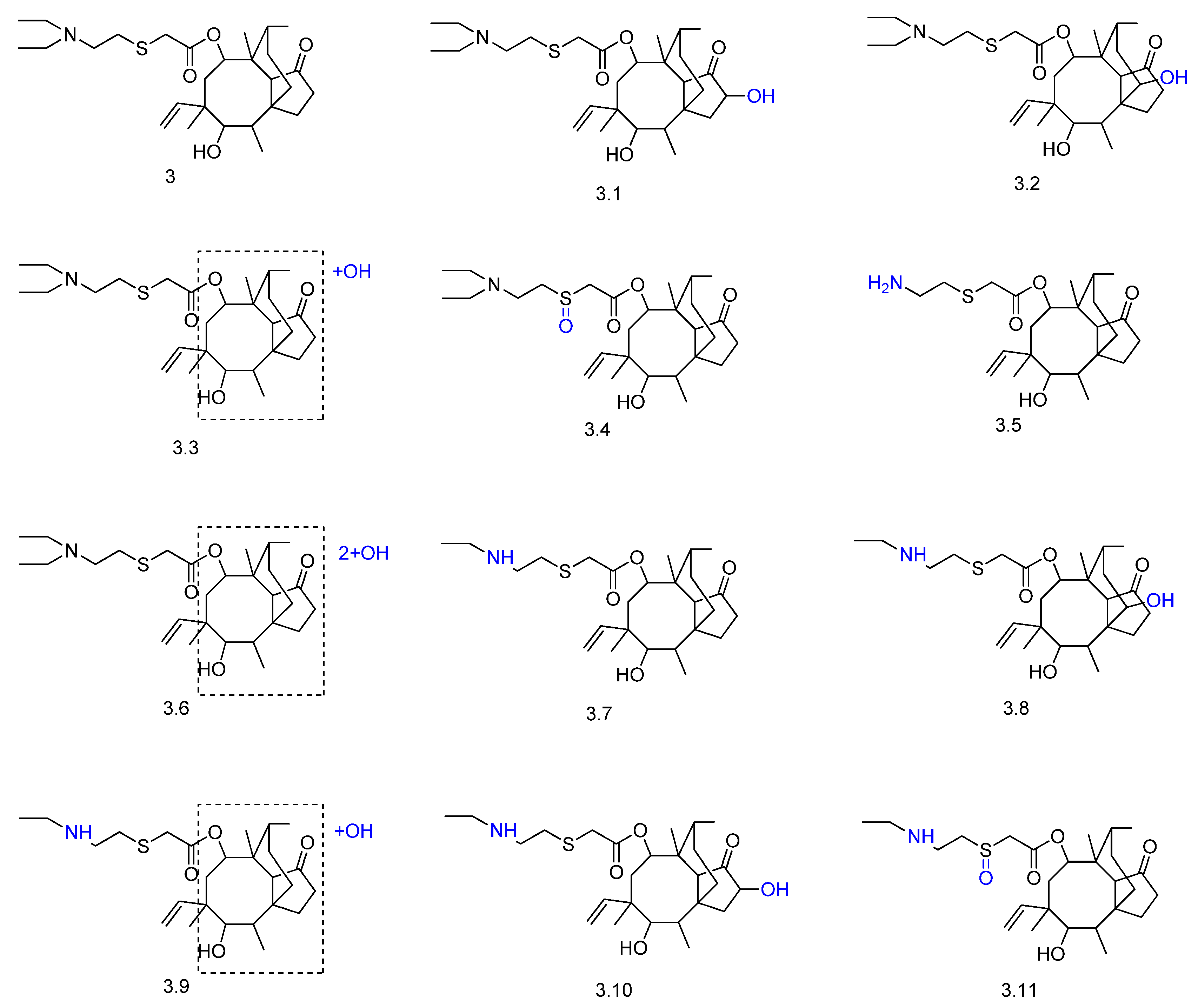{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
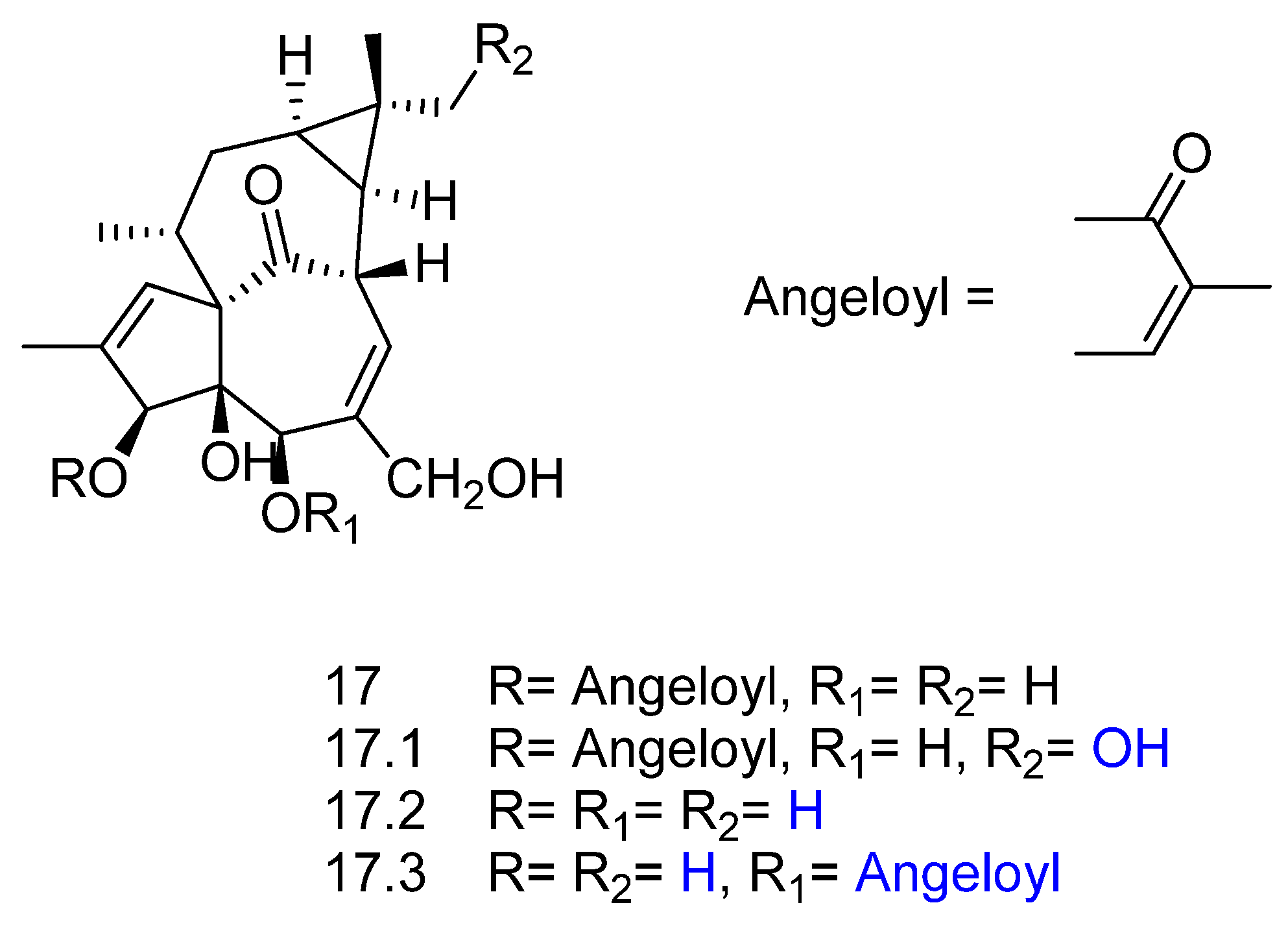{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
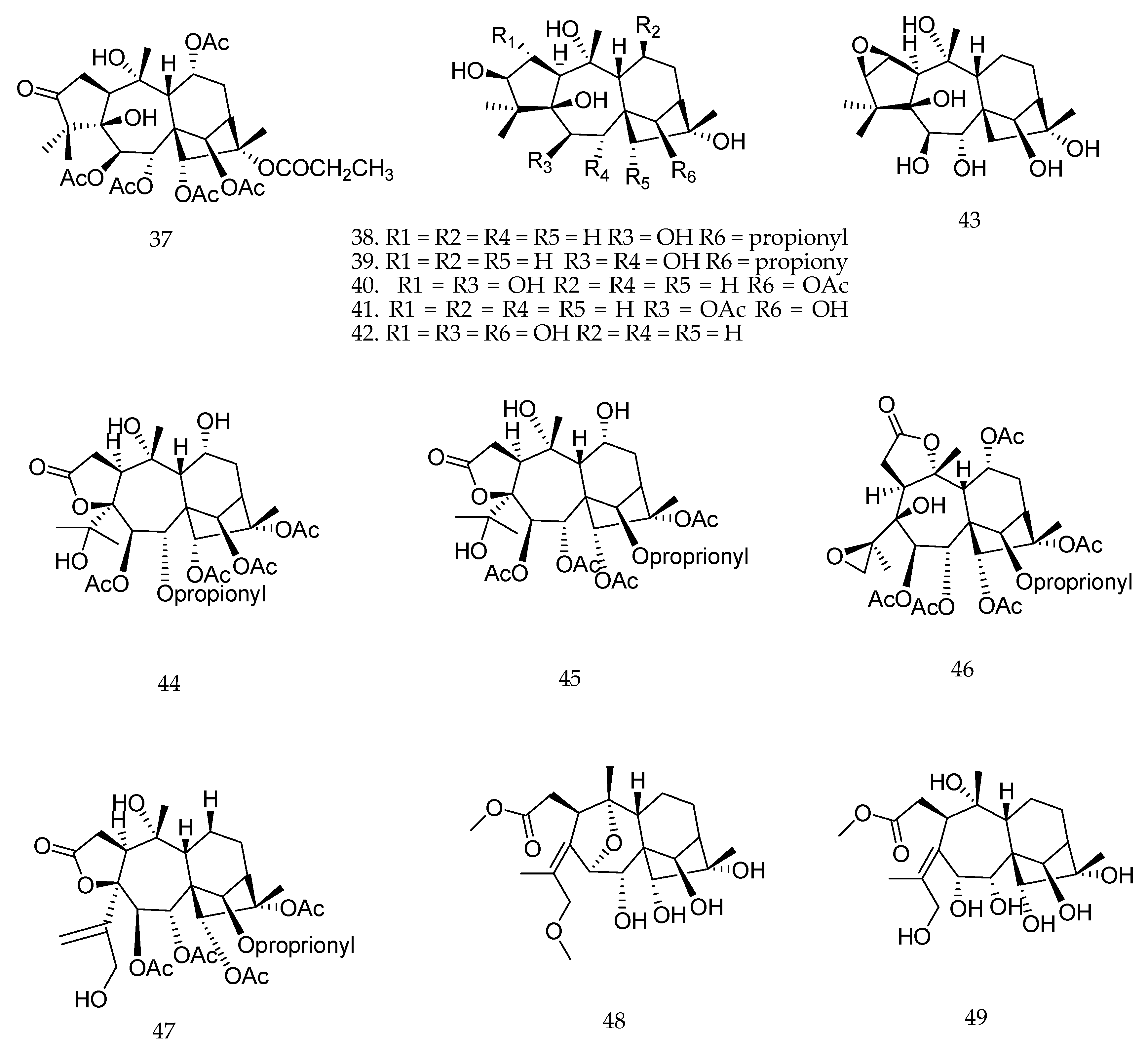
| Parent Compound | Biotransformation Biocatalyst | Biological Activity Evaluated | Enzymatic Reaction | Effect on Biological Activity * | References |
|---|---|---|---|---|---|
| sclareolide | fungi | cytotoxicity in vitro | hydroxylation | enhance | [72] |
| ingenol-3-angelate | plant cell cultures | cytotoxicity in vitro | Hydroxylation deacylation | decrease decrease | [66] |
| ent-15α-hydroxy-kaur-16-en-19-oic | fungi | allelopathic | hydroxylation | enhance | [73] |
| ent-8(14),15-pimaradiene | fungi | antibacterial | hydroxylation dihydroxylation | enhance no effect | [74] |
| mulin-11,13-dien-20 oic acid | fungi | gastroprotective in vivo | hydroxylation dihydroxylation | enhance enhance | [75] |
| 20-deoxyingenol | fungi | cytotoxicity in vitro | hydroxylation | no effect | [81] |
| 13-oxyingenol dodecanoate | fungi | cytotoxicity in vitro | hydroxylation | decrease | [81] |
| pseudolaric acid B | fungi | antifungal | conjugation with amino acids epimerization migration of double bond | decrease decrease decrease | [46] |
| ent-trachyloban-18-oic acid | fungi | cytotoxicity in vitro | hydroxylation backbone rearrangement | decrease decrease | [80] |
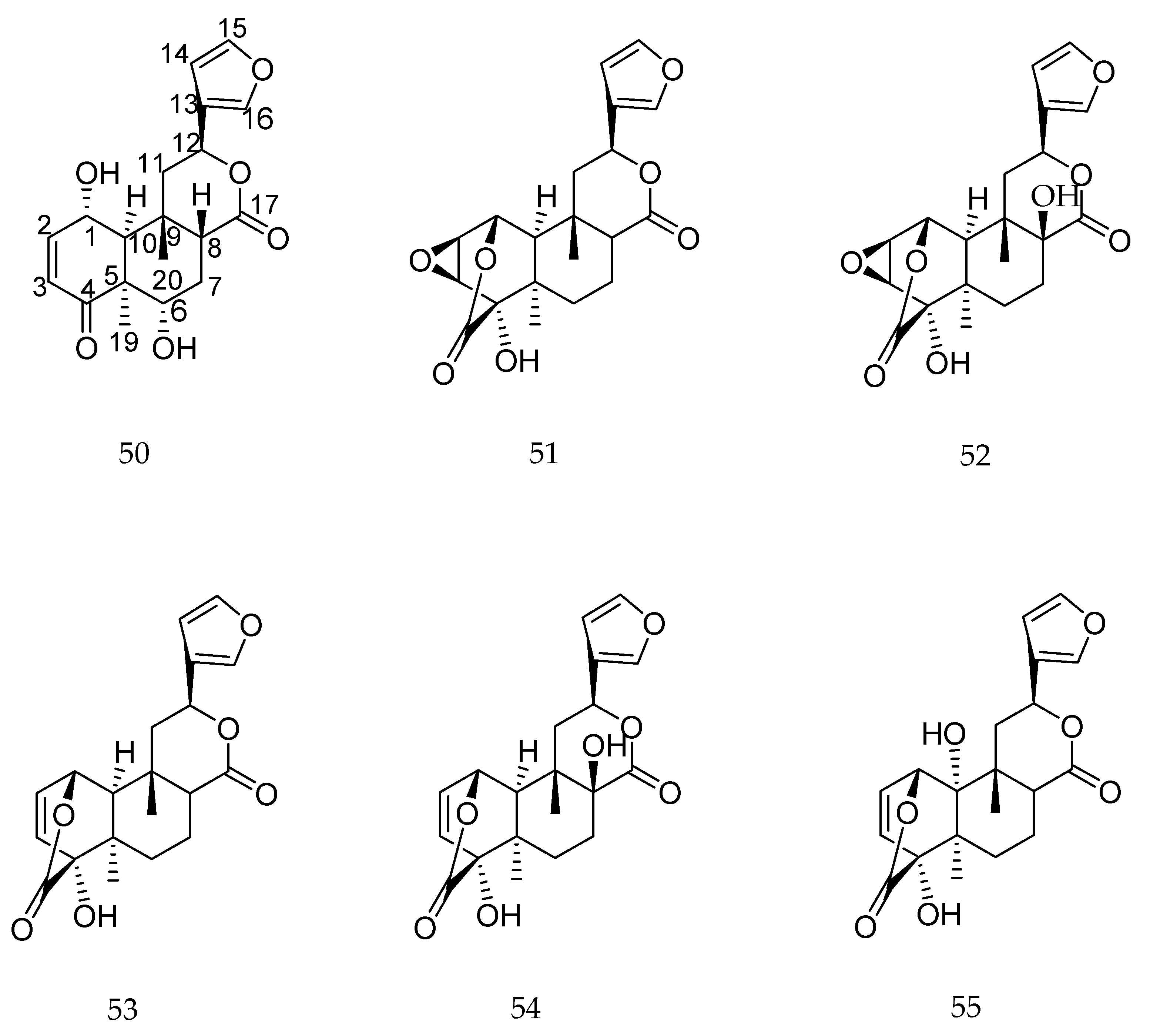
| (+)-(4R,5S,8R,9S)-18-hydroxy-ent-halima-1(10),13-(E)-dien-15-oic | fungi | anticholinesterase (Hr-AChE) | oxidation carboxylation hydroxylation | enhance enhance enhance | [55] |
| cryptotanshinone | fungi | antiviral | Degradation and rearrangement | enhance | [54] |
| 16-oxacleroda-3,13(14)E-dien-15-oic acid | fungi | antifungal | hydroxylation | enhance | [82] |
| trachyloban-19-oic acid | fungi | anticholinesterase | oxidation rearrangement | enhance enhance | [65] |
| 10-oxo-2R,5R,14β-triacetox-ytaxa-4(20),11(12)-diene | bacteria | tumor MDR reversal activity | hydroxylation dihydroxylation oxidation and acetalization | no effect no effect enhance | [59] |
| deoxyandrographolide | fungi | LPS-induced NO production | oxidation of –OH to ketone hydroxylation epoxidation cleavage of the lactone | enhance decrease decrease decrease | [77] |
| andrographolide | fungi | cytotoxicity in vitro | oxidation of –OH to ketone hydration epimerization | decrease decrease enhance | [56,57] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Sousa, I.P.; Sousa Teixeira, M.V.; Jacometti Cardoso Furtado, N.A. An Overview of Biotransformation and Toxicity of Diterpenes. Molecules 2018, 23, 1387. https://doi.org/10.3390/molecules23061387
De Sousa IP, Sousa Teixeira MV, Jacometti Cardoso Furtado NA. An Overview of Biotransformation and Toxicity of Diterpenes. Molecules. 2018; 23(6):1387. https://doi.org/10.3390/molecules23061387
Chicago/Turabian StyleDe Sousa, Ingrid P., Maria V. Sousa Teixeira, and Niege A. Jacometti Cardoso Furtado. 2018. "An Overview of Biotransformation and Toxicity of Diterpenes" Molecules 23, no. 6: 1387. https://doi.org/10.3390/molecules23061387
APA StyleDe Sousa, I. P., Sousa Teixeira, M. V., & Jacometti Cardoso Furtado, N. A. (2018). An Overview of Biotransformation and Toxicity of Diterpenes. Molecules, 23(6), 1387. https://doi.org/10.3390/molecules23061387