
Bioconversion of Corticosterone into Corticosterone-Glucoside by Glucosyltransferase

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
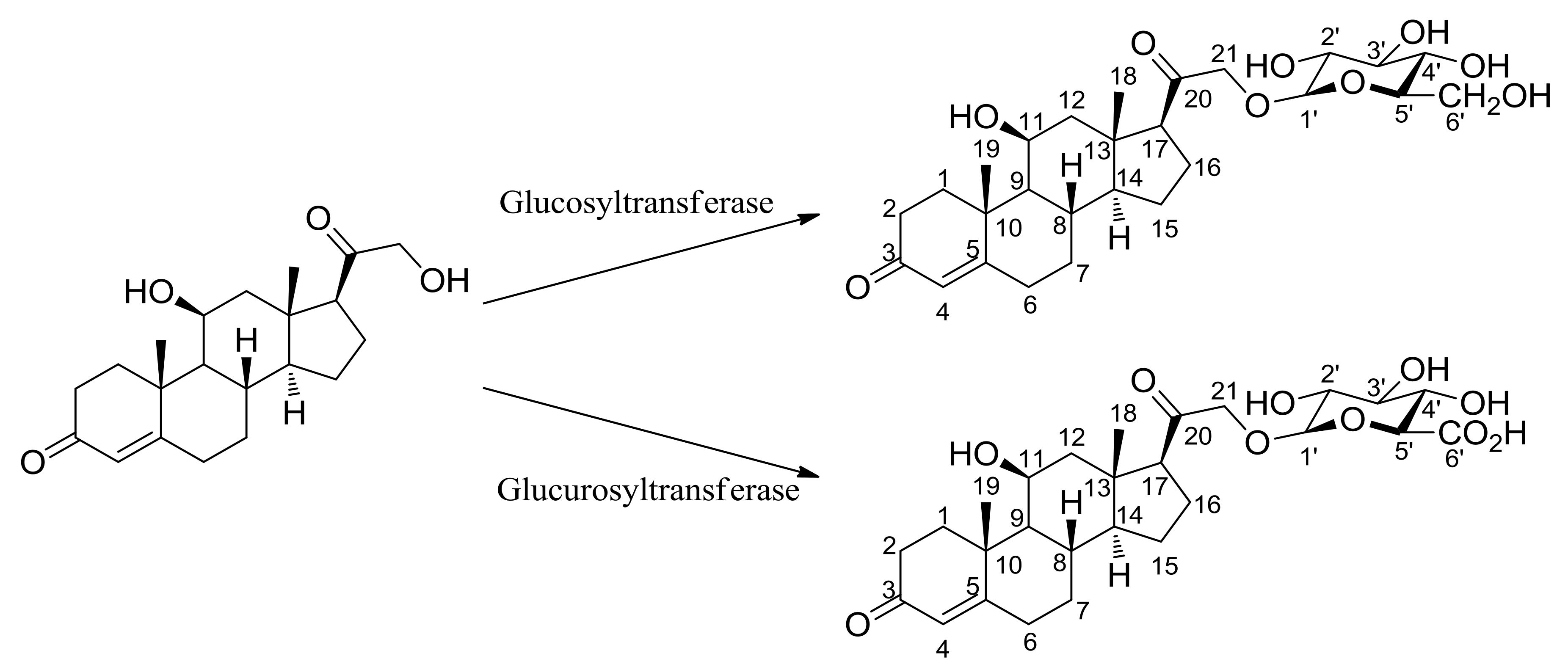
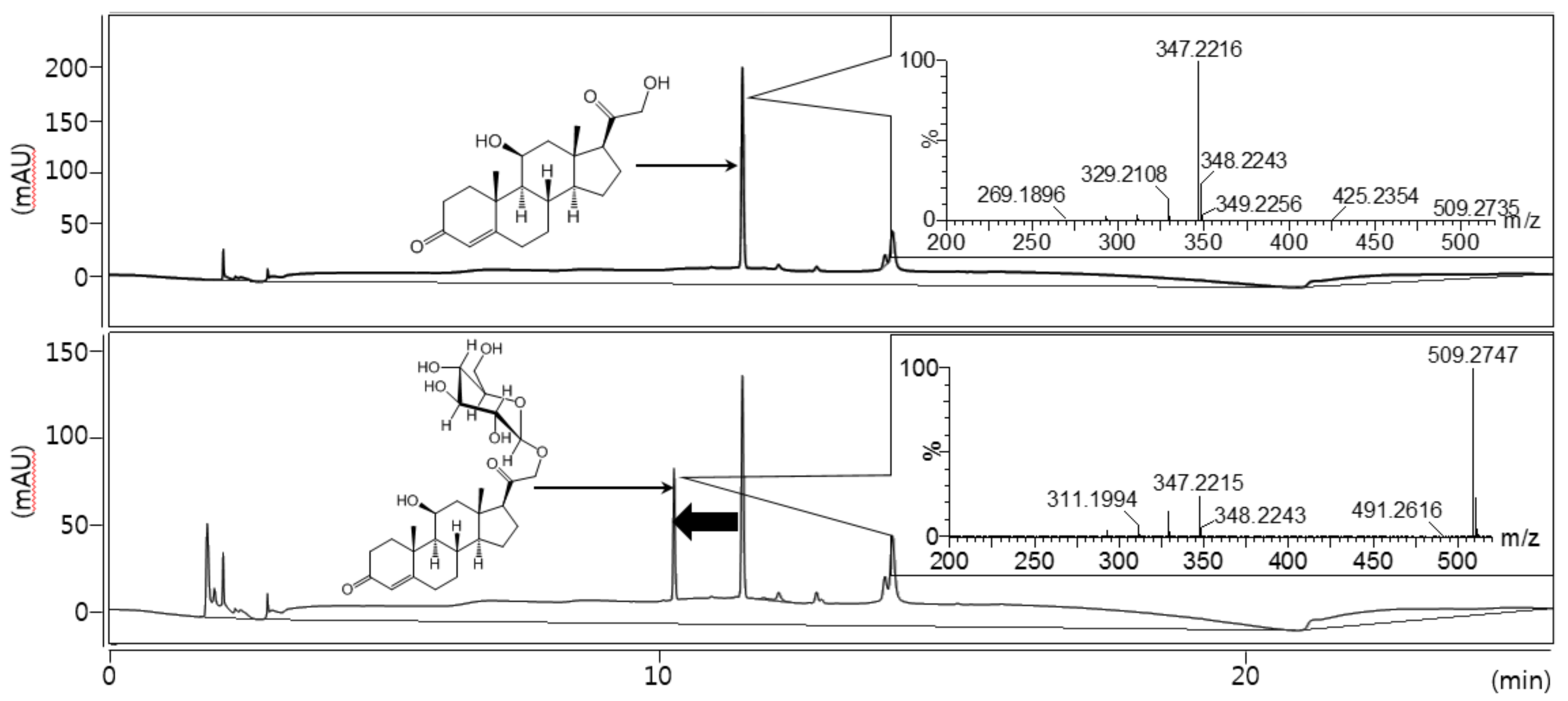
2.1. Bioconversion of Corticosterone 21-Glucoside
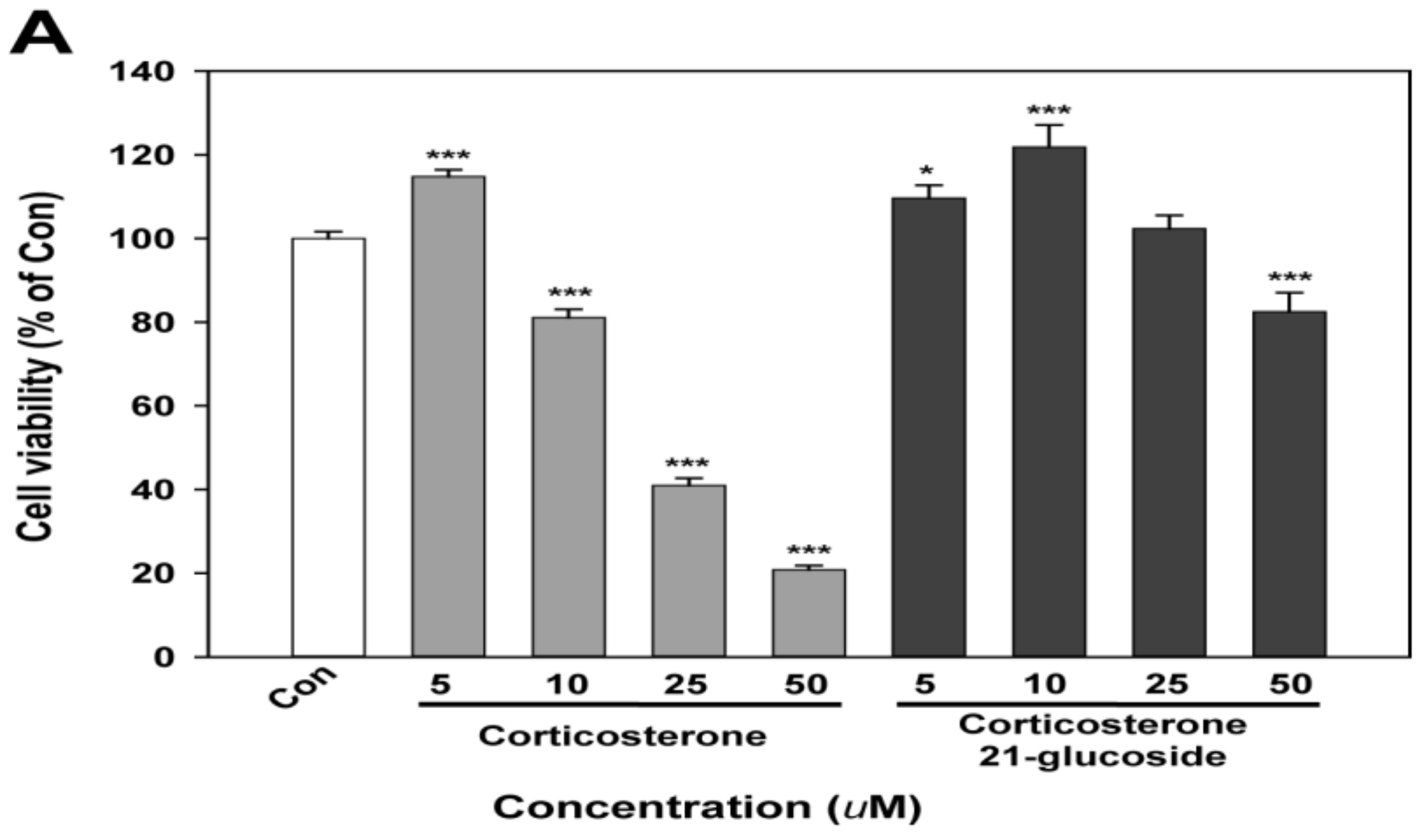
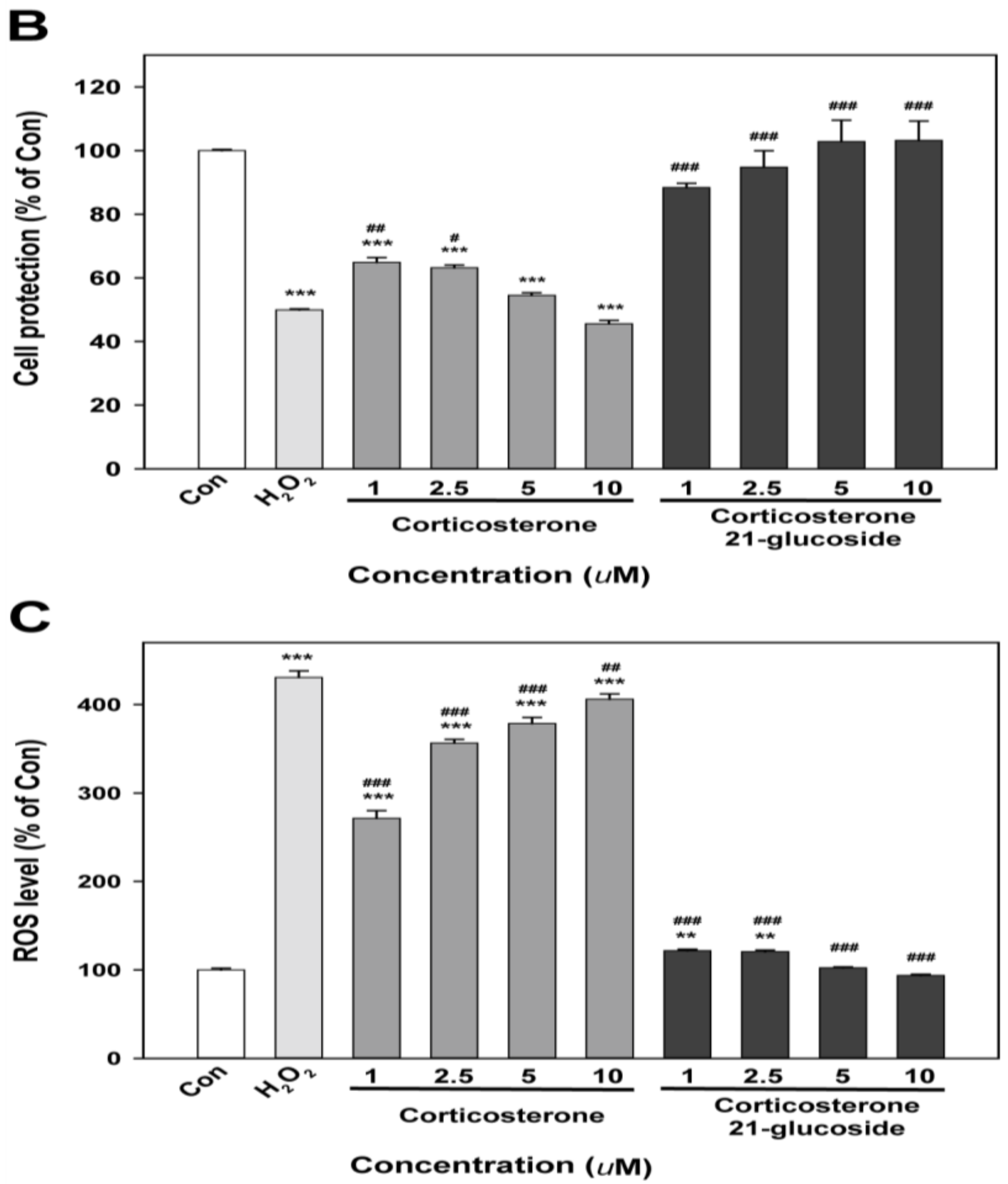
2.2. Bioactivity of Corticosterone 21-Glucoside
3. Materials and Methods
3.1. General Experimental Procedures for Corticosterone 21-Glucoside
3.1.1. Isolation of Corticosterone 21-Glucoside
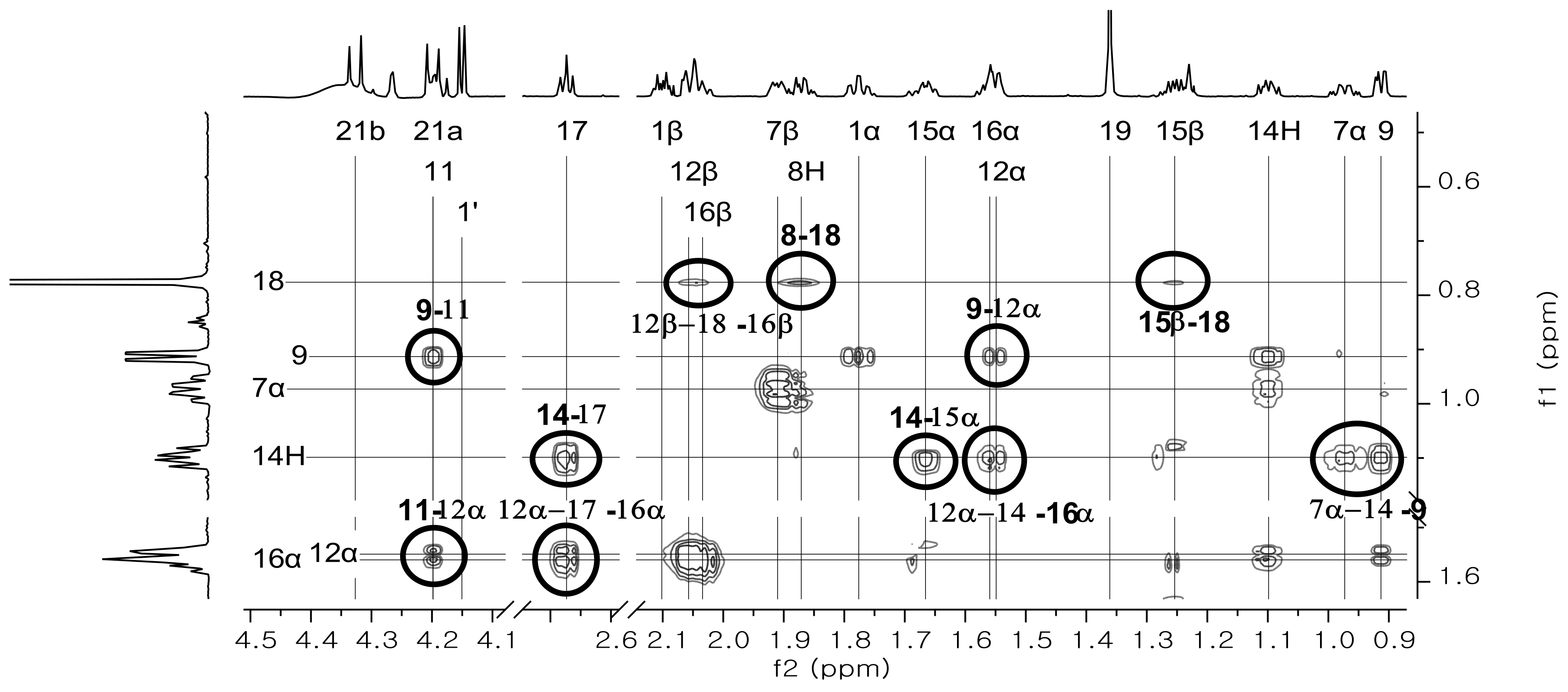
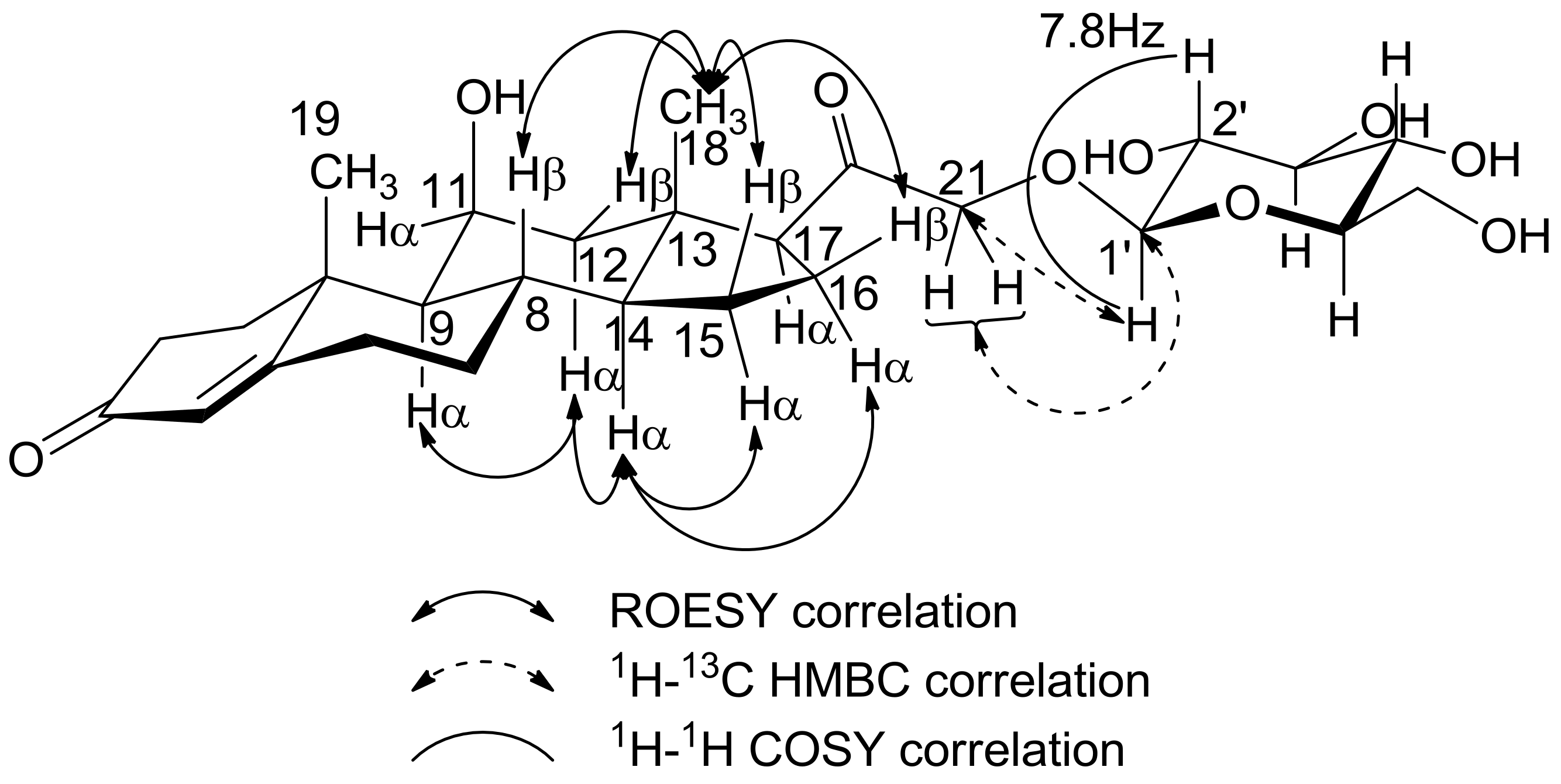
3.1.2. Nuclear Magnetic Resonance and Mass Spectrometry
3.2. Bioconversion Reaction
3.3. Cell Assay for Corticosterone 21-Glucoside
3.3.1. Cell Cytotoxicity and Protection Assay
3.3.2. Reactive Oxygen Species (ROS) Production Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kornel, L.; Saito, Z.; Yuan, L.C. Corticosteroids in human blood-VII. Isolation, characterization and quantitation of glucuronide-conjugated metabolites of cortisol in human plasma. J. Steroid Biochem. 1980, 13, 751–771. [Google Scholar] [CrossRef]
- Daidoji, T.; Inoue, H.; Kato, S.; Yokota, H. Glucuronidation and excretion of nonylphenol in perfused rat liver. Drug Metab. Dispos. 2003, 31, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Yokota, H.; Iwano, H.; Endo, M.; Kobayashi, T.; Inoue, H.; Ikushiro, S. Glucoronidation of the environmental oestrogen bisphenol A by an isoform of UDP-glucoronosyltransferase, UGT2B1, in the rat liver. Biochem. J. 1999, 340, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Slaunwhite, W.R., Jr.; Sandberg, A.A. Transcortin: A corticosteroid-binding protein of plasma. J. Clin. Investig. 1959, 38, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.S. “Bound” to Work: The free hormone hypothesis revisited. Cell 2005, 122, 647–649. [Google Scholar] [CrossRef] [PubMed]
- Hauet, T.; Yao, Z.; Bose, H.S.; Wall, C.T.; Han, Z.; Li, W.; Hales, D.B.; Miller, W.L.; Culty, M.; Papadopoulos, V. Peripheral-type benzodiazepine receptor-mediated action of steroidogenic acute regulatory protein on cholesterol entry into Leydig cell mitochondria. Mol. Endocrinol. 2005, 19, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Steroidogenic acute regulatory protein (StAR), a novel mitochondrial cholesterol transporter. Biochim. Biophys. Acta 2007, 1771, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, V.; Baraldi, M.; Guilarte, T.R.; Knudsen, T.B.; Lacapère, J.J.; Lindemann, P.; Norenberg, M.D.; Nutt, D.; Weizman, A.; Zhang, M.; et al. Translocator protein (18 kDa): New nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function. Trends Pharmacol. Sci. 2006, 27, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. Early steps in androgen biosynthesis: From cholesterol to DHEA. Baillieres. Clin. Endocrinol. MeTab. 1998, 12, 67–81. [Google Scholar] [CrossRef]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef] [PubMed]
- Geissler, W.M.; Davis, D.L.; Wu, L.; Bradshaw, K.D.; Patel, S.; Mendonca, B.B.; Elliston, K.O.; Wilson, J.D.; Russell, D.W. Male pseudohermaphroditism caused by mutations of testicular 17β-hydroxysteroid dehydrogenase 3. Nat. Genet. 1994, 7, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Tanaka, E.; Suzuki, T.; Okumura, K.; Nomura, S.; Miyasho, T.; Haeno, S.; Yokota, H. Accurate determination of tissue steroid hormones, precursors and conjugates in adult male rat. J. Biochem. 2013, 153, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Tahata, S.; Yagi, T.; Tanaka, E.; Masu, K.; Sato, M.; Haeno, S.; Onaga, T.; Yokota, H. Assessment of testicular corticosterone biosynthesis in adult male rats. PLoS ONE 2015, 10, e0117795. [Google Scholar] [CrossRef] [PubMed]
- Goppelt-Struebe, M.; Wolter, D.; Resch, K. Glucocorticoids inhibit prostaglandin synthesis not only at the level of phospholipase A2 but also at the level of cyclo-oxygenase/PGE isomerase. Br. J. Pharmacol. 1989, 98, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Pazirandeh, A.; Xue, Y.; Prestegaard, T.; Jondal, M.; Okret, S. Effects of altered glucocorticoid sensitivity in the T-cell lineage on thymocyte and T-Cell homeostasis. FASEB. J. 2002, 16, 727–729. [Google Scholar] [CrossRef] [PubMed]
- Banuelos, J.; Shin, S.; Cao, Y.; Bochner, B.S.; Morales-Nebreda, L.; Budinger, G.R.; Zhou, L.; Li, S.; Xin, J.; Lingen, M.W.; et al. BCL-2 protects human and mouse Th17 cells from glucocorticoid-induced apoptosis. Allergy 2016, 71, 640–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhen, T.; Cidlowski, J.A. Antiinflammatory action of glucocorticoids-new mechanisms for old drags. N. Engl. J. Med. 2005, 20, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.Y.; Bloom, J.W. Update on glucocorticoid action and resistance. J. Allergy Clin. Immunol. 2003, 111, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, P.; Gomez, F.; King, M.; Lopez, R.; Darby, C.; Schreiber, A.D. In vivo glucocorticoid modulation of guinea pig splenic macrophage Fcγ receptors. J. Clin. Investig. 1991, 88, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Guan, J.; Kang, Y.; Xiu, H.; Chen, Y.; Deng, B.; Liu, K. Inhibition of dehydration-induced water intake by glucocorticoids is associated with activation of hypothalamic natriuretic peptide receptor-A in rat. PLoS ONE 2010, 5, e15607. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, Y.; Kang, Y.; Ni, Z.; Xiu, H.; Guan, J.; Liu, K. Glucocorticoids improve renal responsiveness to atrial natriuretic peptide by up-regulating natriuretic peptide receptor-A expression in the renal inner medullary collecting duct in decompensated heart failure. J. Pharmacol. Exp. Ther. 2011, 339, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Niedziela, M.; Sippell, W. Growth impairment in a boy with late-onset congenital adrenal hyperplasia and anorexia nervosa. Exp. Clin. Endocrinol. Diabetes 2010, 118, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Sapolsky, R.M. Glucocorticoids, stress and exacerbation of excitotoxic neuron death. Neurosciences 1994, 6, 323–331. [Google Scholar] [CrossRef]
- Cahill, L.; McGaugh, J.L. Mechanisms of emotional arousal and lasting declarative memory. Trends Neurosci. 1998, 21, 294–297. [Google Scholar] [CrossRef]
- Belanoff, J.K.; Gross, K.; Yager, A.; Schatzberg, A.F. Corticosteroids and cognition. J. Psychiatr. Res. 2001, 35, 127–145. [Google Scholar] [CrossRef]
- De Quervain, D.J.; Roozendaal, B.; McGaugh, J.L. Stress and glucocorticoids impair retrieval of long-term spatial memory. Nature 1998, 394, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; Maheu, F.; Tu, M.; Fiocco, A.; Schramek, T.E. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn. 2007, 65, 209–237. [Google Scholar] [CrossRef] [PubMed]
- Ennis, G.E.; Moffat, S.D.; Hertzog, C. The cortisol awakening response and cognition across the adult lifespan. Brain Cogn. 2016, 105, 66–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Dykens, J.A.; Perez, E.; Liu, R.; Yang, S.; Covey, D.F.; Simpkins, J.W. Neuroprotective effects of 17beta-estradiol and nonfeminizing estrogens against H2O2 toxicity in human neuroblastoma SK-N-SH cells. Mol. Pharmacol. 2006, 70, 295–404. [Google Scholar]
- Zhan, Y.; Ma, R.; Shen, J.; Su, H.; Xing, D.; Du, L. A mouse model of depression induced by repeated corticosterone injection. Eur. J. Pharmacol. 2008, 581, 113–120. [Google Scholar]
- Kalynchuk, L.E.; Gregus, A.; Boudreau, D.; Perrot-Sinal, T.S. Corticosterone increases depression-like behavior, with some effects on predator odor-induced defensive behavior, in male and female rats. Behav. Neurosci. 2004, 118, 1365–1377. [Google Scholar] [CrossRef] [PubMed]
- Jäntti, S.E.; Kiriazis, S.; Reinilä, R.R.; Kostiainen, R.K.; Ketola, R.A. Enzyme-assisted synthesis and characterization of glucuronide conjugates of neuroactive steroids. Steroids 2007, 72, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Stachulski, A.V.; Jenkins, G.N. The synthesis of O-glucuronides. Nat. Prod. Rep. 1998, 15, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Mattox, V.R.; Goodrich, J.E.; Vrieze, W.D. Synthesis of C-21 glucosiduronates of cortisone and related corticosteroids. Biochemistry 1969, 8, 1188–1199. [Google Scholar] [CrossRef] [PubMed]
- Ciuffreda, P.; Casati, S.; De Mieri, M.; Ferrabosch, P. Corticosteroids 21-glucuronides: Synthesis and complete characterization by 1H and 13C NMR. Steroids 2009, 74, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Kirk, D.N.; Toms, H.C.; Douglas, C.; White, K.A. A Survey of the High-field 1H NMR Spectra of the Steroid Hormones, their Hydroxylated Derivatives, and Related Compounds. J. Chem. Soc. Perkin Trans. 1990, 2, 1957–1594. [Google Scholar]
- Ben-Menachem, G.; Kubler-Kielb, J.; Coxon, B.; Yergey, A.; Schneerson, R. A newly discovered cholesteryl galactoside from Borrelia burgdorferi. Proc. Natl. Acad. Sci. USA 2003, 100, 7913–7918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaki, T.; Zähringer, U.; Warnecke, D.C.; Fahl, A.; Knogge, W.; Heinz, E. Sterol glycosides and cerebrosides accumulate in Pichia pastoris, Rhynchosporium secalis and other fungi under normal conditions or under heat shock and ethanol stress. Yeast 2001, 18, 679–695. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.F. Biosynthesis of cholesteryl glucoside by Mycoplasma gallinarum. J. Bacteriol. 1971, 108, 986–991. [Google Scholar] [PubMed]
- Warnecke, D.; Erdmann, R.; Fahl, A.; Hube, B.; Müller, F.; Zank, T.; Zähringer, U.; Heinz, E. Cloning and functional expression of UGT genes encoding sterol glucosyltransferases from Saccharomyces cerevisiae, Candida albicans, Pichia pastoris, and Dictyostelium discoideum. J. Biol. Chem. 1999, 274, 13048–13059. [Google Scholar] [CrossRef] [PubMed]
- Warnecke, D.C.; Baltrusch, M.; Buck, F.; Wolter, F.P.; Heinz, E. UDP-glucose:sterol glucosyltransferase: Cloning and functional expression in Escherichia coli. Plant Mol. Biol. 1997, 35, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Schulze, E.; Weiss, E.; Weiss, F.; Böhme, H.J.; Hofmann, E. An improved method for adrenalectomy of suckling rats. The influence of thrombin treatment and deoxycorticosterone substitution on survival and on hepatic and renal enzyme activities. Biomed. Biochim. Acta 1986, 45, 1049–1056. [Google Scholar] [PubMed]
- Csaba, G.; Lantos, T.; Nagy, S.U.; Arányi, P.; Náray, A. Effects of steroids on Tetrahymena. Acta Biol. Med. Ger. 1978, 37, 1377–1380. [Google Scholar] [PubMed]
- Chou, F.P.; Tsai, C.T.; Chiou, Y.S.; Chen, Y.J.; Li, M.E.; Guo, T.W.; Lyu, J.W.; Chou, S.H.; Wu, T.K. An enzymatic approach to configurationally rare trans-androsteronyl-α-glucoside and its potential anticancer application. Chem. Biol. Drug Des. 2017, 89, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.G.; Gwilliam, C.; Layne, D.S. Steroid β-d-glucosidase in steer liver and kidney. Can. J. Biochem. 1977, 55, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Kuro, T.; Konishi, F.; Takaoka, M.; Gariepy, C.E.; Yanagisawa, M. Enhanced blood pressure sensitivity to DOCA-salt treatment in endothelin ET(B) receptor-deficient rats. Br. J. Pharmacol. 2000, 129, 1060–1062. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.; Chan, V.; Brown, L. The DOCA-Salt Hypertensive Rat as a Model of Cardiovascular Oxidative and Inflammatory Stress. Curr. Cardiol. Rev. 2010, 6, 291–297. [Google Scholar] [CrossRef] [PubMed]
- De Pádua, R.M.; Meitinger, N.; de Souza Filho, J.D.; Waibel, R.; Gmeiner, P.; Braga, F.C.; Kreis, W. Biotransformation of 21-O-acetyl-deoxycorticosterone by cell suspension cultures of Digitalis lanata (strain W.1.4). Steroids 2012, 77, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- IUPAC Commission on the Nomenclature of Organic Chemistry (CNOC); IUPAC-IUB Commission on Biochemical Nomenclature (CBN). The nomenclature of steroids. Revised tentative rules. Eur. J. Biochem. 1969, 10, 1–19. [Google Scholar]
- Mehler, M.F.; Gokhan, S. Mechanisms underlying neural cell death in neurodegenerative diseases: Alterations of a developmentally-mediated cellular rheostat. Trends Neurosci. 2000, 23, 599–605. [Google Scholar] [CrossRef]
- Facecchia, K.; Fochesato, L.A.; Ray, S.D.; Stohs, S.J.; Pandey, S. Oxidative toxicity in neurodegenerative diseases: Role of mitochondrial dysfunction and therapeutic strategies. J. Toxicol. 2011, 2011, 683728. [Google Scholar] [CrossRef] [PubMed]
- Im, A.-R.; Kim, Y.-H.; Uddin, M.R.; Chae, S.; Lee, H.W.; Kim, Y.H.; Kim, Y.S.; Lee, M.-Y. Betaine protects against rotenone-induced neurotoxicity in PC12 cells. Cell. Mol. Neurobiol. 2013, 33, 625–635. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds corticosterone 21-glucoside are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 13C | 1H with Coupling Constants JHH1 | ||||||
|---|---|---|---|---|---|---|---|
| 21-Glucuronide 2 | 21-Glucoside 3 | 21-Glucuronide 2 | 21-Glucoside 3 | ||||
| δ (ppm) | δ (ppm) | δ (ppm) | JHH (Hz) | δ (ppm) | JHH (Hz) | ||
| Aglycone moiety | Aglycone moiety | ||||||
| 1 | 34.45 | 34.04 | 1α | 1.77 | 13.2, 4.0 | 1.78 | ddd, 13.6, 13.6, 4.4 |
| 1β | 2.10 | 3.4, 3.1 | 2.10 | ddd, 13.3, 4.7, 4.7 | |||
| 2 | 33.93 | 33.48 | 2α | 2.19 | 4.0, 3.4 | 2.18 | ddd, 16.8, 4.0, 4.0 |
| 2β | 2.39 | 13.2, 3.1 | 2.38 | ddd, 16.5, 13.8, 5.0 | |||
| 3 | 198.52 | 198.1 | |||||
| 4 | 121.97 | 121.52 | 4 | 5.63 | 5.56 | d, 1.7 | |
| 5 | 172.73 | 172.32 | |||||
| 6 | 31.79 | 31.35 | 6α | 2.17 | 13.0, 4.3, 2.1 | 2.18 | ddd, 16.8, 4.0, 4.0 |
| 6β | 2.44 | 13.2, 13.0. 5.0 | 2.44 | dddd, 14.2, 14.2, 5.6, 1.6 | |||
| 7 | 32.95 | 32.52 | 7α | 0.98 | 13.2, 12.6, 11.2, 4.3 | 0.97 | dddd, 14.6, 12.6, 11.2, 4.6 |
| 7β | 1.87 | 12.6, 5.0, 3.5, 2.1 | 1.91 | dddd, 12.2, 5.8, 4.1, 2.2 | |||
| 8 | 31.55 | 31.13 | 8 | 1.92 | 12.2, 11.2, 11.2, 3.5 | 1.87 | dddd, 11.2, 11.2, 11.1, 4.1 |
| 9 | 55.95 | 55.46 | 9 | 0.91 | 12.2, 4.1 | 0.91 | dd, 11.2, 3.4 |
| 10 | 39.44 | 38.86 | |||||
| 11 | 66.67 | 66.14 | 11α | 4.21 | 4.1, 4.1, 2.7 | 4.20 | dddd, 9.7, 3.3, 3.2, 3.2 |
| 11β | - | - | |||||
| 12 | 46.96 | 46.52 | 12α | 2.02 | 12.7, 4.1 | 1.57–1.53 | m |
| 12β | 1.54 | 12.7, 2.7 | 2.07–2.04 | m | |||
| 13 | 43.82 | 43.41 | |||||
| 14 | 57.33 | 56.91 | 14 | 1.09 | 11.2, 6.3 | 1.10 | ddd, 12.5, 10.6, 7.1 |
| 15 | 24.54 | 24.13 | 15α | 1.66 | 12.4, 12.4, 6.3, 2.2 | 1.67 | dddd, 12.1, 9.7, 7.1, 2.9 |
| 15β | 1.25 | 12.4, 11.2, 11.2, 5.8 | 1.25 | dddd, 12.1, 12.1, 12.0, 6.6 | |||
| 16 | 22.29 | 21.8 | 16α | 1.57 | 13.6, 12.4, 8.4, 5.8 | 1.59–1.53 | m |
| 16β | 2.04 | 13.6, 11.2, 8.4, 2.2 | 2.03 | ddd, 13.5, 9.3, 2.9 | |||
| 17 | 58.51 | 58.06 | 17 | 2.69 | 8.4, 8.4 | 2.67 | t, 9.2 |
| 18 | 16.17 | 15.76 | 18-CH3 | 0.79 | 0.78 | s | |
| 19 | 20.82 | 20.37 | 19-CH3 | 1.37 | 1.36 | s | |
| 20 | 207.79 | 207.61 | |||||
| 21 | 74.56 | 73.44 | 21 | 4.29 | 18.2 | 4.33 | d, 17.3 |
| 21 | 4.16 | 18.2 | 4.20 | d, 17.3 | |||
| Sugar moiety | Sugar moiety | ||||||
| 1′ | 103.2 | 102.14 | 1′ | 4.28 | 7.8 | 4.15 | d, 7.8 |
| 2′ | 73.48 | 73.30 | 2′ | 3.07 | 8.6, 7.8 | 3.00 | dd, 8.4, 8.4 |
| 3′ | 76.36 | 76.55 | 3′ | 3.18 | 8.6, 8.0 | 3.12 | dd, 8.9, 8.9 |
| 4′ | 71.85 | 69.97 | 4′ | 3.33 | 9.5, 8.0 | 3.02 | dd, 9.3, 9.3 |
| 5′ | 76.17 | 77.03 | 5′ | 3.63 | 9.5 | 3.08 | ddd, 9.9, 6.1, 2.1 |
| 6′ | 170.71 | 61.09 | 6′ | 3.42 | dd, 11.6, 6.1 | ||
| 6′ | 3.66 | dd, 11.7, 2.1 | |||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamaguchi, T.; Lee, J.-H.; Lim, A.-R.; Sim, J.-S.; Yu, E.-J.; Oh, T.-J. Bioconversion of Corticosterone into Corticosterone-Glucoside by Glucosyltransferase. Molecules 2018, 23, 1783. https://doi.org/10.3390/molecules23071783
Yamaguchi T, Lee J-H, Lim A-R, Sim J-S, Yu E-J, Oh T-J. Bioconversion of Corticosterone into Corticosterone-Glucoside by Glucosyltransferase. Molecules. 2018; 23(7):1783. https://doi.org/10.3390/molecules23071783
Chicago/Turabian StyleYamaguchi, Tokutaro, Joo-Ho Lee, A-Rang Lim, Joon-Soo Sim, Eun-Ji Yu, and Tae-Jin Oh. 2018. "Bioconversion of Corticosterone into Corticosterone-Glucoside by Glucosyltransferase" Molecules 23, no. 7: 1783. https://doi.org/10.3390/molecules23071783
APA StyleYamaguchi, T., Lee, J.-H., Lim, A.-R., Sim, J.-S., Yu, E.-J., & Oh, T.-J. (2018). Bioconversion of Corticosterone into Corticosterone-Glucoside by Glucosyltransferase. Molecules, 23(7), 1783. https://doi.org/10.3390/molecules23071783