Spectroscopic Characterization of Natural Melanin from a Streptomyces cyaneofuscatus Strain and Comparison with Melanin Enzymatically Synthesized by Tyrosinase and Laccase




 and
and 
Abstract
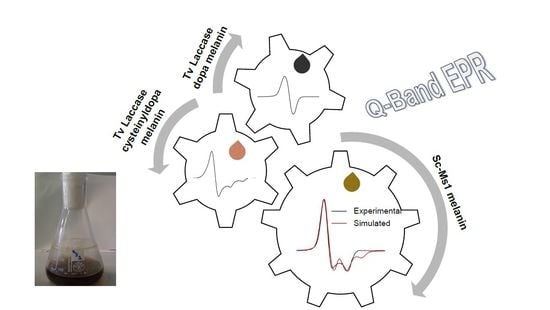
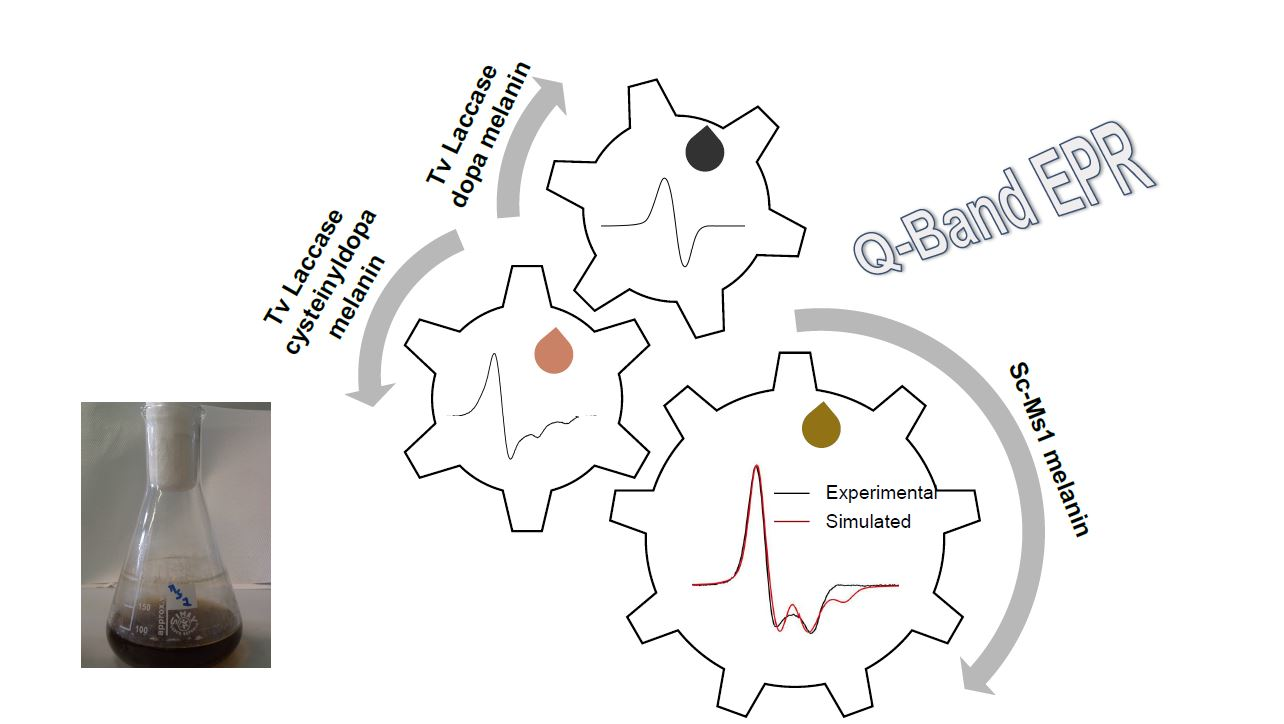
:
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Sc-Ms1 Tyrosinase Purification
3.2. Melanin Isolation and Purification
3.3. Melanin-Like Pigments Synthesis by Sc-Ms1 Tyrosinase and Tv Laccase
3.4. UV-Visible and FT-IR Spectroscopies
3.5. Electron Paramagnetic Resonance Spectroscopy
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Solano, F. Melanin and melanin-related polymers as materials with biomedical and biotechnological applications—Cuttlefish ink and mussel foot proteins as inspired biomolecules. Int. J. Mol. Sci. 2017, 18, 1561. [Google Scholar] [CrossRef] [PubMed]
- Meredith, P.; Sarna, T. The physical and chemical properties of eumelanin. Pigment Cell Res. 2006, 19, 572–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ischia, M.; Napolitano, A.; Pezzella, A.; Meredith, P.; Sarna, T. Chemical and structural diversity in eumelanins: Unexplored bio-optoelectronic materials. Angew. Chem. Int. Ed. 2009, 48, 3914–3921. [Google Scholar] [CrossRef] [PubMed]
- Felix, C.C.; Hyde, J.S.; Sarna, T.; Sealy, R. Interactions of Melanin with Metal Ions. Electron Spin Resonance Evidence for Chelate Complexes of Metal Ions with Free Radicals. J. Am. Chem. Soc. 1978, 100, 3922–3926. [Google Scholar] [CrossRef]
- Solano, F. Melanins: Skin Pigments and Much More—Types, Structural Models, Biological Functions, and Formation Routes. New J. Sci. 2014, 2014, 498276. [Google Scholar] [CrossRef]
- D’Ischia, M.; Wakamatsu, K.; Napolitano, A.; Briganti, S.; Garcia-Borron, J.C.; Kovacs, D.; Meredith, P.; Pezzella, A.; Picardo, M.; Sarna, T.; et al. Melanins and melanogenesis: Methods, standards, protocols. Pigment Cell Melanoma Res. 2013, 26, 616–633. [Google Scholar] [CrossRef] [PubMed]
- Iacomino, M.; Mancebo-Aracil, J.; Guardingo, M.; Martín, R.; D’Errico, G.; Perfetti, M.; Manini, P.; Crescenzi, O.; Busqué, F.; Napolitano, A.; et al. Replacing nitrogen by sulfur: From structurally disordered eumelanins to regioregular thiomelanin polymers. Int. J. Mol. Sci. 2017, 18, 2169. [Google Scholar] [CrossRef] [PubMed]
- Mostert, A.B.; Hanson, G.R.; Sarna, T.; Gentle, I.R.; Powell, B.J.; Meredith, P. Hydration-controlled X-band EPR spectroscopy: A tool for unravelling the complexities of the solid-state free radical in eumelanin. J. Phys. Chem. B 2013, 117, 4965–4972. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.T.; Chuang, C.; Cao, J.; Ball, V.; Ruch, D.; Buehler, M.J. Excitonic effects from geometric order and disorder explain broadband optical absorption in eumelanin. Nat. Commun. 2014, 5, 3859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Ischia, M.; Napolitano, A.; Ball, V.; Chen, C.T.; Buehler, M.J. Polydopamine and eumelanin: From structure-property relationships to a unified tailoring strategy. Acc. Chem. Res. 2014, 47, 3541–3550. [Google Scholar] [CrossRef] [PubMed]
- Arzillo, M.; Mangiapia, G.; Pezzella, A.; Heenan, R.K.; Radulescu, A.; Paduano, L.; D’Ischia, M. Eumelanin buildup on the nanoscale: Aggregate growth/assembly and visible absorption development in biomimetic 5,6-dihydroxyindole polymerization. Biomacromolecules 2012, 13, 2379–2390. [Google Scholar] [CrossRef] [PubMed]
- Drewnowska, J.M.; Zambrzycka, M.; Kalska-Szostko, B.; Fiedoruk, K.; Swiecicka, I. Melanin-like pigment synthesis by soil bacillus weihenstephanensis isolates from Northeastern Poland. PLoS ONE 2015, 10, e0125428. [Google Scholar] [CrossRef] [PubMed]
- Harir, M.; Bellahcene, M.; Baratto, M.C.; Pollini, S.; Rossolini, G.M.; Trabalzini, L.; Fatarella, E.; Pogni, R. Isolation and characterization of a novel tyrosinase produced by Sahara soil actinobacteria and immobilization on nylon nanofiber membranes. J. Biotechnol. 2018, 265, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Le Roes-Hill, M.; Prins, A.; Meyers, P.R. Streptomyces swartbergensis sp. nov., a novel tyrosinase and antibiotic producing actinobacterium. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2018, 111, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Claus, H.; Decker, H. Bacterial tyrosinases. Syst. Appl. Microbiol. 2006, 29, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Ito, S. IFPCS Presidential Lecture A Chemist’s View of Melanogenesis. Pigment Cell Res. 2003, 16, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M.; Grabacka, M. Melanin synthesis in microorganisms—Biotechnological and medical aspects. Acta Biochim. Pol. 2006, 53, 429–443. [Google Scholar]
- Varga, M.; Berkesi, O.; Darula, Z.; May, N.V.; Palágyi, A. Structural characterization of allomelanin from black oat. Phytochemistry 2016, 130, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida-Paes, R.; Frases, S.; de Sousa Araújo, G.; Evangelista de Oliveira, M.M.; Gerfen, G.J.; Nosanchuk, J.D.; Zancopé-Oliveira, R.M. Biosynthesis and functions of a melanoid pigment produced by species of the sporothrix complex in the presence of L-Tyrosine. Appl. Environ. Microbiol. 2012, 78, 8623–8630. [Google Scholar] [CrossRef] [PubMed]
- Schmaler-Ripcke, J.; Sugareva, V.; Gebhardt, P.; Winkler, R.; Kniemeyer, O.; Heinekamp, T.; Brakhage, A.A. Production of pyomelanin, a second type of melanin, via the tyrosine degradation pathway in Aspergillus fumigatus. Appl. Environ. Microbiol. 2009, 75, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Ivins, B.E.; Holmes, R.K. Factors affecting phaeomelanin production by a melanin-producing (mel) mutant of Vibrio cholerae. Infect. Immun. 1981, 34, 895–899. [Google Scholar] [PubMed]
- Li, C.; Ji, C.; Tang, B. Purification, characterization and biological activity of melanin from Streptomyces sp. FEMS Microbiol. Lett. 2018, fny077. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Guo, G. yi; Lu, Y.; Song, S.; Wang, H. yan; Yang, L. Purification, structure and anti-radiation activity of melanin from Lachnum YM404. Int. J. Biol. Macromol. 2014, 63, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Zhang, X.; Sun, S.; Zhang, L.; Shan, S.; Zhu, H. Production of natural melanin by Auricularia auricula and study on its molecular structure. Food Chem. 2016, 190, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Buszman, E.; Pilawa, B.; Zdybel, M.; Wilczyński, S.; Gondzik, A.; Witoszyńska, T.; Wilczok, T. EPR examination of Zn2+ and Cu2+ binding by pigmented soil fungi Cladosporium cladosporioides. Sci. Total Environ. 2006, 363, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Faccio, G.; Kruus, K.; Saloheimo, M.; Thöny-meyer, L. Bacterial tyrosinases and their applications. Process Biochem. 2012, 47, 1749–1760. [Google Scholar] [CrossRef]
- Selinheimo, E.; Nieidhin, D.; Steffensen, C.; Nielsen, J.; Lomascolo, A.; Halaouli, S.; Record, E.; Beirne, D.O.; Buchert, J.; Kruus, K. Comparison of the characteristics of fungal and plant tyrosinases. J. Biotechnol. 2007, 130, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M. Electron paramagnetic resonance as a unique tool for skin and hair research. Exp. Dermatol. 2009, 18, 472–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdybel, M.; Pilawa, B.; Drewnowska, J.M.; Swiecicka, I. Comparative EPR studies of free radicals in melanin synthesized by Bacillus weihenstephanensis soil strains. Chem. Phys. Lett. 2017, 679, 185–192. [Google Scholar] [CrossRef]
- Sealy, R.C.; Hyde, J.S.; Felix, C.C.; Menon, I.A.; Prota, G. Eumelanins and pheomelanins: characterization by electron spin resonance spectroscopy. Science (80-) 1982, 217, 545–547. [Google Scholar] [CrossRef]
- Sealy, R.C.; Hyde, J.S.; Felix, C.C.; Menont, I.A.; Prota, G.; Swartz, H.M.; Persadt, S.; Habermant, H.F. Novel free radicals in synthetic and natural pheomelanins: Distinction between dopa melanins and cysteinyldopa melanins by ESR spectroscopy. Proc. Natl. Acad. Sci. USA 1982, 79, 2885–2889. [Google Scholar] [CrossRef] [PubMed]
- Hannson, C.; Agrup, G.; Rorsman, H.; Rosengren, A.M.; Rosengren, E. Electron Spin Resonance Studies on Pheomelanins. Acta Dermatovener 1979, 59, 453–456. [Google Scholar]
- Chikvaidze, E.N.; Partskhaladze, T.M.; Gogoladze, T.V. Electron spin resonance (ESR/EPR) of free radicals observed in human red hair: A new, simple empirical method of determination of pheomelanin/eumelanin ratio in hair. Magn. Reson. Chem. 2014, 52, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Selinheimo, E.; Gasparetti, C.; Mattinen, M.; Steffensen, C.L.; Buchert, J.; Kruus, K. Comparison of substrate specificity of tyrosinases from Trichoderma reesei and Agaricus bisporus. Enzyme Microb. Technol. 2009, 44, 1–10. [Google Scholar] [CrossRef]
- Hernandez-Romero, D.; Sanchez-amat, A.; Solano, F. A tyrosinase with an abnormally high tyrosine hydroxylase/dopa oxidase ratio Role of the seventh histidine and accessibility to the active site. FEBS J. 2006, 273, 257–270. [Google Scholar] [CrossRef] [PubMed]
- Commoner, B.; Townsend, J.; Pake, G.E. Free Radicals in biological materials. Nature 1954, 4432, 689–691. [Google Scholar] [CrossRef]
- Pasenkiewicz-Gierula, M.; Sealy, R.C. Analysis of the ESR spectrum of synthetic dopa melanin. Biochim. Biophys. Acta 1986, 884, 510–516. [Google Scholar] [CrossRef]
- Brogioni, B.; Biglino, D.; Sinicropi, A.; Reijerse, E.J.; Giardina, P.; Sannia, G.; Lubitz, W.; Basosi, R.; Pogni, R. Characterization of radical intermediates in laccase-mediator systems. A multifrequency EPR, ENDOR and DFT/PCM investigation. Phys. Chem. Chem. Phys. 2008, 10, 7284–7292. [Google Scholar] [CrossRef] [PubMed]
- Pogni, R.; Baratto, M.C.; Sinicropi, A.; Basosi, R. Spectroscopic and computational characterization of laccases and their substrate radical intermediates. Cell. Mol. Life Sci. 2015, 72, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; De Lucia, M.; Panzella, L.; D’Ischia, M. The “benzothiazine” chromophore of pheomelanins: A reassessment. Photochem. Photobiol. 2008, 84, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Di Donato, P.; Prota, G. New regulatory mechanisms in the biosynthesis of pheomelanins: Rearrangement vs. redox exchange reaction routes of a transient 2H-1,4-benzothiazine-o-quinonimine intermediate. Biochim. Biophys. Acta–Gen. Subj. 2000, 1475, 47–54. [Google Scholar] [CrossRef]
- Stoll, S.; Schweiger, A. EasySpin, a comprehensive software package for spectral simulation and analysis in EPR. J. Magn. Reson. 2006, 178, 42–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the Sc-Ms1 natural melanin and enzymic synthesized melanins are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ANiso | ANz | 2ANz | giso * | gz | gx = gy | ΔBpp § (mT) |
|---|---|---|---|---|---|---|---|
| Sc-Ms1 melanin | 2.0047 | 0.7 | |||||
| Sc-Ms1 Tyr. dopa melanin | 2.0038 | 0.6 | |||||
| Tv Lac. dopa melanin | 2.0036 | 0.5 | |||||
| Tv Lac. cysteinyldopa melanin | 0.7 | 1.6 | 3.2 | 2.0050 | 2.0028 | 2.0060 | 3.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Khatib, M.; Harir, M.; Costa, J.; Baratto, M.C.; Schiavo, I.; Trabalzini, L.; Pollini, S.; Rossolini, G.M.; Basosi, R.; Pogni, R. Spectroscopic Characterization of Natural Melanin from a Streptomyces cyaneofuscatus Strain and Comparison with Melanin Enzymatically Synthesized by Tyrosinase and Laccase. Molecules 2018, 23, 1916. https://doi.org/10.3390/molecules23081916
Al Khatib M, Harir M, Costa J, Baratto MC, Schiavo I, Trabalzini L, Pollini S, Rossolini GM, Basosi R, Pogni R. Spectroscopic Characterization of Natural Melanin from a Streptomyces cyaneofuscatus Strain and Comparison with Melanin Enzymatically Synthesized by Tyrosinase and Laccase. Molecules. 2018; 23(8):1916. https://doi.org/10.3390/molecules23081916
Chicago/Turabian StyleAl Khatib, Maher, Mohamed Harir, Jessica Costa, Maria Camilla Baratto, Irene Schiavo, Lorenza Trabalzini, Simona Pollini, Gian Maria Rossolini, Riccardo Basosi, and Rebecca Pogni. 2018. "Spectroscopic Characterization of Natural Melanin from a Streptomyces cyaneofuscatus Strain and Comparison with Melanin Enzymatically Synthesized by Tyrosinase and Laccase" Molecules 23, no. 8: 1916. https://doi.org/10.3390/molecules23081916
APA StyleAl Khatib, M., Harir, M., Costa, J., Baratto, M. C., Schiavo, I., Trabalzini, L., Pollini, S., Rossolini, G. M., Basosi, R., & Pogni, R. (2018). Spectroscopic Characterization of Natural Melanin from a Streptomyces cyaneofuscatus Strain and Comparison with Melanin Enzymatically Synthesized by Tyrosinase and Laccase. Molecules, 23(8), 1916. https://doi.org/10.3390/molecules23081916