Biopesticide Activity from Drimanic Compounds to Control Tomato Pathogens



Abstract
:1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. General
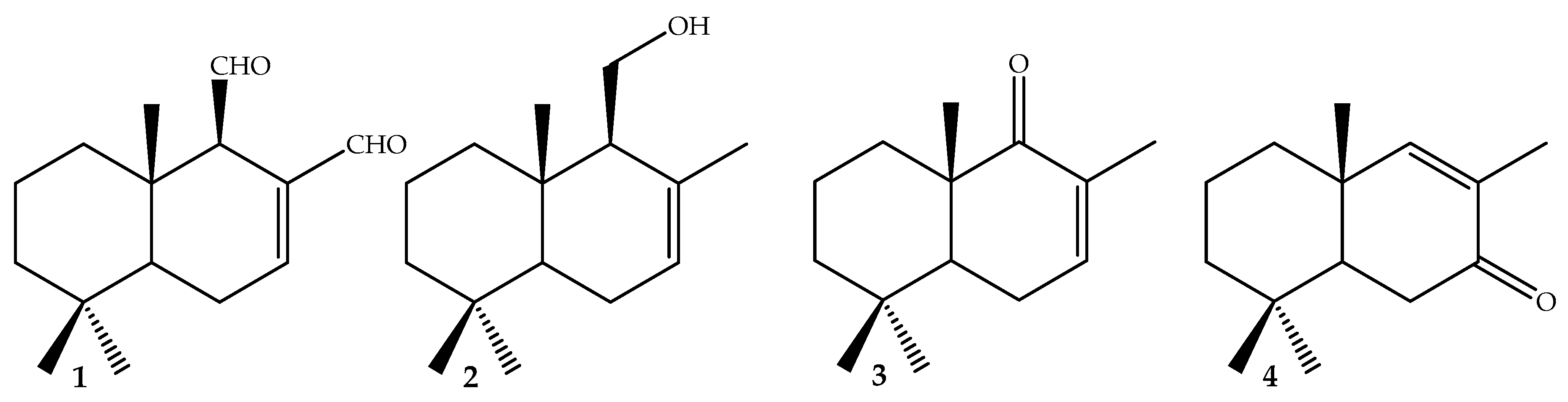
3.2. Isolation, Synthesis, and Characterization of Compounds 1–4
3.3. Microorganisms
3.4. In Vitro Antibacterial Assay
3.5. Effect of Compounds on Mycelia Growth of Phytophthora and Fusarium
3.6. Measurement of Cellular Leakage
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Albornoz, F.; Torres, A.; Tapia, M.L.; Acevedo, E. Cultivo de tomate (Lycopersicon esculentum mill.) hidropónico con agua desalinizada y desborificada en el valle de Lluta. Idesia 2007, 25, 73–80. [Google Scholar] [CrossRef]
- Ulrichs, C.; Fischer, G.; Büttner, C.; Mewis, I. Comparison of lycopene, β-carotene and phenolic contents of tomato using conventional and ecological horticultural practices, and arbuscular mycorrhizal fungi (AMF). Agron. Colomb. 2008, 26, 40–46. [Google Scholar]
- Villarreal Romero, M.; Parra Terraza, S.; Sánchez Peña, P.; Hernández Verdugo, S.; Osuna Enciso, T.; Corrales Madrid, J.L.; Armenta Bojorquez, A.D. Fertirrigación con diferentes formas de nitrógeno en el cultivo de tomate en un suelo arcilloso. Interciencia 2009, 34, 135–139. [Google Scholar]
- Available online: http://cei-rd.gov.do/ceird/transparencia/Default.aspx (accessed on 22 July 2018).
- Available online: http://faostat.fao.org/DesktopDefault (accessed on 22 July 2018).
- Available online: https://www.worldatlas.com/articles/which-are-the-world-s-leading-tomato-producing-countries.html (accessed on 3 August 2018).
- Daferera, D.; Ziogas, B.; Polissiou, M. The effectiveness of plant essential oils on the growth of Botrytis cinerea, Fusarium sp. and Clavibacter michiganensis subsp. michiganensis. Crop Prot. 2003, 22, 39–44. [Google Scholar] [CrossRef]
- Sandoval, C. Manual Técnico Manejo integrado de Enfermedades en Cultivos Hidropónicos; FAO & Universidad de Talca: Talca, Chile, 2004; pp. 31–53. [Google Scholar]
- Almeida, N.; Yan, S.; Lindeberg, M.; Studholme, D.; Schneider, D.; Condon, B.; Liu, H.; Viana, C.; Warren, A.; Evans, C.; et al. A draft genome sequence of Pseudomonas syringae pv. tomato T1 reveals a type III effector repertoire significantly divergent from that of Pseudomonas syringae pv. tomato DC3000. Mol. Plant Microbe. Interact. 2009, 22, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, P. Detection of fungal pathogens in plants. In Microbial Plant Pathogens-Detection and Disease Diagnosis; Springer: Dordrecht, The Netherlands, 2011; pp. 5–199. [Google Scholar]
- Oyarzun, P.; Gallegos, P.; Asaquibay, C.; Forbes, G.; Ochoa, J.; Paucar, B.; Prado, M.; Revelo, J.; Sherwood, S.; Fumisaca, F. El cultivo de la papa en Ecuador; Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP): Quito, Ecuador, 2002; pp. 85–169. [Google Scholar]
- Lamour, K.; Kamoun, S. Oomycete Genetics and Genomics: Diversity, Interactions, and Research Tools; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 1–24. [Google Scholar]
- Anke, H.; Sterner, O. Comparison of the antimicrobial and cytotoxic activities of twenty unsaturated sesquiterpene dialdehydes from plants and mushrooms. Planta Med. 1991, 57, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Moodie, L.W.K.; Trepos, R.; Cervin, G.; Larsen, L.; Larsen, D.S.; Pavia, H.; Hellio, C.; Cahill, P.; Svenson, J. Probing the structure—Activity relationship of the natural antifouling agent polygodial against both micro-and macrofoulers by semisynthetic modification. J. Nat. Prod. 2017, 80, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Dacunto, M. Total Synthesis of Terpenoidic Unsatured Dialdehydes and Evaluation of their Activity towards TRP Receptors. Ph.D. Thesis, Università Degli Studi di Salerno, Salerno, Italy, 2012. [Google Scholar]
- van Beek, T.A.; de Groot, A.E. Terpenoid antifeedants, part I. An overview of terpenoid antifeedants of natural origin. Rec. Trav. Chim. Pays-Bas. 1986, 105, 513–527. [Google Scholar] [CrossRef]
- Montenegro, I.J.; del Corral, S.; Diaz-Napal, G.; Carpinella, M.C.; Mellado, M.; Madrid, A.M.; Villena, J.; Palacios, S.M.; Cuellar, M.A. Antifeedant effect of polygodial and drimenol derivatives against Spodoptera frugiperda and Epilachna paenulata and quantitative structure-activity analysis. Pest Manag. Sci. 2018, 74, 1623–1629. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Fujita, K.; Lee, S.H. Antifungal mechanism of polygodial. J. Agric. Food Chem. 2001, 49, 1607–1611. [Google Scholar] [CrossRef] [PubMed]
- Derita, M.; Montenegro, I.; Garibotto, F.; Enriz, R.D.; Fritis, M.C.; Zacchino, S.A. Structural requirements for the antifungal activities of natural drimane sesquiterpenes and analogues, supported by conformational and electronic studies. Molecules 2013, 18, 2029–2051. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, I.; Pino, L.; Werner, E.; Madrid, A.; Espinoza, L.; Moreno, L.; Villena, J.; Cuellar, M. Comparative study on the larvicidal activity of drimane sesquiterpenes and nordrimane compounds against Drosophila melanogaster til-til. Molecules 2013, 18, 4192–4208. [Google Scholar] [CrossRef] [PubMed]
- Huq, A.K.M.; Jamal, J.A.; Stanslas, J. Ethnobotanical, phytochemical, pharmacological, and toxicological aspects of Persicaria hydropiper (L.) Delarbre. Evid. Based Compl. Alternat. Med. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, I. Estudio Comparativo de Actividades Biológicas Para Metabolitos Secundarios y Derivados Sintéticos Desde Drimys Winteri Forst. Ph.D. Thesis, Universidad Técnica Federico Santa María, Valparaiso, Chile, 2014. [Google Scholar]
- Montenegro, I.; Tomasoni, G.; Bosio, C.; Quiñones, N.; Madrid, A.; Carrasco, H.; Olea, A.; Martinez, R.; Cuellar, M.; Villena, J. Study on the cytotoxic activity of drimane sesquiterpenes and nordrimane compounds against cancer cell lines. Molecules 2014, 19, 18993–19006. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Fujita, K.; Lee, S.H.; Ha, T.J. Antibacterial activity of polygodial. Phytother. Res. 2005, 19, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Cimino, G.; Sodano, G.; Spinella, A. Correlation of the reactivity of 1, 4-dialdehydes with methylamine in biomimetic conditions to their hot taste: Covalent binding to primary amines as a molecular mechanism in hot taste receptors. Tetrahedron 1987, 43, 5401–5410. [Google Scholar] [CrossRef]
- Fujita, K.I.; Kubo, I. Multifunctional action of antifungal polygodial against Saccharomyces cerevisiae: involvement of pyrrole formation on cell surface in antifungal action. Phytother. Res. Med. Chem. 2005, 13, 6742–6747. [Google Scholar] [CrossRef] [PubMed]
- Montenegro, I. Estudio Preliminar del Efecto Biológico de Productos Naturales en Termitas, Bacterias y Hongos. Bachelor’s Thesis, Título de Bioquímico-Universidad Austral de Chile, Valdivia, Chile, 2006. [Google Scholar]
- Derita, M.; Leiva, M.; Zacchino, S. Influence of plant part, season of collection and content of the main active constituent, on the antifungal properties of Polygonum acuminatum Kunth. J. Ethnopharmacol. 2009, 124, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, H.; Robles-Kelly, C.; Rubio, J.; Olea, A.F.; Martínez, R.; Silva-Moreno, E. Antifungal effect of polygodial on Botrytis cinerea, a fungal pathogen affecting table grapes. Int. J. Mol. Sci. 2017, 18, 2251. [Google Scholar] [CrossRef] [PubMed]
- Castelli, M.V.; Lodeyro, A.F.; Malheiros, A.; Zacchino, S.A.; Roveri, O.A. Inhibition of the mitochondrial ATP synthesis by polygodial, a naturally occurring dialdehyde unsaturated sesquiterpene. Biochem. Pharmacol. 2005, 70, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Dasari, S.; Samy, A.L.; Narvekar, P.; Dontaraju, V.S.; Dasari, R.; Kornienko, A.; Munirathinam, G. Polygodial analog induces apoptosis in LNCaP prostate cancer cells. Eur. J. Pharmacol. 2018, 828, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Robles-Kelly, C.; Rubio, J.; Thomas, M.; Sedán, C.; Martinez, R.; Olea, A.F.; Carrasco, H.; Taborga, L.; Silva-Moreno, E. Effect of drimenol and synthetic derivatives on growth and germination of Botrytis cinerea: Evaluation of possible mechanism of action. Pestic. Biochem. Physiol. 2017, 141, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Moreno, L.; Cortés, M.; Armstrong, V.; Bailén, M.; González, A. Antifeedant Activity of some polygodial derivatives. Z. Naturforsch. C 2008, 63, 215–220. [Google Scholar] [CrossRef]
- Rodríguez, B.; Zapata, N.; Medina, P.; Viñuela, E. A complete 1H and 13C-NMR data assignment for four drimane sesquiterpenoids isolated from Drimys winteri. Magn. Reson. Chem. 2005, 43, 82–84. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, M.; Besoain, X.; Durand, K.; Cesbron, S.; Fuentes, S.; Claverías, F.; Jacques, M.A.; Seeger, S. Clavibacter michiganensis subsp. michiganensis strains from Central Chile exhibit low genetic diversity and sequence types match strains in other parts of the world. Plant Pathol. 2018. [Google Scholar] [CrossRef]
- Peterson, L.R.; Shanholtzer, C.J. Tests for bactericidal effects of antimicrobial agents: technical performance and clinical relevance. Clin. Microbiol. Rev. 1992, 5, 420–432. [Google Scholar] [CrossRef] [PubMed]
- De León, L.; Siverio, F.; López, M.M.; Rodríguez, A. Comparative efficiency of chemical compounds for in vitro and in vivo activity against Clavibacter michiganensis subsp. michiganensis, the causal agent of tomato bacterial canker. Crop Prot. 2008, 27, 1277–1283. [Google Scholar]
- Elliott, M.; Shamoun, S.F.; Sumampong, G. Effects of systemic and contact fungicides on life stages and symptom expression of Phytophthora ramorum in vitro and in planta. Crop Prot. 2015, 67, 136–144. [Google Scholar] [CrossRef]
- Lunde, C.; Kubo, I. Effect of polygodial on the mitochondrial ATPase of Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 2000, 44, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Flores, S.; Montenegro, I.; Villena, J.; Cuellar, M.; Werner, E.; Godoy, P.; Madrid, A. Synthesis and evaluation of novel oxyalkylated derivatives of 2′,4′-dihydroxychalcone as anti-oomycete agents against bronopol resistant strains of Saprolegnia sp. Int. J. Mol. Sci. 2016, 17, 1366. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the meads are available from the authors. |


{kind=link}
| Compounds | MIC (µg/mL) | MBC (µg/mL) | ||
|---|---|---|---|---|
| Cmm | Pst | Cmm | Pst | |
| 1 | 16 | 32 | 64 | 32 |
| 2 | 256 | >256 | >256 | >256 |
| 3 | 256 | >256 | >256 | >256 |
| 4 | 32 | 64 | 64 | 128 |
| CuSO4·5H2O b | 25 | 12.5 | 100 | 25 |
| Dimethyl sulfoxide (DMSO c) | i | i | i | i |
| Compounds | MIC (µg/mL) | MFC (µg/mL) | ||
|---|---|---|---|---|
| Pp | Fol | Pp | Fol | |
| 1 | 64 | 64 | 128 | 128 |
| 2 | >256 | 128 | >256 | 256 |
| 3 | >256 | 256 | >256 | >256 |
| 4 | 128 | 128 | 128 | 128 |
| Difenoconazole b | 100 | 100 | 150 | 125 |
| Metalaxyl b | 10 | 125 | 20 | 150 |
| DMSO c | i | i | i | i |
| Compounds | %Damage a | |
|---|---|---|
| (100 µg/mL) | Pp | Fol |
| 1 | 100 | 100 |
| 2 | 0 | 30 |
| 3 | 0 | 0 |
| 4 | 70 | 80 |
| Difenoconazole b | 100 | 100 |
| SDS c | 100 | 100 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montenegro, I.; Madrid, A.; Cuellar, M.; Seeger, M.; Alfaro, J.F.; Besoain, X.; Martínez, J.P.; Ramirez, I.; Olguín, Y.; Valenzuela, M. Biopesticide Activity from Drimanic Compounds to Control Tomato Pathogens. Molecules 2018, 23, 2053. https://doi.org/10.3390/molecules23082053
Montenegro I, Madrid A, Cuellar M, Seeger M, Alfaro JF, Besoain X, Martínez JP, Ramirez I, Olguín Y, Valenzuela M. Biopesticide Activity from Drimanic Compounds to Control Tomato Pathogens. Molecules. 2018; 23(8):2053. https://doi.org/10.3390/molecules23082053
Chicago/Turabian StyleMontenegro, Iván, Alejandro Madrid, Mauricio Cuellar, Michael Seeger, Juan Felipe Alfaro, Ximena Besoain, Juan Pablo Martínez, Ingrid Ramirez, Yusser Olguín, and Miryam Valenzuela. 2018. "Biopesticide Activity from Drimanic Compounds to Control Tomato Pathogens" Molecules 23, no. 8: 2053. https://doi.org/10.3390/molecules23082053
APA StyleMontenegro, I., Madrid, A., Cuellar, M., Seeger, M., Alfaro, J. F., Besoain, X., Martínez, J. P., Ramirez, I., Olguín, Y., & Valenzuela, M. (2018). Biopesticide Activity from Drimanic Compounds to Control Tomato Pathogens. Molecules, 23(8), 2053. https://doi.org/10.3390/molecules23082053