
A Review of Lignan Metabolism, Milk Enterolactone Concentration, and Antioxidant Status of Dairy Cows Fed Flaxseed

Abstract
1. Introduction
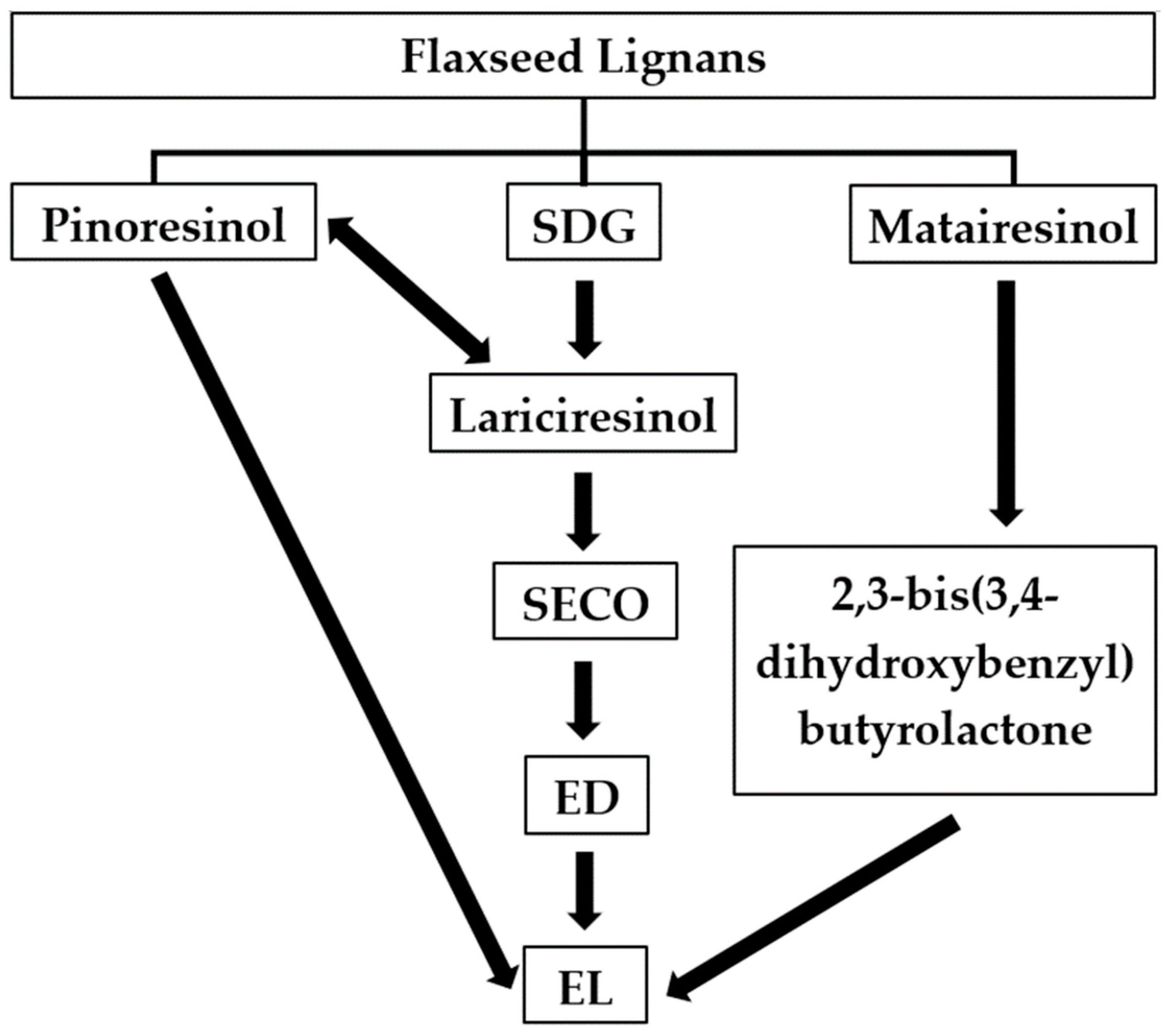
2. Metabolism of Lignans in the Gastrointestinal Tract
3. Effects of Flaxseed Products on Milk EL Concentration
3.1. Dose-Response Studies and Milk EL Concentration
3.2. Comparison of Flaxseed Products and Animal Variation in Milk EL Concentration
3.3. Impact of NSC Sources and FM on Milk EL Concentration
3.4. Dairy Breed and Milk EL Concentration
4. Pharmacokinetics of Milk EL and Potential Implications on Animal and Human Health
5. Antioxidant Activity of Flaxseed Products and Dairy Cow Health
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCG2 | ATP-binding cassette subfamily G2 |
| CAT | catalase |
| DM | dry matter |
| ED | enterodiol |
| EL | enterolactone |
| FH | flaxseed hulls |
| FM | flaxseed meal |
| FO | flaxseed oil |
| GPx | glutathione peroxidase |
| NFE2L2 | nuclear factor (erythroid-derived 2)-like 2 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NF-κB1 | nuclear factor kappa-light-chain-enhancer of activated B cells subunit 1 |
| NFR2 | nuclear factor erythroid 2–related factor 2 |
| NSC | nonstructural carbohydrate |
| PUFA | polyunsaturated fatty acids |
| ROS | reactive oxygen substances |
| SECO | secoisolariciresinol |
| SDG | secoisolariciresinol diglucoside |
| SOD | superoxide dismutase |
| TBARS | thiobarbituric acid-reactive substances |
| WF | whole flaxseed |
References
- Adolphe, J.L.; Whiting, S.J.; Juurlink, B.H.J.; Thorpe, L.U.; Alcorn, J. Health effects with consumption of the flax lignan secoisolariciresinol diglucoside. Br. J. Nutr. 2010, 103, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Högger, P. Nutrition-derived bioactive metabolites produced by gut microbiota and their potential impact on human health. Nutr. Med. 2013, 1, 1–32. [Google Scholar]
- Imran, M.; Ahmad, N.; Anjum, F.M.; Khan, M.K.; Mushtaq, Z.; Nadeem, M.; Hussain, S. Potential protective properties of flax lignan secoisolariciresinol diglucoside. Nutr. J. 2015, 14, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Carreau, C.; Flouriot, G.; Bennetau-Pelissero, C.; Potier, M. Enterodiol and enterolactone, two major diet-derived polyphenol metabolites have different impact on ERα transcriptional activation in human breast cancer cells. J. Steroid Biochem. Mol. Biol. 2008, 110, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, H.; Mazur, W. Phyto-oestrogens and western diseases. Ann. Med. 1997, 29, 95–120. [Google Scholar] [CrossRef]
- Thompson, L.U.; Robb, P.; Serraino, M.; Cheung, F. Mammalian lignan production from various foods. Nutr. Cancer 1991, 16, 43–52. [Google Scholar] [CrossRef]
- Côrtes, C.; Gagnon, N.; Benchaar, C.; Da Silva, D.; Santos, G.T.D.; Petit, H.V. In vitro metabolism of flax lignans by ruminal and faecal microbiota of dairy cows. J. Appl. Microbiol. 2008, 105, 1585–1594. [Google Scholar] [CrossRef]
- Gagnon, N.; Côrtes, C.; da Silva, D.; Kazama, R.; Benchaar, C.; dos Santos, G.; Zeoula, L.; Petit, H.V. Ruminal metabolism of flaxseed (Linum usitatissimum) lignans to the mammalian lignan enterolactone and its concentration in ruminal fluid, plasma, urine and milk of dairy cows. Br. J. Nutr. 2009, 102, 1015–1023. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, G.; Han, Z.; Yao, W.; Zhu, W. Metabolism of flaxseed lignans in the rumen and its impact on ruminal metabolism and flora. Anim. Feed Sci. Technol. 2009, 150, 18–26. [Google Scholar] [CrossRef]
- Schogor, A.L.B.; Palin, M.F.; Santos, G.T.; Benchaar, C.; Petit, H.V. β-glucuronidase activity and enterolactone concentration in ruminal fluid, plasma, urine, and milk of Holstein cows fed increased levels of flax (Linum usitatissimum) meal. Anim. Feed Sci. Technol. 2017, 223, 23–29. [Google Scholar] [CrossRef]
- Petit, H.V.; Gagnon, N. Milk concentrations of the mammalian lignans enterolactone and enterodiol, milk production, and whole tract digestibility of dairy cows fed diets containing different concentrations of flaxseed meal. Anim. Feed Sci. Technol. 2009, 152, 103–111. [Google Scholar] [CrossRef]
- Jin, J.S.; Kakiuchi, N.; Hattori, M. Enantioselective oxidation of enterodiol to enterolactone by human intestinal bacteria. Biol. Pharm. Bull. 2007, 30, 2204–2206. [Google Scholar] [CrossRef] [PubMed]
- Muehlhoff, E.; Bennett, A.; McMahon, D. Milk and Dairy Products in Human Nutrition; Food and Agriculture Organization of the United Nations (FAO): Rome, Itay, 2013; pp. 11–30. [Google Scholar]
- Alexandratos, N.; Bruinsma, J. World Agriculture Towards 2030/2050: The 2012 Revision; Food and Agriculture Organization of the United Nations (FAO): Rome, Itay, 2012; pp. 4–5. [Google Scholar]
- De Kleijn, M.J.J.; van der Schouw, Y.T.; Wilson, P.W.F.; Adlercreutz, H.; Mazur, W.; Grobbee, D.E.; Jacques, P.F. Intake of dietary phytoestrogens is low in postmenopausal women in the United States: The framingham study. J. Nutr. 2001, 131, 1826–1832. [Google Scholar] [CrossRef] [PubMed]
- Petit, H.V.; Gagnon, N.; Mir, P.S.; Cao, R.; Cui, S. Milk concentration of the mammalian lignan enterolactone, milk production, milk fatty acid profile, and digestibility in dairy cows fed diets containing whole flaxseed or flaxseed meal. J. Dairy Res. 2009, 76, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Petit, H.V.; Gagnon, N. Production performance and milk composition of dairy cows fed different concentrations of flax hulls. Anim. Feed Sci. Technol. 2011, 169, 46–52. [Google Scholar] [CrossRef]
- Brito, A.F.; Petit, H.V.; Pereira, A.B.D.; Soder, K.J.; Ross, S. Interactions of corn meal or molasses with a soybean-sunflower meal mix or flaxseed meal on production, milk fatty acid composition, and nutrient utilization in dairy cows fed grass hay-based diets. J. Dairy Sci. 2015, 98, 443–457. [Google Scholar] [CrossRef]
- Ghedini, C.P.; Moura, D.C.; Santana, R.A.V.; Oliveira, A.S.; Brito, A.F. Replacing ground corn with incremental amounts of liquid molasses does not change milk enterolactone but decreases production in dairy cows fed flaxseed meal. J. Dairy Sci. 2018, 101, 2096–2109. [Google Scholar] [CrossRef]
- Gaál, T.; Ribiczeyné-Szabó, P.; Stadler, K.; Jakus, J.; Reiczigel, J.; Kövér, P.; Mezes, M.; Sümeghy, L. Free radicals, lipid peroxidation and the antioxidant system in the blood of cows and newborn calves around calving. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 391–396. [Google Scholar] [CrossRef]
- Abuelo, A.; Hernandez, J.; Benedito, J.L.; Castillo, C. A pilot study to compare oxidative status between organically and conventionally managed dairy cattle during the transition period. Reprod. Domest. Anim. 2015, 50, 538–544. [Google Scholar] [CrossRef]
- Prasad, K. Antioxidant activity of secoisolariciresinol diglucoside-derived metabolites, secoisolariciresinol, enterodiol, and enterolactone. Int. J. Angiol. 2000, 9, 220–225. [Google Scholar] [CrossRef]
- Rajesha, J.; Murthy, K.N.C.; Kumar, M.K.; Madhusudhan, B.; Ravishankar, G.A. Antioxidant potentials of flaxseed by in vivo model. J. Agric. Food Chem. 2006, 54, 3794–3799. [Google Scholar] [CrossRef]
- Gaya, P.; Medina, M.; Sánchez-Jiménez, A.; Landete, J.M. Phytoestrogen metabolism by adult human gut microbiota. Molecules 2016, 21, 1034. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U. Flaxseed, lignans, and cancer. In Flaxseed in Human Nutrition; Thompson, L.U., Cunnane, S.C., Eds.; AOCS Press: Champaign, IL, USA, 2003; pp. 199–227. [Google Scholar]
- Morris, D.H. Flax: A Health and Nutrition Primer; Flax Council of Canada: Winnipeg, MB, Canada, 2007. [Google Scholar]
- Xie, L.; Akao, T.; Hamasaki, K.; Deyama, T.; Hattori, M. Biotransformation of pinoresinol diglucoside to mammalian lignans by human intestinal microflora, and isolation of Enterococcus faecalis strain PDG-1 responsible for the transformation of (+)-pinoresinol to (+)-lariciresinol. Chem. Pharm. Bull. 2003, 51, 508–515. [Google Scholar] [CrossRef]
- Clavel, T.; Borrmann, D.; Braune, A.; Doré, J.; Blaut, M. Occurrence and activity of human intestinal bacteria involved in the conversion of dietary lignans. Anaerobe 2006, 12, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Clavel, T.; Doré, J.; Blaut, M. Bioavailability of lignans in human subjects. Nutr. Res. Rev. 2006, 19, 187–196. [Google Scholar] [CrossRef]
- Clavel, T.; Lippman, R.; Gavini, F.; Doré, J.; Blaut, M. Clostridium saccharogumia sp. nov. and Lactonifactor longoviformis gen. nov., sp. nov., two novel human faecal bacteria involved in the conversion of the dietary phytoestrogen secoisolariciresinol diglucoside. Syst. Appl. Microbiol. 2007, 30, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhao, Y.; Nakamura, N.; Akao, T.; Kakiuchi, N.; Min, B.; Hattori, M. Enantioselective dehydroxylation of enterodiol and enterolactone precursors by human intestinal bacteria. Biol. Pharm. Bull. 2007, 30, 2113–2119. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Hattori, M. Human intestinal bacterium, strain END-2 is responsible for demethylation as well as lactonization during plant lignan metabolism. Biol. Pharm. Bull. 2010, 33, 1443–1447. [Google Scholar] [CrossRef]
- Roncaglia, L.; Amaretti, A.; Raimondi, S.; Leonardi, A.; Rossi, M. Role of bifidobacteria in the activation of the lignan secoisolariciresinol diglucoside. Appl. Microbiol. Biotechnol. 2011, 92, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Jansen, G.H.E.; Arts, I.C.W.; Nielen, M.W.F.; Müller, M.; Hollman, P.C.H.; Keijer, J. Uptake and metabolism of enterolactone and enterodiol by human colon epithelial cells. Arch. Biochem. Biophys. 2005, 435, 74–82. [Google Scholar] [CrossRef]
- Lin, C.; S Krol, E.; Alcorn, J. The comparison of rat and human intestinal and hepatic glucuronidation of enterolactone derived from flaxseed lignans. J. Nat. Prod. 2013, 3, 159–171. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Adlercreutz, H. Mammalian lignans and phytooestrogens recent studies on their formation, metabolism and biological role in health and disease. In Role of the Gut Flora in Toxicity and Cancer; Rowland, I.R., Ed.; Elsevier Inc.: Carshalton, UK, 1988. [Google Scholar]
- Hullar, M.A.J.; Lancaster, S.M.; Li, F.; Tseng, E.; Beer, K.; Atkinson, C.; Wähälä, K.; Copeland, W.K.; Randolph, T.W.; Newton, K.M. Enterolignan-producing phenotypes are associated with increased gut microbial diversity and altered composition in premenopausal women in the United States. Cancer Epidemiol. Biomarkers Prev. 2015, 24, 546–554. [Google Scholar] [CrossRef] [PubMed]
- Bess, E.N.; Bisanz, J.E.; Spanogiannopoulos, P.; Ang, Q.Y.; Bustion, A.; Kitamura, S.; Alba, D.L.; Wolan, D.W.; Koliwad, S.K.; Turnbaugh, P.J. The genetic basis for the cooperative bioactivation of plant lignans by a human gut bacterial consortium. bioRxiv 2018, 357640. [Google Scholar] [CrossRef]
- Brossillon, V.; Reis, S.F.; Moura, D.C.; Galvão, J.G.B., Jr.; Oliveira, A.S.; Côrtes, C.; Brito, A.F. Production, milk and plasma fatty acid profile, and nutrient utilization in Jersey cows fed flaxseed oil and corn grain with different particle size. J. Dairy Sci. 2018, 101, 2127–2143. [Google Scholar] [CrossRef] [PubMed]
- Henderson, C. The effects of fatty acids on pure cultures of rumen bacteria. J. Agric. Sci. 1973, 81, 107–112. [Google Scholar] [CrossRef]
- Maia, F.J.; Branco, A.F.; Mouro, G.F.; Coneglian, S.M.; Santos, G.T.d.; Minella, T.F.; Guimarães, K.C. Feeding vegetable oil to lactating goats: Milk production and composition and milk fatty acids profile. R. Bras. Zootec. 2006, 35, 1504–1513. [Google Scholar] [CrossRef]
- Schogor, A.L.B.; Huws, S.A.; Santos, G.T.D.; Scollan, N.D.; Hauck, B.D.; Winters, A.L.; Kim, E.J.; Petit, H.V. Ruminal Prevotella spp. may play an important role in the conversion of plant lignans into human health beneficial antioxidants. PLoS ONE 2014, 9, e87949. [Google Scholar] [CrossRef]
- Lundh, T.J.O. Conjugation of the plant estrogens formononetin and daidzein and their metabolite equol by gastrointestinal epithelium from cattle and sheep. J. Agric. Food Chem. 1990, 38, 1012–1016. [Google Scholar] [CrossRef]
- Njåstad, K.M.; Adler, S.A.; Hansen-Møller, J.; Thuen, E.; Gustavsson, A.M.; Steinshamn, H. Gastrointestinal metabolism of phytoestrogens in lactating dairy cows fed silages with different botanical composition. J. Dairy Sci. 2014, 97, 7735–7750. [Google Scholar] [CrossRef]
- Jenab, M.; Thompson, L.U. The influence of flaxseed and lignans on colon carcinogenesis and β-glucuronidase activity. Carcinogenesis 1996, 17, 1343–1348. [Google Scholar] [CrossRef]
- Raffaelli, B.; Hoikkala, A.; Leppälä, E.; Wähälä, K. Enterolignans. J. Chromatogr. B 2002, 777, 29–43. [Google Scholar] [CrossRef]
- De Marchi, F.E.; Palin, M.F.; Santos, G.T.; Benchaar, C.; Petit, H.V. Effects of duodenal infusion of sunflower oil on β-glucuronidase activity and enterolactone concentration in dairy cows fed flax meal. Anim. Feed Sci. Technol. 2016, 220, 143–150. [Google Scholar] [CrossRef]
- Côrtes, C.; da Silva-Kazama, D.; Kazama, R.; Benchaar, C.; dos Santos, G.; Zeoula, L.M.; Gagnon, N.; Petit, H.V. Effects of abomasal infusion of flaxseed (Linum usitatissimum) oil on microbial β-glucuronidase activity and concentration of the mammalian lignan enterolactone in ruminal fluid, plasma, urine and milk of dairy cows. Br. J. Nutr. 2013, 109, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Beaud, D.; Tailliez, P.; Anba-Mondoloni, J. Genetic characterization of the β-glucuronidase enzyme from a human intestinal bacterium, Ruminococcus gnavus. Microbiology 2005, 151, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, R.R. Anatomy of the gastro-intestinal tract. In The Ruminant Animal: Digestive Phisiology and Nutrition; Church, D.C., Ed.; Waveland Press: Long Grove, IL, USA, 1993; pp. 14–43. [Google Scholar]
- Dehority, B.A. Rumen Microbiology; Nottingham University Press: Nottingham, UK, 2003. [Google Scholar]
- Vargas, J.E.; Andrés, S.; Snelling, T.J.; López-Ferreras, L.; Yáñez-Ruíz, D.R.; García-Estrada, C.; López, S. Effect of sunflower and marine oils on ruminal microbiota, in vitro fermentation and digesta fatty acid profile. Front. Microbiol. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed]
- Petit, H.V.; Gagnon, N. Concentration of the mammalian lignans enterolactone and enterodiol in milk of cows fed diets containing different concentrations of whole flaxseed. Animal 2009, 3, 1428–1435. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, N.; Côrtes, C.; Petit, H.V. Weekly excretion of the mammalian lignan enterolactone in milk of dairy cows fed flaxseed meal. J. Dairy Res. 2009, 76, 455–458. [Google Scholar] [CrossRef]
- Petit, H.V.; Côrtes, C.; da Silva, D.; Kazama, R.; Gagnon, N.; Benchaar, C.; dos Santos, G.T.; Zeoula, L.M. The interaction of monensin and flaxseed hulls on ruminal and milk concentration of the mammalian lignan enterolactone in late-lactating dairy cows. J. Dairy Res. 2009, 76, 475–482. [Google Scholar] [CrossRef]
- Côrtes, C.; Palin, M.F.; Gagnon, N.; Benchaar, C.; Lacasse, P.; Petit, H.V. Mammary gene expression and activity of antioxidant enzymes and concentration of the mammalian lignan enterolactone in milk and plasma of dairy cows fed flax lignans and infused with flax oil in the abomasum. Br. J. Nutr. 2012, 108, 1390–1398. [Google Scholar] [CrossRef]
- Lima, L.S.; Palin, M.F.; Santos, G.T.; Benchaar, C.; Petit, H.V. Dietary flax meal and abomasal infusion of flax oil on microbial β-glucuronidase activity and concentration of enterolactone in ruminal fluid, plasma, urine and milk of dairy cows. Anim. Feed Sci. Technol. 2016, 215, 85–91. [Google Scholar] [CrossRef]
- Ghedini, C.P. Improving the Understanding of Different Diets on the Concentration and Metabolism of the Mammalian Lignan Enterolactone in Dairy Cattle. Ph.D. Thesis, University of New Hampshire, Durham, NH, USA, 2017. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle, 7th ed.; NRC: Washington, DC, USA, 2001. [Google Scholar]
- Olmos Colmenero, J.O.; Broderick, G. Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. J. Dairy Sci. 2006, 89, 1704–1712. [Google Scholar] [CrossRef]
- Höjer, A.; Adler, S.; Purup, S.; Hansen-Møller, J.; Martinsson, K.; Steinshamn, H.; Gustavsson, A.M. Effects of feeding dairy cows different legume-grass silages on milk phytoestrogen concentration. J. Dairy Sci. 2012, 95, 4526–4540. [Google Scholar] [CrossRef] [PubMed]
- Miguel, V.; Otero, J.A.; Garcia-Villalba, R.; Tomás-Barberán, F.; Espin, J.C.; Merino, G.; Álvarez, A.I. Role of ABCG2 in transport of the mammalian lignan enterolactone and its secretion into milk in Abcg2 knockout mice. Drug Metab. Dispos. 2014, 42, 943–946. [Google Scholar] [CrossRef]
- Zhu, W.; Xu, H.; Wang, S.W.; Hu, M. Breast cancer resistance protein (BCRP) and sulfotransferases contribute significantly to the disposition of genistein in mouse intestine. AAPS J. 2010, 12, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, A.I.; Vallejo, F.; Barrera, B.; Merino, G.; Prieto, J.G.; Tomás-Barberán, F.; Espín, J.C. Bioavailability of the glucuronide and sulfate conjugates of genistein and daidzein in Bcrp1 knockout mice. Drug Metab. Dispos. 2011, 39, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.W.; Li, Y.; Paxton, J.W.; Birch, N.P.; Scheepens, A. Identification of novel dietary phytochemicals inhibiting the efflux transporter breast cancer resistance protein (BCRP/ABCG2). Food Chem. 2013, 138, 2267–2274. [Google Scholar] [CrossRef] [PubMed]
- Otero, J.A.; Miguel, V.; González-Lobato, L.; García-Villalba, R.; Espín, J.C.; Prieto, J.G.; Merino, G.; Álvarez, A.I. Effect of bovine ABCG2 polymorphism Y581S SNP on secretion into milk of enterolactone, riboflavin and uric acid. Animal 2016, 10, 238–247. [Google Scholar] [CrossRef]
- Real, R.; González-Lobato, L.; Baro, M.F.; Valbuena, S.; de la Fuente, A.; Prieto, J.G.; Alvarez, A.I.; Marques, M.M.; Merino, G. Analysis of the effect of the bovine adenosine triphosphate-binding cassette transporter G2 single nucleotide polymorphism Y581S on transcellular transport of veterinary drugs using new cell culture models. J. Anim. Sci. 2011, 89, 4325–4338. [Google Scholar] [CrossRef]
- Otero, J.A.; Real, R.; de la Fuente, Á.; Prieto, J.G.; Marqués, M.; Álvarez, A.I.; Merino, G. The bovine ATP-binding cassette transporter ABCG2 Tyr581Ser single-nucleotide polymorphism increases milk secretion of the fluoroquinolone danofloxacin. Drug Metab. Dispos. 2013, 41, 546–549. [Google Scholar] [CrossRef]
- Otero, J.A.; Barrera, B.; de la Fuente, A.; Prieto, J.G.; Marqués, M.; Alvarez, A.I.; Merino, G. The gain-of-function Y581S polymorphism of the ABCG2 transporter increases secretion into milk of danofloxacin at the therapeutic dose for mastitis treatment. J. Dairy Sci. 2015, 98, 312–317. [Google Scholar] [CrossRef]
- Mendez, M.A.; Anthony, M.S.; Arab, L. Soy-based formulae and infant growth and development: A review. J. Nutr. 2002, 132, 2127–2130. [Google Scholar] [CrossRef] [PubMed]
- Tuohy, P. Soy infant formula and phytoestrogens. J. Paediatr. Child Health 2003, 39, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Troina, A.A.; Figueiredo, M.S.; Passos, M.C.F.; Reis, A.M.; Oliveira, E.; Lisboa, P.C.; Moura, E.G. Flaxseed bioactive compounds change milk, hormonal and biochemical parameters of dams and offspring during lactation. Food Chem. Toxicol. 2012, 50, 2388–2396. [Google Scholar] [CrossRef] [PubMed]
- Steinshamn, H.; Purup, S.; Thuen, E.; Hansen-Møller, J. Effects of clover-grass silages and concentrate supplementation on the content of phytoestrogens in dairy cow milk. J. Dairy Sci. 2008, 91, 2715–2725. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, I.M.C.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, D.G.; Robertson, S.; Choung, J.J. Sugars versus starch as supplements to grass silage: Effects on ruminal fermentation and the supply of microbial protein to the small intestine, estimated from the urinary excretion of purine derivatives, in sheep. J. Sci. Food Agric. 1993, 63, 189–194. [Google Scholar] [CrossRef]
- Broderick, G.A.; Radloff, W.J. Effect of molasses supplementation on the production of lactating dairy cows fed diets based on alfalfa and corn silage. J. Dairy Sci. 2004, 87, 2997–3009. [Google Scholar] [CrossRef]
- Purushe, J.; Fouts, D.E.; Morrison, M.; White, B.A.; Mackie, R.I.; Coutinho, P.M.; Henrissat, B.; Nelson, K.E.; Bacteria, N.A.C.f.R. Comparative genome analysis of Prevotella ruminicola and Prevotella bryantii: Insights into their environmental niche. Microb. Ecol. 2010, 60, 721–729. [Google Scholar] [CrossRef]
- Li, X.; Park, B.K.; Shin, J.S.; Choi, S.H.; Smith, S.B.; Yan, C.G. Effects of dietary linseed oil and propionate precursors on ruminal microbial community, composition, and diversity in Yanbian yellow cattle. PLoS ONE 2015, 10, e0126473. [Google Scholar] [CrossRef]
- Aikman, P.C.; Reynolds, C.K.; Beever, D.E. Diet digestibility, rate of passage, and eating and rumination behavior of Jersey and Holstein cows. J. Dairy Sci. 2008, 91, 1103–1114. [Google Scholar] [CrossRef]
- Heinonen, S.; Nurmi, T.; Liukkonen, K.; Poutanen, K.; Wähälä, K.; Deyama, T.; Nishibe, S.; Adlercreutz, H. In vitro metabolism of plant lignans: New precursors of mammalian lignans enterolactone and enterodiol. J. Agric. Food Chem. 2001, 49, 3178–3186. [Google Scholar] [CrossRef] [PubMed]
- Milder, I.E.J.; Feskens, E.J.M.; Arts, I.C.W.; de Mesquita, H.B.B.; Hollman, P.C.H.; Kromhout, D. Intake of the plant lignans secoisolariciresinol, matairesinol, lariciresinol, and pinoresinol in Dutch men and women. J. Nutr. 2005, 135, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Kuijsten, A.; Arts, I.C.W.; Hollman, P.C.H.; van’t Veer, P.; Kampman, E. Plasma enterolignans are associated with lower colorectal adenoma risk. Cancer Epidemiol. Biomarkers Prev. 2006, 15, 1132–1136. [Google Scholar] [CrossRef] [PubMed]
- Vanharanta, M.; Voutilainen, S.; Lakka, T.A.; van der Lee, M.; Adlercreutz, H.; Salonen, J.T. Risk of acute coronary events according to serum concentrations of enterolactone: A prospective population-based case-control study. Lancet 1999, 354, 2112–2115. [Google Scholar] [CrossRef]
- Vanharanta, M.; Voutilainen, S.; Rissanen, T.H.; Adlercreutz, H.; Salonen, J.T. Risk of cardiovascular disease–related and all-cause death according to serum concentrations of enterolactone: Kuopio ischaemic heat disease risk factor study. Arch. Intern. Med. 2003, 163, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, P.D.; Lam, Y.; Thompson, L.U. Human metabolism of mammalian lignan precursors in raw and processed flaxseed. Am. J. Clin. Nutr. 1999, 69, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, N.M.; Smeds, A.; Mäkelä, S.I.; Ämmälä, J.; Hakala, K.; Pihlava, J.M.; Ryhänen, E.L.; Sjöholm, R.; Santti, R. Structural determinants of plant lignans for the formation of enterolactone in vivo. J. Chromatogr. B 2002, 777, 311–319. [Google Scholar] [CrossRef]
- Kuijsten, A.; Arts, I.C.W.; Vree, T.B.; Hollman, P.C.H. Pharmacokinetics of enterolignans in healthy men and women consuming a single dose of secoisolariciresinol diglucoside. J. Nutr. 2005, 135, 795–801. [Google Scholar] [CrossRef]
- Guglielmini, P.; Rubagotti, A.; Boccardo, F. Serum enterolactone levels and mortality outcome in women with early breast cancer: A retrospective cohort study. Breast Cancer Res. Treat. 2012, 132, 661–668. [Google Scholar] [CrossRef]
- Grummer, R.R. Impact of changes in organic nutrient metabolism on feeding the transition dairy cow. J. Anim. Sci. 1995, 73, 2820–2833. [Google Scholar] [CrossRef]
- Miller, J.K.; Brzezinska-Slebodzinska, E.; Madsen, F.C. Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 1993, 76, 2812–2823. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Aitken, S.L. Impact of oxidative stress on the health and immune function of dairy cattle. Vet. Immunol. Immunopathol. 2009, 128, 104–109. [Google Scholar] [CrossRef]
- Brenneisen, P.; Steinbrenner, H.; Sies, H. Selenium, oxidative stress, and health aspects. Mol. Aspects Med. 2005, 26, 256–267. [Google Scholar] [CrossRef]
- Finch, J.M.; Turner, R.J. Effects of selenium and vitamin E on the immune responses of domestic animals. Res. Vet. Sci. 1996, 60, 97–106. [Google Scholar] [CrossRef]
- Matés, J.M.; Sánchez-Jiménez, F. Antioxidant enzymes and their implications in pathophysiologic processes. Front. Biosci. 1999, 4, 0339–0345. [Google Scholar] [CrossRef]
- Lledías, F.; Rangel, P.; Hansberg, W. Oxidation of catalase by singlet oxygen. J. Biol. Chem. 1998, 273, 10630–10637. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Harding, J.J. Glycation-induced inactivation and loss of antigenicity of catalase and superoxide dismutase. Biochem. J. 1997, 328, 599–605. [Google Scholar] [CrossRef]
- Dribnenki, J.C.P.; McEachern, S.F.; Green, A.G.; Kenaschuk, E.O.; Rashid, K.Y. Linola™’1084’low-linolenic acid flax. Can. J. Plant Sci. 1999, 79, 607–609. [Google Scholar] [CrossRef]
- Lima, L.S.; Palin, M.F.; Santos, G.T.; Benchaar, C.; Petit, H.V. Effects of supplementation of flax meal and flax oil on mammary gene expression and activity of antioxidant enzymes in mammary tissue, plasma and erythrocytes of dairy cows. Livest. Sci. 2015, 176, 196–204. [Google Scholar] [CrossRef]
- Schogor, A.L.B.; Palin, M.-F.; dos Santos, G.T.; Benchaar, C.; Lacasse, P.; Petit, H.V. Mammary gene expression and activity of antioxidant enzymes and oxidative indicators in the blood, milk, mammary tissue and ruminal fluid of dairy cows fed flax meal. Br. J. Nutr. 2013, 110, 1743–1750. [Google Scholar] [CrossRef]
- De Marchi, F.E.; Palin, M.F.; Dos Santos, G.T.; Lima, L.S.; Benchaar, C.; Petit, H.V. Flax meal supplementation on the activity of antioxidant enzymes and the expression of oxidative stress- and lipogenic-related genes in dairy cows infused with sunflower oil in the abomasum. Anim. Feed Sci. Technol. 2015, 199, 41–50. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Clair, D.K.S. Regulation of superoxide dismutase genes: Implications in disease. Free Radic. Biol. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef]
- Kang, K.W.; Lee, S.J.; Kim, S.G. Molecular mechanism of nrf2 activation by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef]
- Zhu, H.; Itoh, K.; Yamamoto, M.; Zweier, J.L.; Li, Y. Role of Nrf2 signaling in regulation of antioxidants and phase 2 enzymes in cardiac fibroblasts: Protection against reactive oxygen and nitrogen species-induced cell injury. FEBS Lett. 2005, 579, 3029–3036. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. Cold Spring Harbor Perspectives in Biology. Available online: https://cshperspectives.cshlp.org/content/early/2009/10/04/cshperspect.a001651.abstract (accessed on 24 November 2018).
- Abarikwu, S.O.; Pant, A.B.; Farombi, E.O. Quercetin decreases steroidogenic enzyme activity, NF-κB expression, and oxidative stress in cultured Leydig cells exposed to atrazine. Mol. Cell. Biochem. 2013, 373, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Do Prado, R.M.; Palin, M.F.; Do Prado, I.N.; Dos Santos, G.T.; Benchaar, C.; Petit, H.V. Milk yield, milk composition, and hepatic lipid metabolism in transition dairy cows fed flaxseed or linola. J. Dairy Sci. 2016, 99, 8831–8846. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H. Estimation of peroxidative damage: A critical review. Pathol. Biol. 1996, 44, 25–28. [Google Scholar]
- Chang, Y.; Chang, W.; Tsai, N.; Huang, C.; Kung, C.; Su, Y.; Lin, W.; Cheng, B.; Su, C.; Chiang, Y. The roles of biomarkers of oxidative stress and antioxidant in Alzheimer’s disease: A systematic review. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.S.; Palin, M.F.; Santos, G.T.; Benchaar, C.; Lima, L.C.R.; Chouinard, P.Y.; Petit, H.V. Effect of flax meal on the production performance and oxidative status of dairy cows infused with flax oil in the abomasum. Livest. Sci. 2014, 170, 53–62. [Google Scholar] [CrossRef]
- De Marchi, F.E.; Santos, G.T.; Petit, H.V.; Benchaar, C. Oxidative status of dairy cows fed flax meal and infused with sunflower oil in the abomasum. Anim. Feed Sci. Technol. 2017, 228, 115–122. [Google Scholar] [CrossRef]
- Scislowski, V.; Bauchart, D.; Gruffat, D.; Laplaud, P.M.; Durand, D. Effects of dietary n-6 or n-3 polyunsaturated fatty acids protected or not against ruminal hydrogenation on plasma lipids and their susceptibility to peroxidation in fattening steers. J. Anim. Sci. 2005, 83, 2162–2174. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| References | No. of Cows | DIM 1 | Experimental Design 2 | Treatments 3 | Milk Enterolactone Concentration 4 |
|---|---|---|---|---|---|
| [16] | 24 | 119 | RCB | CON, 10% FM, 10% WF | 10% FM = 10% WF > CON |
| [8] | 4 | 92 | 4 × 4 Latin square | FO & FH at ABO/ABO, RUM/ABO, RUM/RUM, ABO/RUM | ABO/RUM = RUM/RUM > RUM/ABO = ABO/ABO |
| [11] | 32 | 231 | RCB | 0%, 5%, 10%, 15% FM | Linear increase |
| [54] | 12 | 61 | RCB | CON, 20% FM | 20% FM > CON |
| [53] | 32 | 175 | RCB | 0%, 5%, 10%, 15% WF | Tendency for linear increase |
| [55] | 4 | 190 | 4 × 4 Latin square | CON, 20% FH, MON, 20% FH + MON | 20% FH = 20% FH + MON > CON = MON |
| [17] | 45 | 140 | RCB | 0%, 5%, 10%, 15%, 20% FH | Linear increase |
| [56] | 8 | 163 | 4 × 4 Latin square | CON, 9.88% FH, 500 g/d FO at ABO, 9.88% FH + 500 g/d FO at ABO | 9.88% FH = 9.88% FH + 500 g/d > CON > 500 g/d FO |
| [48] | 6 | 95 | 6 × 6 Latin square | 2 × 3 factorial: FH (0%, 15.9%) × FO (0, 250, 500 g/d) | 15.9% FH diets > 0% FH diets |
| [18] | 16 | 135 | 4 × 4 Latin square | 2 × 2 factorial: GRC + 16% SBM-SFM mix, GRC + 16% FM, LM + 16% SBM-SFM mix, LM + 16% FM | 16% FM diets > 16% SBM-SFM mix diets & LM diets > GRC diets |
| [47] | 8 | 56 | 4 × 4 Latin square | 2 × 2 factorial: CON, 13.7% FM, 250 g/d SO at ABO, 13.7% FM + 250 g/d SO at ABO | No treatment differences |
| [57] | 8 | 108 | 4 × 4 Latin square | 2 × 2 factorial: CON, 12.4% FM, 250 g/d FO at ABO, 12.4% FM + 250 g/d FO at ABO | 12.4% FM = 12.4% + 250 g/d FO > CON = 250 g/d FO |
| [10] | 8 | 112 | 4 × 4 Latin square | 0%, 5%, 10%, 15% FM | Linear increase |
| [58] | 16 | 95 | 4 × 4 Latin square | CON, 15% FM + 5% sucrose, 15% FM + 3% FO, 15% FM + 5% sucrose + 3% FO | 15% FM + 5% sucrose = 15% FM + 3% FO = 15% FM + 5% sucrose + 3% FO > CON |
| [19] | 16 | 101 | 4 × 4 Latin square | Different GRC to LM ratios (12:0, 8:4, 4:8, and 0:12) + 15% FM | Tendency for cubic effect |
| Item | Flax Products | ||
|---|---|---|---|
| Flaxseed Hulls 2 (n = 5) | Flaxseed Meal 3 (n = 6) | Whole Flaxseed 4 (n = 1) | |
| Crude protein | 22.4 ± 2.41 | 37.2 ± 1.35 | 23.5 |
| Neutral detergent fiber | 20.6 ± 2.64 | 30.6 ± 4.61 | 20.7 |
| Acid detergent fiber | 15.8 ± 3.44 | 15.9 ± 1.39 | 13.7 |
| Crude fat | 28.4 ± 3.09 | 3.70 ± 4.11 | 34.9 |
| SDG | 1.00 ± 0.08 | 1.60 ± 0.21 | 0.60 |
| Item 3 | Treatments and References 2 | |||||
|---|---|---|---|---|---|---|
| Non-FH vs. 9.88% FH Diets [56] | 0%, 5%, 10%, 15% FM [99] | Non-FM vs. 16% FM Diets [18] | Non-FM vs. 13.7% FM Diets [100] | Non-FM vs. 12.4% FM Diets [98] | CON vs. 7.7% WF, 7.7% WL [108] | |
| Plasma 4 | ||||||
| CAT | NS | NS 8 | − | NS | ↓ | − |
| GPX | NS | NS 9 | NS | NS | NS | − |
| SOD | NS | NS | NS | NS | NS | − |
| Erythrocytes 5 | ||||||
| CAT | NS | NS | − | NS | ↑, † | − |
| GPX | NS | NS | − | NS | NS | − |
| SOD | NS | NS | − | NS | NS | − |
| Mammary tissue 6 | ||||||
| CAT | NS | NS | − | NS | NS | − |
| GPX | NS | NS | − | NS | NS | − |
| SOD | ↑, † | NS | − | NS | NS | − |
| Hepatic tissue 7 | ||||||
| CAT | − | − | − | − | − | NS |
| GPX | − | − | − | − | − | NS |
| SOD | − | − | − | − | − | NS |
| Items 3 | Treatments 2 and References | |||
|---|---|---|---|---|
| Non-FH vs. 9.88% FH diets [56] | 0%, 5%, 10%, 15% FM [99] | Non-FM vs. 13.7% FM diets [100] | Non-FM vs. 12.4% FM diets [98] | |
| CAT | ↑ | linear increase† | NS | − |
| GPX1 | ↑ | NS | NS | NS |
| GPX3 | ↓ | NS | NS | − |
| SOD1 | ↑ | NS | NS | − |
| SOD2 | ↓ | NS | NS | − |
| SOD3 | ↓ | NS | NS | − |
| NFE2L2 | − | linear increase | − | − |
| NF-κB | − | NS | − | − |
| NF-κB1 | − | − | ↓, † | NS |
| NRF2 | − | − | NS | NS |
| Items 3 | Treatments 2 and References | ||
|---|---|---|---|
| 0%, 5%, 10%, 15% FM [99] | Non-FM vs. 12.4% FM Diets [111] | Non-FM vs. 13.7% FM Diets [112] | |
| Milk TBARS | Q,C 4 | NS | NS |
| Plasma TBARS | NS | NS | NS |
| Ruminal TBARS | NS 5 | NS 6 | ↑ 7 |
| Plasma peroxidizability index | − | NS | NS |
| Plasma total antioxidant capacity | − | NS | NS |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brito, A.F.; Zang, Y. A Review of Lignan Metabolism, Milk Enterolactone Concentration, and Antioxidant Status of Dairy Cows Fed Flaxseed. Molecules 2019, 24, 41. https://doi.org/10.3390/molecules24010041
Brito AF, Zang Y. A Review of Lignan Metabolism, Milk Enterolactone Concentration, and Antioxidant Status of Dairy Cows Fed Flaxseed. Molecules. 2019; 24(1):41. https://doi.org/10.3390/molecules24010041
Chicago/Turabian StyleBrito, André F., and Yu Zang. 2019. "A Review of Lignan Metabolism, Milk Enterolactone Concentration, and Antioxidant Status of Dairy Cows Fed Flaxseed" Molecules 24, no. 1: 41. https://doi.org/10.3390/molecules24010041
APA StyleBrito, A. F., & Zang, Y. (2019). A Review of Lignan Metabolism, Milk Enterolactone Concentration, and Antioxidant Status of Dairy Cows Fed Flaxseed. Molecules, 24(1), 41. https://doi.org/10.3390/molecules24010041