
From Fish Eggs to Fish Name: Caviar Species Discrimination by COIBar-RFLP, an Efficient Molecular Approach to Detect Fraud in the Caviar Trade



Abstract
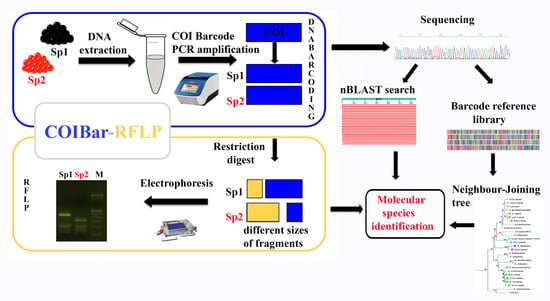
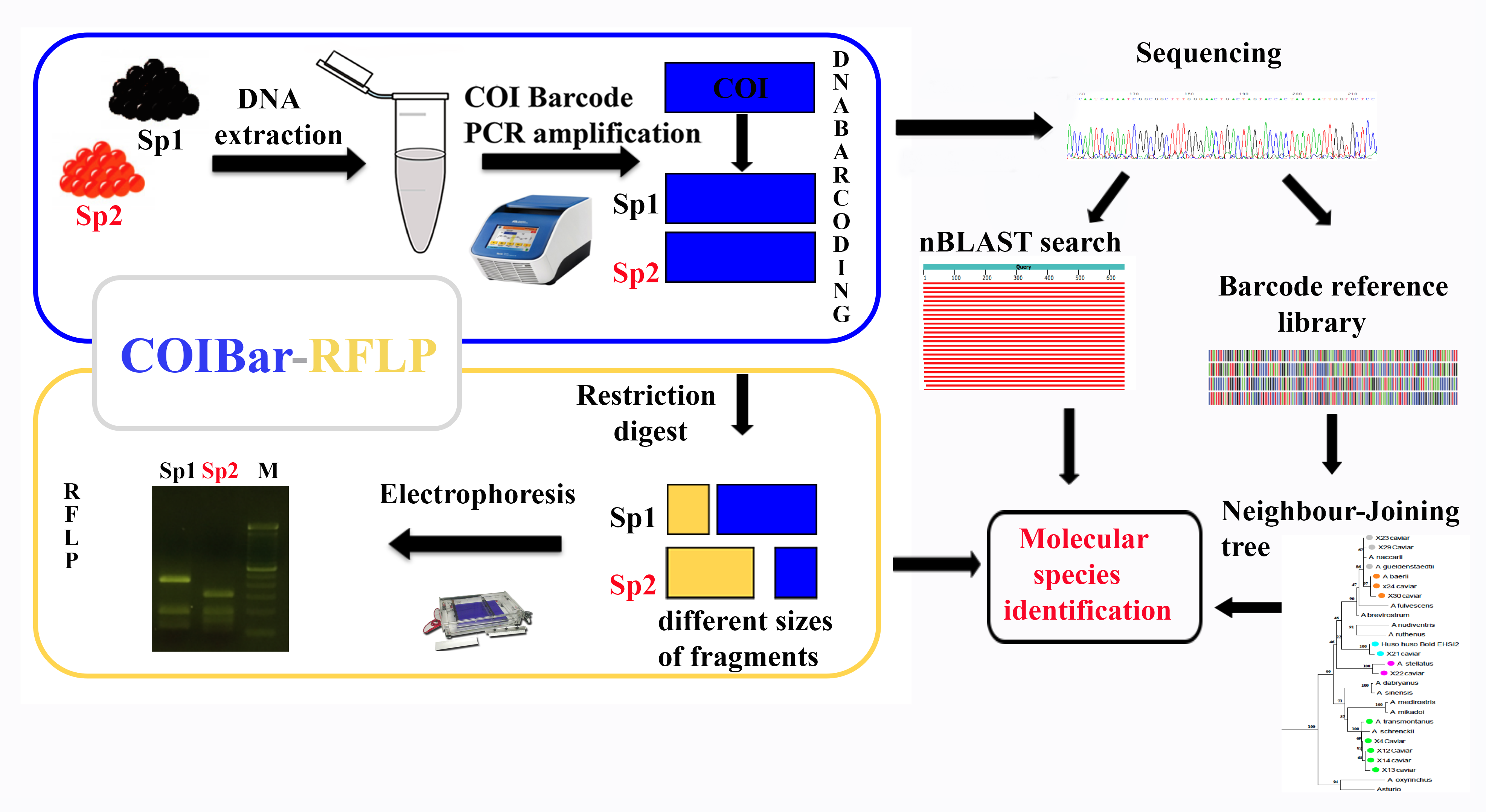
:
1. Introduction
2. Results
2.1. COI Sequences
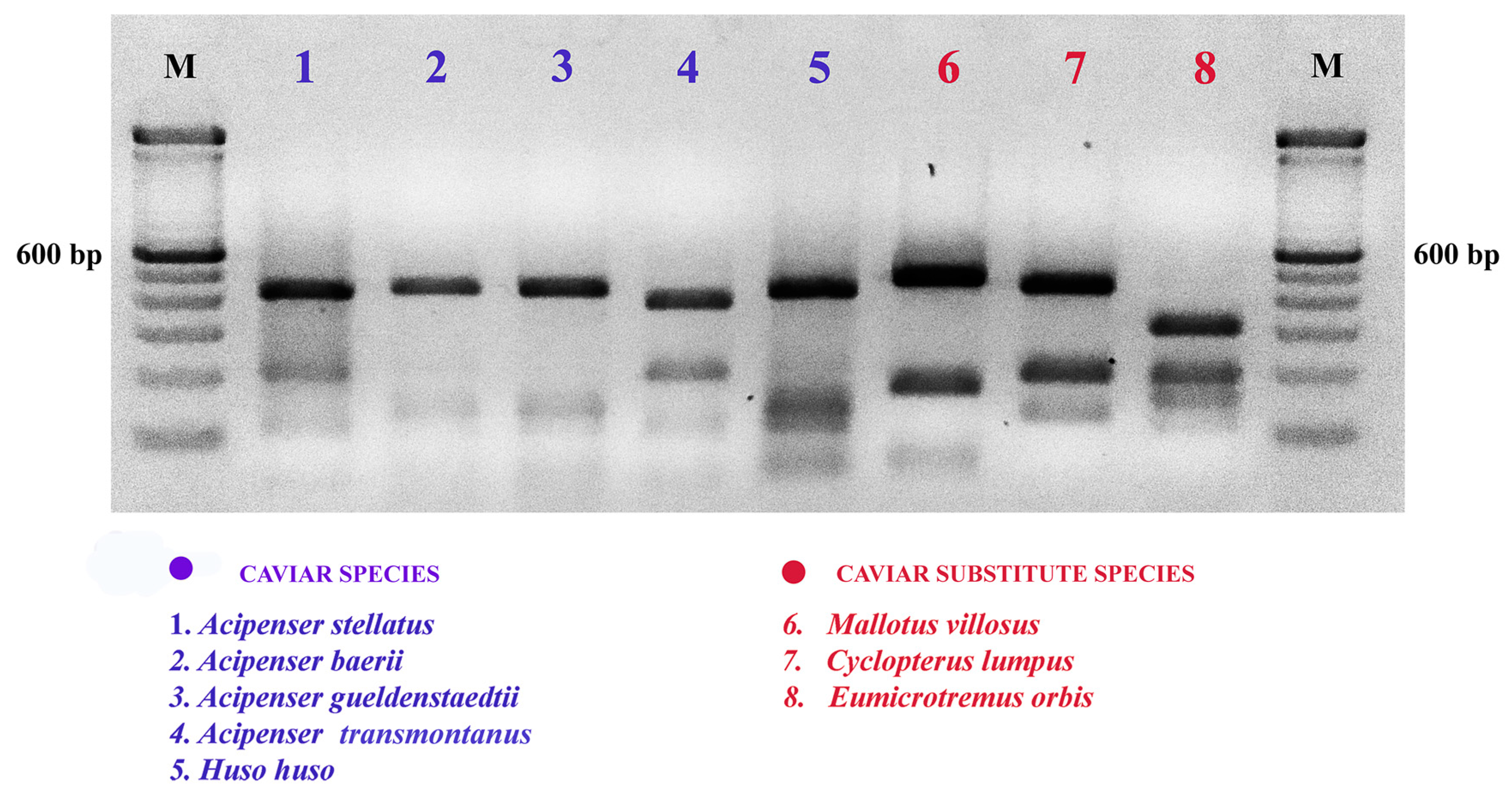
2.2. COIBar-RFLP
3. Discussion
4. Materials and Methods
4.1. Sampling
4.2. COI Barcode Amplification, Sequencing and Data Analysis
4.3. COIBar-RFLP
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Raymakers, C. CITES, the convention on international trade in endangered species of wild Fauna and Flora: Its role in the conservation of Acipenseriformes. J. Appl. Ichthyol. 2006, 22, 53–65. [Google Scholar] [CrossRef]
- CAC, Codex Alimentarius Commission. Codex Stan 291-2010. Standard for Sturgeon Caviar. Available online: www.fao.org (accessed on 5 July 2019).
- Bronzi, P.; Rosenthal, H. Present and future sturgeon and caviar production and marketing: A global market overview. J. Appl. Ichthyol. 2014, 30, 1536–1546. [Google Scholar] [CrossRef]
- EUMOFA, The caviar market. Production, trade and consumption in and outside the EU. Available online: www.eumofa.eu (accessed on 5 July 2019).
- Pappalardo, A.M.; Guarino, F.; Reina, S.; Messina, A.; De Pinto, V. Geographically widespread swordfish barcode stock identification: A case study of its application. PLoS ONE 2011, 6, e25516. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Ferrito, V. DNA barcoding species identification unveils mislabeling of processed flatfish products in southern Italy markets. Fish. Res. 2015, 164, 153–158. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Copat, C.; Ferrito, V.; Grasso, A.; Ferrante, M. Heavy metal content and molecular species identification in canned tuna: Insights into human food safety. Mol. Med. Rep. 2017, 15, 3430–3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Roy. Soc. Lond. B. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; deWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergence, among closely related species. Proc. Roy. Soc. Lond. B 2003, 270, S96–S99. [Google Scholar] [CrossRef]
- Paquin, R.; Hedin, M. The power and perils of ‘molecular taxonomy’: A case study of eyeless and endangered Cicurina (Araneae: Dictynidae) from Texas caves. Mol. Ecol. 2004, 13, 3239–3255. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Phil. Trans. Roy. Soc. B. 2005, 360, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Hajibabaei, M.; Smith, M.A.; Janzen, D.H.; Rodriguez, J.J.; Whitfield, J.B.; Hebert, P.D.N. A minimalist barcode can identify a specimen whose DNA is degraded. Mol. Ecol. Notes 2006, 6, 959–964. [Google Scholar] [CrossRef]
- Lefebure, T.; Douady, C.J.; Gouy, M.; Gibert, J. Relationship between morphological taxonomy and molecular divergence within Crustacea: Proposal of a molecular threshold to help species delimitation. Mol. Phyl. Evol. 2006, 40, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Benard-Capelle, J.; Guillonneau, V.; Nouvian, C.; Fournier, N.; Le Loët, K.; Dettai, A. Fish mislabeling in France: Substitution rates and retail types. PeerJ 2015, 2, e714. [Google Scholar] [CrossRef] [PubMed]
- Pappalardo, A.M.; Cuttitta, A.; Sardella, A.; Musco, M.; Maggio, T.; Patti, B.; Mazzola, S.; Ferrito, V. DNA barcoding and COI sequence variation in Mediterranean lanternfishes larvae. Hydrobiologia 2015, 745, 155–167. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Federico, C.; Sabella, G.; Saccone, S.; Ferrito, V. A COI nonsynonymous mutation as diagnostic tool for intraspecific discrimination in the European Anchovy Engraulis encrasicolus (Linnaeus). PLoS ONE 2015, 10, e0143297. [Google Scholar] [CrossRef] [PubMed]
- Vitale, D.G.M.; Viscuso, R.; D’Urso, V.; Gibilras, S.; Sardella, A.; Marletta, A.; Pappalardo, A.M. Morphostructural analysis of the male reproductive system and DNA barcoding in Balclutha brevis Lindberg 1954 (Homoptera, Cicadellidae). Micron 2015, 79, 36–45. [Google Scholar] [CrossRef]
- Christiansen, H.; Dettai, A.; Heindler, F.M.; Collins, M.A.; Duhamel, G.; Hautecoeur, M.; Steinke, D.; Volckaert, A.M.; Van de Putte, A.P. Diversity of Mesopelagic fishes in the Southern Ocean–A phylogeographic perspective using DNA barcoding. Front. Ecol. Evol. 2018, 6, 120. [Google Scholar] [CrossRef]
- Conti, E.; Mulder, C.; Pappalardo, A.M.; Ferrito, V.; Costa, G. How soil granulometry, temperature and water predict genetic differentiation in namibian Ariadna spiders and explain their behaviour. Ecol. Evol. 2019, 9, 4382–4391. [Google Scholar] [CrossRef]
- Dawnay, N.; Ogden, R.; McEwing, R.; Carvalho, G.R.; Thorpe, R.S. Validation of the barcoding gene COI for use in forensic genetic species identification. Forensic Sci. Int. 2007, 173, 1–6. [Google Scholar] [CrossRef]
- Kappel, K.; Schröder, U. Substitution of high-priced fish with low-priced species: Adulteration of common sole in German restaurants. Food Control 2016, 59, 478–486. [Google Scholar] [CrossRef]
- Pardo, M.A.; Jimenez, E.; Viđarsson, J.R.; Olafsson, K.; Olafsdottir, G.; Danielsdottir, A.K.; Perez-Villareal, B. DNA barcoding revealing mislabeling of seafood in European mass caterings. Food Control 2018, 92, 7–16. [Google Scholar] [CrossRef]
- Sultana, S.; Ali, M.E.; Hossain, M.A.M.; Asing Naquiah, N.; Zaidul, I.S.M. Universal mini COI barcode for the identification of fish species in processed products. Food Res. Int. 2018, 105, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Yao, L.; Ying, X.; Lu, L.; Guardone, L.; Armani, A.; Guidi, A.; Xiong, X. Multiple fish species identified from China’s roasted Xue Yu fillet products using DNA and mini-DNA barcoding: Implications on human health and marine sustainability. Food Control 2018, 88, 123–130. [Google Scholar] [CrossRef]
- Aranishi, F.; Okimoto, T.; Izumi, S. Identification of gadoid species (Pisces, Gadidae) by PCR-RFLP analysis. J. Appl. Genet. 2005, 46, 69–73. [Google Scholar]
- Hsieh, C.; Chang, W.; Chang, H.C.; Hsieh, H.; Chung, Y.; Hwang, D. Puffer fish-based commercial fraud identification in a segment of cytochrome b region by PCR-RFLP analysis. Food Chem. 2010, 121, 1305–1311. [Google Scholar] [CrossRef]
- Rea, S.; Storani, G.; Mascaro, N.; Stocchi, R.; Loschi, A.R. Species identification in anchovy pastes from the market by PCR-RFLP technique. Food Control 2009, 20, 515–520. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, Y.; Li, H.; Wang, J.; Chen, W.; Zhou, Y.; Zhou, S. Differentiation of fish species in Taiwan Strait by PCR-RFLP and lab-on-a-chip system. Food Control 2014, 44, 26–34. [Google Scholar] [CrossRef]
- Pappalardo, A.M.; Ferrito, V. A COIBar-RFLP strategy for the rapid detection of Engraulis encrasicolus in processed anchovy products. Food Control 2015, 57, 385–392. [Google Scholar] [CrossRef]
- Ferrito, V.; Bertolino, V.; Pappalardo, A.M. White fish authentication by COIBar-RFLP: Toward a common strategy for the rapid identification of species in convenience seafood. Food Control 2016, 70, 130–137. [Google Scholar] [CrossRef]
- Ferrito, V.; Pappalardo, A.M. Seafood species identification by DNA barcoding, a molecular tool for food traceability. Biodivers. J. 2017, 8, 65–72. [Google Scholar]
- Pappalardo, A.M.; Federico, C.; Saccone, S.; Ferrito, V. Differential flatfish species detection by COIBar-RFLP in processed seafood products. Eur. Food Res. Technol. 2018, 244, 2191–2201. [Google Scholar] [CrossRef]
- Ludwig, A. Identification of Acipenseriformes species in trade. J. Appl. Ichthyol. 2008, 24, 2–19. [Google Scholar] [CrossRef]
- Zhang, D.X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. 1996, 11, 247–251. [Google Scholar] [CrossRef]
- Birstein, V.J.; Doukakis, P.; Sorkin, B.; DeSalle, R. Population aggregation analysis of three caviar-producing species of sturgeons and implications for the species identification of black caviar. Conserv. Biol. 1998, 12, 766–775. [Google Scholar] [CrossRef]
- Bronzi, P.; Rosenthal, H.; Gessner, J. Global sturgeon aquaculture production: An overview. J. Appl. Ichthyol. 2011, 27, 169–175. [Google Scholar] [CrossRef]
- Ludwig, A.; Lieckfeldt, D.; Jahrll, J. Mislabeled and counterfeit sturgeon caviar from Bulgaria and Romania. J. App. Ichthyol. 2015, 31, 587–591. [Google Scholar] [CrossRef]
- Fain, S.R.; Straughan, D.J.; Hamlin, B.C.; Hoesch, R.M.; LeMay, J.P. Forensic genetic identification of sturgeon caviars traveling in world trade. Conserv. Genet. 2013, 14, 855–874. [Google Scholar] [CrossRef]
- Sicuro, B. The future of caviar production on the light of social changes: A new dawn for caviar? Rev. Aquac. 2019, 11, 204–219. [Google Scholar] [CrossRef]
- Teletchea, F. Molecular identification methods of fish species: Reassessment and possible applications. Rev. Fish. Biol. Fish. 2009, 19, 265–293. [Google Scholar] [CrossRef]
- Ludwig, A.; Debus, L.; Jenneckens, I. A molecular approach to control the international trade in black caviar. Int. Rev. Hydrob. 2002, 87, 661–674. [Google Scholar] [CrossRef]
- Trocchia, S.; Rabbito, D.; D’Angelo, R.; Ciarcia, G.; Abdel-Gawad, F.K.; Guerriero, G. Blue economy and biodiversity surveillance: Fish caviar substitute rapid discrimination. Biodivers. Endanger. Species 2015, S1, 2. [Google Scholar] [CrossRef]
- Behrens-Chapuis, S.; Malewski, T.; Suchecka, E.; Geiger, M.F.; Herder, F.; Bogdanowicz, W. Discriminating European cyprinid specimens by barcode high-resolution melting analysis (Bar-HRM)–A cost efficient and faster way for specimen assignment? Fish. Res. 2018, 204, 61–73. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D.N. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 225–364. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Lago, F.C.; Alonso, M.; Vieites, J.M.; Espiñeira, M. Fish and Seafood Authenticity–Species Identification. In Seafood Processing: Technology, Quality and Safety; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2014. [Google Scholar]
Sample Availability: Not available. |


{kind=link}
{kind=link}
{kind=link}
| Species | Common Name | GenBank Accession Number | References |
|---|---|---|---|
| Acipenser baerii | Siberian sturgeon | KP833625 | Unpublished |
| Acipenser brevirostrum | Shortnose sturgeon | EU523870 | Hubert et al. 2008 |
| Acipenser dabryanus | Yangtze sturgeon | KP218558 | Li et al. 2015 |
| Acipenser fulvescens | Lake sturgeon | EU524392 | Hubert et al. 2008 |
| Acipenser gueldenstaedtii | Russian sturgeon | JQ623904 | Unpublished |
| Acipenser medirostris | Green sturgeon | EU523879 | Hubert et al. 2008 |
| Acipenser mikadoi | Sakhalin sturgeon | NC031188 | Unpublished |
| Acipenser naccarii | Adriatic sturgeon | KJ552424 | Geiger et al. 2014 |
| Acipenser nudiventris | Ship sturgeon | JQ 623905 | Unpublished |
| Acipenser oxyrinchus | Atlantic sturgeon | KX145066 | Unpublished |
| Acipenser persicus | Persian sturgeon | FJ809724 | Unpublished |
| Acipenser ruthens | Sterlet | HQ960576 | Unpublished |
| Acipenser schrenckii | Amur sturgeon | KP218530 | Li et al. 2015 |
| Acipenser sinensis | Chinese sturgeon | KP218545 | Li et al. 2015 |
| Acipenser stellatus | Stellate sturgeon | KC500131 | Keskin & Atar 2013 |
| Acipenser sturio | Common sturgeon | KJ552406 | Geiger et al. 2014 |
| Acipenser transmontanus | White sturgeon | EU523889 | Hubert et al. 2008 |
| Huso huso | Beluga sturgeon | EHSI214-06 * | Unpublished |
| Mallotus villosus | Capelin | HQ712650 | Macklenburg et al. 2011 |
| Cyclopterus lumpus | Lumpfish | MG421955 | Unpublished |
| Eumicrotremus orbis | Pacific spiny lumpsucker) | JQ354090 | Unpublished |
| Code | Processed Fish Product | n° Sample | Declared Species | GenBank Access N° | Species Matched by BLAST | Matched Accession from BLAST | % Identity |
|---|---|---|---|---|---|---|---|
| X1 | capelin | 3 | Mallotus villosus | MK903699 | Mallotus villosus | FJ205579 | 99.70 |
| X2 | capelin | 3 | Mallotus villosus | MK903700 | Mallotus villosus | FJ205579 | 99.24 |
| X3 | lumpfish | 3 | Cyclopterus lumpus | MK903701 | Cyclopterus lumpus | KJ204826 | 99.85 |
| X4 | white caviar | 3 | Acipenser transmontanus | MK903717 | Acipenser transmontanus | KX145032 | 99.85 |
| X5 | black capelin | 3 | Mallotus villosus | MK903702 | Mallotus villosus | FJ205579 | 99.39 |
| X6 | lumpfish | 3 | Cyclopterus lumpus | MK903703 | Cyclopterus lumpus | KJ204826 | 99.54 |
| X7 | capelin | 3 | Mallotus villosus | MK903704 | Mallotus villosus | FJ205579 | 99.09 |
| X8 | caviar | 3 | not specified | MK903718 | Eumicrotremus orbis | JQ354090 | 96.91 |
| X9 | caviar | 3 | not specified | MK903705 | Mallotus villosus | FJ205579 | 99.55 |
| X10 | red capelin | 3 | Mallotus villosus | MK903706 | Mallotus villosus | FJ205579 | 99.09 |
| X11 | black capelin | 3 | Mallotus villosus | MK903706 | Mallotus villosus | FJ205579 | 99.09 |
| X12 | white caviar | 3 | Acipenser transmontanus | MK903719 | Acipenser transmontanus | KX145032 | 99.85 |
| X13 | white caviar | 3 | Acipenser transmontanus | MK903720 | Acipenser transmontanus | KX145032 | 99.54 |
| X14 | white caviar | 3 | Acipenser transmontanus | MK903719 | Acipenser transmontanus | KX145032 | 99.85 |
| X15 | capelin | 3 | Mallotus villosus | MK903707 | Mallotus villosus | FJ205579 | 98.94 |
| X16 | lumpfish | 3 | Cyclopterus lumpus | MK903708 | Cyclopterus lumpus | KJ204826 | 99.69 |
| X17 | lumpfish | 3 | Cyclopterus lumpus | MK903709 | Cyclopterus lumpus | KJ204826 | 99.38 |
| X18 | red capelin | 3 | Mallotus villosus | MK903710 | Mallotus villosus | FJ205581 | 98.94 |
| X19 | caviar | 3 | not specified | MK903711 | Mallotus villosus | FJ205581 | 98.79 |
| X20 | capelin | 3 | Mallotus villosus | MK903712 | Mallotus villosus | FJ205579 | 98.79 |
| X21 | beluga caviar | 2 | Huso huso | MK903721 | Huso huso | AY442351 | 99.69 |
| X22 | sevruga caviar | 2 | Acipenser stellatus | MK903722 | Acipenser stellatus | HQ960585 | 99.08 |
| X23 | oscietra caviar | 2 | Acipenser gueldenstaedtii | MK903723 | Acipenser gueldenstaedtii | KC500088 | 99.85 |
| X24 | baeri caviar | 3 | Acipenser baerii | MK903724 | Acipenser baerii | KM286420 | 99.85 |
| X25 | caviar | 3 | not specified | MK903713 | Mallotus villosus | FJ205581 | 98.49 |
| X26 | red capelin | 3 | Mallotus villosus | MK903714 | Mallotus villosus | FJ205579 | 98.64 |
| X27 | lumpfish | 3 | Cyclopterus lumpus | MK903715 | Cyclopterus lumpus | KJ204826 | 99.54 |
| X28 | lumpfish | 3 | Cyclopterus lumpus | MK903709 | Cyclopterus lumpus | KJ204826 | 99.38 |
| X29 | oscietra caviar | 3 | Acipenser gueldenstaedtii | MK903725 | Acipenser gueldenstaedtii | KC500088 | 99.85 |
| X30 | baeri caviar | 3 | Acipenser baerii | MK903726 | Acipenser baerii | KM286420 | 99.69 |
| X31 | lumpfish | 3 | Cyclopterus lumpus | MK903716 | Cyclopterus lumpus | KJ204826 | 99.38 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pappalardo, A.M.; Petraccioli, A.; Capriglione, T.; Ferrito, V. From Fish Eggs to Fish Name: Caviar Species Discrimination by COIBar-RFLP, an Efficient Molecular Approach to Detect Fraud in the Caviar Trade. Molecules 2019, 24, 2468. https://doi.org/10.3390/molecules24132468
Pappalardo AM, Petraccioli A, Capriglione T, Ferrito V. From Fish Eggs to Fish Name: Caviar Species Discrimination by COIBar-RFLP, an Efficient Molecular Approach to Detect Fraud in the Caviar Trade. Molecules. 2019; 24(13):2468. https://doi.org/10.3390/molecules24132468
Chicago/Turabian StylePappalardo, Anna Maria, Agnese Petraccioli, Teresa Capriglione, and Venera Ferrito. 2019. "From Fish Eggs to Fish Name: Caviar Species Discrimination by COIBar-RFLP, an Efficient Molecular Approach to Detect Fraud in the Caviar Trade" Molecules 24, no. 13: 2468. https://doi.org/10.3390/molecules24132468
APA StylePappalardo, A. M., Petraccioli, A., Capriglione, T., & Ferrito, V. (2019). From Fish Eggs to Fish Name: Caviar Species Discrimination by COIBar-RFLP, an Efficient Molecular Approach to Detect Fraud in the Caviar Trade. Molecules, 24(13), 2468. https://doi.org/10.3390/molecules24132468