Ampelopsin E Reduces the Invasiveness of the Triple Negative Breast Cancer Cell Line, MDA-MB-231

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Cytotoxicity of Ampelopsin E
2.2. Rate of Migration of MDA-MB-231 Cells
2.3. Cell Transmigration and Invasion
2.4. Invadopodia Formation and Gelatin Degradation
2.5. Proteins Involved in Invadopodia Formation
3. Discussion
4. Materials and Methods
4.1. Ampelopsin E
4.2. Cell Culture
4.3. Cytotoxicity of Ampelopsin E towards MDA-MB-231 Cells
4.4. Migration of MDA-MB-231 Cells
4.5. Transmigration and Invasion of MDA-MB-231 Cells
4.6. Preparation of Gelatin-Coated Sterile Coverslip
4.7. Invadopodia Detection
4.8. Gelatin Degradation
4.9. Detection of Matrix Metalloproteases and Platelet-Derived Growth Factor
4.10. Statistical Analysis
5. Conclusions
6. Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| ANOVA | Analysis of variance |
| BHMC | 2,6-bis-(-4-Hydroxy-3-methoxybenzylidine) cyclohexane |
| D. | Dryobalanops |
| DMSO | Dimethyl sulfoxide |
| Doxo | Doxorubicin |
| ECM | Extracellular matrix |
| EMT | Epithelial-to-mesenchymal transition |
| ER | Estrogen receptor |
| HER-2 or c-erbB2 | Human epidermal growth factor receptor-2 |
| MMP | Matrix metalloprotease |
| MTT assay | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay |
| PDGF | Platelet-derived growth factor |
| PR | Progesterone receptor |
| TNBC | Triple negative breast cancer |
References
- Dubey, A.K.; Gupta, U.; Jain, S. Breast Cancer Statistics and Prediction Methodology: A Systematic Review and Analysis. Asian Pac. J. Cancer Prev. 2015, 16, 4237–4245. [Google Scholar] [CrossRef]
- Tazzite, A.; Jouhadi, H.; Saiss, K.; Benider, A.; Nadifi, S. Relationship Between Family History of Breast Cancer and Clinicopathological Features in Moroccan Patients. Ethiop. J. Health Sci. 2013, 23, 150–157. [Google Scholar]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-tieulent, J.; Jemal, A. Global Cancer Statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef]
- Ferlay, J.; Soerjomataram, I.; Ervik, M.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.; Forman, D.; Bray, F. Globocan 2012 v1.0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11; International Agency for Research on Cancer: Lyon, France; Available online: http://globocan.iarc.fr (accessed on 22 April 2017).
- Lu, X.; Kang, Y. Organotropism of Breast Cancer Metastasis. J. Mammary Gland Biol. Neoplasia 2007, 12, 153–162. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Reis-Filho, J.S.; Simpson, P.T.; Gale, T.; Lakhani, S.R. The Molecular Genetics of Breast Cancer: The Contribution of Comparative Genomic Hybridization. Pathol. Res. Pract. 2005, 201, 713–725. [Google Scholar] [CrossRef]
- Simpson, P.T.; Reis-Filho, J.S.; Gale, T.; Lakhani, S.R. Molecular Evolution of Breast Cancer. J. Pathol. 2005, 205, 248–254. [Google Scholar] [CrossRef]
- Lacroix, M.; Toillon, R.A.; Leclercq, G. Stable “portrait” of Breast Tumors during Progression: Data from Biology, Pathology and Genetics. Endocr. Relat. Cancer 2004, 11, 497–522. [Google Scholar] [CrossRef]
- Kabir, M.H.; Suh, E.J.; Lee, C. Comparative Phosphoproteome Analysis Reveals More ERK Activation in MDA-MB-231 than in MCF-7. Int. J. Mass Spectrom. 2012, 309, 1–12. [Google Scholar] [CrossRef]
- Collignon, J.; Lousberg, L.; Schroeder, H.; Jerusalem, G. Triple-Negative Breast Cancer: Treatment Challenges and Solutions. Breast Cancer Targets Ther. 2016, 8, 93–107. [Google Scholar]
- Reis-Filho, J.S.; Tutt, A.N.J. Triple Negative Tumours: A Critical Review. Histopathology 2008, 52, 108–118. [Google Scholar] [CrossRef]
- Brady-West, D.C.; McGrowder, D.A. Triple Negative Breast Cancer: Therapeutic and Prognostic Implications. Asian Pac. J. Cancer Prev. 2011, 12, 2139–2143. [Google Scholar]
- Foulkes, W.; Smith, I.; Reis-Filho, J. Triple-Negative Breast Cancer. N. Engl. J. Med. 2010, 363, 1938–1948. [Google Scholar] [CrossRef]
- Brenton, J.D.; Carey, L.A.; Ahmed, A.A.; Caldas, C. Molecular Classification and Molecular Forecasting of Breast Cancer: Ready for Clinical Application? J. Clin. Oncol. 2005, 23, 7350–7360. [Google Scholar] [CrossRef]
- Viale, G.; Rotmensz, N.; Maisonneuve, P.; Bottiglieri, L.; Montagna, E.; Luini, A.; Veronesi, P.; Intra, M.; Torrisi, R.; Cardillo, A.; et al. Invasive Ductal Carcinoma of the Breast with the “Triple-Negative” Phenotype: Prognostic Implications of EGFR Immunoreactivity. Breast Cancer Res. Treat. 2009, 116, 317–328. [Google Scholar] [CrossRef]
- Bauer, K.R.; Brown, M.; Cress, R.D.; Parise, C.A.; Caggiano, V. Descriptive Analysis of Estrogen Receptor (ER)-Negative, Progesterone Receptor (PR)-Negative, and HER2-Negative Invasive Breast Cancer, the so-Called Triple-Negative Phenotype: A Population-Based Study from the California Cancer Registry. Cancer 2007, 109, 1721–1728. [Google Scholar] [CrossRef]
- Morris, G.J.; Naidu, S.; Topham, A.K.; Guiles, F.; Xu, Y.; McCue, P.; Schwartz, G.F.; Park, P.K.; Rosenberg, A.L.; Brill, K.; et al. Differences in Breast Carcinoma Characteristics in Newly Diagnosed African-American and Caucasian Patients: A Single-Institution Compilation Compared with the National Cancer Institute’s Surveillance, Epidemiology, and End Results Database. Cancer 2007, 110, 876–884. [Google Scholar] [CrossRef]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [Green Version]
- Carey, L.; Winer, E.; Viale, G.; Cameron, D.; Gianni, L. Triple-Negative Breast Cancer: Disease Entity or Title of Convenience? Nat. Rev. Clin. Oncol. 2010, 7, 683–692. [Google Scholar] [CrossRef]
- Kreike, B.; van Kouwenhove, M.; Horlings, H.; Weigelt, B.; Peterse, H.; Bartelink, H.; van de Vijver, M.J. Gene Expression Profiling and Histopathological Characterization of Triple-Negative/Basal-like Breast Carcinomas. Breast Cancer Res. 2007, 9, 1–14. [Google Scholar] [CrossRef]
- Anders, C.; Carey, L. Biology, Metastatic Patterns, and Treatment of Patients with Triple-Negative Breast Cancer. Clin. Breast Cancer 2009, 9 (Suppl. 11), S73–S81. [Google Scholar] [CrossRef] [PubMed]
- Pistelli, M.; Pagliacci, A.; Battelli, N.; Santinelli, A.; Biscotti, T.; Ballatore, Z.; Berardi, R.; Cascinu, S. Prognostic Factors in Early-Stage Triple-Negative Breast Cancer: Lessons and Limits from Clinical Practice. Anticancer Res. 2013, 33, 2737–2742. [Google Scholar] [PubMed]
- Bianchini, G.; Balko, J.M.; Mayer, I.A.; Sanders, M.E.; Gianni, L. HHS Public Access. Triple-Negat. Breast Cancer Chall. Oppor. Heterog. Dis. 2017, 13, 674–690. [Google Scholar]
- Gibbs, J.B. Mechanism-Based Target Identification and Drug Discovery in Cancer Research. Science 2000, 287, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seca, A.M.L.; Pinto, D.C.G.A. Plant Secondary Metabolites as Anticancer Agents: Successes in Clinical Trials and Therapeutic Application. Int. J. Mol. Sci. 2018, 19, 263. [Google Scholar] [CrossRef]
- Iqbal, J.; Abbasi, B.A.; Mahmood, T.; Kanwal, S.; Ali, B.; Shah, S.A.; Khalil, A.T. Plant-Derived Anticancer Agents: A Green Anticancer Approach. Asian Pac. J. Trop. Biomed. 2017, 7, 1129–1150. [Google Scholar] [CrossRef]
- Kinghorn, A.D.; Chin, Y.-W.; Swanson, S.M. Discovery of Natural Product Anticancer Agents from Biodiverse Organisms. Curr. Opin. Drug Discov. Dev. 2009, 12, 189–196. [Google Scholar]
- Shabani, A. A Review of Anticancer Properties of Herbal Medicines. J. Pharm. Care Health Syst. 2016, 3, 2–5. [Google Scholar] [CrossRef]
- Mukherjee, A.; Basu, S.; Sarkar, N.; Ghosh, A. Advances in Cancer Therapy with Plant Based Natural Products. Curr. Med. Chem. 2001, 8, 1467–1486. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, N.; Chen, J.; Shen, J. Emerging Glycolysis Targeting and Drug Discovery from Chinese Medicine in Cancer Therapy. Evid. Based Complement. Altern. Med. 2012, 2012, 873175. [Google Scholar] [CrossRef]
- Mitra, S.; Dash, R. Natural Products for the Management and Prevention of Breast Cancer. Evid.-Based Complement. Altern. Med. 2018, 2018, 1–23. [Google Scholar] [CrossRef]
- Xue, J.P.; Wang, G.; Zhao, Z.B.; Wang, Q.; Shi, Y. Synergistic Cytotoxic Effect of Genistein and Doxorubicin on Drug-Resistant Human Breast Cancer MCF-7/Adr Cells. Oncol. Rep. 2014, 32, 1647–1653. [Google Scholar] [CrossRef]
- Charalambous, C.; Constantinou, A.I. Equol Enhances Tamoxifen’s Antitumor Effect by Induction of Caspase-Mediated Apoptosis in MCF-7 Breast Cancer Cells. BMC Cancer 2013, 13, 238. [Google Scholar] [CrossRef] [PubMed]
- Greenwell, M.; Rahman, P. Medicinal Plants: Their Use in Anticancer Treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103–4112. [Google Scholar] [PubMed]
- Unnati, S.; Ripal, S.; Sanjeev, A.; Niyati, A. Novel Anticancer Agents from Plant Sources. Chin. J. Nat. Med. 2013, 11, 16–23. [Google Scholar]
- Kapinova, A.; Stefanicka, P.; Kubatka, P.; Zubor, P.; Uramova, S.; Kello, M.; Mojzis, J.; Blahutova, D.; Qaradakhi, T.; Zulli, A.; et al. Are Plant-Based Functional Foods Better Choice against Cancer than Single Phytochemicals? A Critical Review of Current Breast Cancer Research. Biomed. Pharmacother. 2017, 96, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.; Odhav, B.; Bhoola, K.D. Natural Products for Cancer Prevention: A Global Perspective. Pharmacol. Ther. 2003, 99, 1–13. [Google Scholar] [CrossRef]
- King, T.D.; Suto, M.J.; Li, Y. The Wnt/β-Catenin Signaling Pathway: A Potential Therapeutic Target in the Treatment of Triple Negative Breast Cancer. J. Cell. Biochem. 2012, 113, 13–18. [Google Scholar] [CrossRef]
- Poma, P.; Labbozzetta, M.; D’Alessandro, N.; Notarbartolo, M. NF-ΚB Is a Potential Molecular Drug Target in Triple-Negative Breast Cancers. OMICS J. Integr. Biol. 2017, 21, 225–231. [Google Scholar] [CrossRef]
- Speiser, J.J.; Erşahin, Ç.; Osipo, C. The Functional Role of Notch Signaling in Triple-Negative Breast Cancer. Vitam. Horm. 2013, 93, 277–306. [Google Scholar]
- Gordon, V.; Banerji, S. Molecular Pathways: PI3K Pathway Targets in Triple-Negative Breast Cancers. Clin. Cancer Res. 2013, 19, 3738–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, J.G.; O’Shaughnessy, J.A. The Hedgehog Pathway in Triple-Negative Breast Cancer. Cancer Med. 2016, 5, 2989–3006. [Google Scholar] [CrossRef] [PubMed]
- Giltnane, J.M.; Balko, J.M. Rationale for Targeting the Ras/MAPK Pathway in Triple-Negative Breast Cancer. Discov. Med. 2014, 17, 275–283. [Google Scholar] [PubMed]
- Varghese, E.; Samuel, S.M.; Abotaleb, M.; Cheema, S.; Mamtani, R.; Büsselberg, D. The “Yin and Yang” of Natural Compounds in Anticancer Therapy of Triple-Negative Breast Cancers. Cancers 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Jaitak, V. Natural Products as Multidrug Resistance Modulators in Cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Yuan, R.; Hou, Y.; Sun, W.; Yu, J.; Liu, X.; Niu, Y.; Lu, J.J.; Chen, X. Natural Products to Prevent Drug Resistance in Cancer Chemotherapy: A Review. Ann. N. Y. Acad. Sci. 2017, 1401, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Boumendjel, A.; Di Pietro, A.; Dumontet, C.; Barron, D. Recent Advances in the Discovery of Flavonoids and Analogs with High-Affinity Binding to P-Glycoprotein Responsible for Cancer Cell Multidrug Resistance. Med. Res. Rev. 2002, 22, 512–529. [Google Scholar] [CrossRef] [PubMed]
- Purwaningsih, P. Ecological Distribution of Dipterocarpaceae Species in Indonesia. Biodivers. J. Biol. Divers. 2004, 5, 89–95. [Google Scholar] [CrossRef]
- Newman, M.F.; Burges, P.F.; Whitmore, T.C. Manual of Dipterocarps Series (Sumatra, Kalimantan, Jawa to Nuigini); Prosea: Bogor, Indonesia, 1999. [Google Scholar]
- Ashton, P.S. Dipterocarpaceae. In Flora Malesiana, 9th ed.; The Hague: Martinus Nijhoff Publishers: Leiden, The Netherlands, 1983. [Google Scholar]
- Wibowo, A.; Ahmat, N. Chemotaxonomic Significance of Oligostilbenoids Isolated from Dryobalanops in the Taxonomic of Dipterocarpaceae. Biochem. Syst. Ecol. 2015, 59, 31–35. [Google Scholar] [CrossRef]
- Agarwal, V.S. Econmoic Plants of India; Kailash Prakashan: Calcuta, India, 1986. [Google Scholar]
- Symington, C.F. Malayan Forest Record No. 16; Kepong: Kuala Lumpur, Malaysia, 1943. [Google Scholar]
- Ito, T.; Ito, H.; Nehira, T.; Sawa, R.; Iinuma, M. Structure Elucidation of Highly Condensed Stilbenoids: Chiroptical Properties and Absolute Configuration. Tetrahedron 2014, 70, 5640–5649. [Google Scholar] [CrossRef]
- Kim, J.; Ra, M.; Kim, O.; Thi, N.; Phuong, T.; Yoon, J.; Keun, W.; Bae, K.; Wook, K. Amurensin G Inhibits Angiogenesis and Tumor Growth of Tamoxifen-Resistant Breast Cancer via Pin1 Inhibition. Food Chem. Toxicol. 2012, 50, 3625–3634. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Chaipech, S.; Matsuda, H.; Hamao, M.; Umeda, Y.; Sato, H.; Tamura, H.; Kon’I, H.; Ninomiya, K.; Yoshikawa, M.; et al. Antidiabetogenic Oligostilbenoids and 3-Ethyl-4-Phenyl-3,4- Dihydroisocoumarins from the Bark of Shorea Roxburghii. Bioorg. Med. Chem. 2012, 20, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Manshoor, N.; Aizam Ekhmal, E.A.; Qamarusy Syazwan, K.B.; Mohd Shafarin, S.; Manshoor, N. Mass Fragmentation Patterns as Fingerprints in Identification of Known Oligostilbenes in Dryobalanops Spp. Extracts. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 1147–1152. [Google Scholar]
- Hsouna, A.B.; Trigui, M.; Culioli, G.; Blache, Y.; Jaoua, S. Antioxidant Constituents from Lawsonia inermis Leaves: Isolation, Structure Elucidation and Antioxidative Capacity. Food Chem. 2011, 125, 193–200. [Google Scholar] [CrossRef]
- Oshima, Y.; Ueno, Y.; Ampelopsins, D.E.H.; Cis-Ampelopsin, E. Oligostilbenes from Ampelopsis brevipedunculata Var. Hancei Roots. Phytochemistry 1993, 33, 179–182. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant. 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Wibowo, A.; Ahmat, N.; Hamzah, A.S.; Latif, F.A.; Norrizah, J.S.; Khong, H.Y.; Takayama, H. Identification and Biological Activity of Secondary Metabolites from Dryobalanops beccarii. Phytochem. Lett. 2014, 9, 117–122. [Google Scholar] [CrossRef]
- Rahman, N.A.; Yazan, L.S.; Wibowo, A.; Ahmat, N.; Foo, J.B.; Tor, Y.S.; Yeap, S.K.; Razali, Z.A.; Ong, Y.S.; Fakurazi, S. Induction of Apoptosis and G2/M Arrest by Ampelopsin E from Dryobalanops towards Triple Negative Breast Cancer Cells, MDA-MB-231. BMC Complement. Altern. Med. 2016, 16, 1–9. [Google Scholar] [CrossRef]
- Talmadge, J.E.; Fidler, I.J. AACR Centennial Series: The Biology of Cancer Metastasis: Historical Perspective. Cancer Res. 2010, 70, 5649–5669. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Oikawa, T. Membrane Lipids in Invadopodia and Podosomes: Key Structures for Cancer Invasion and Metastasis. Oncotarget 2010, 1, 320–328. [Google Scholar] [CrossRef]
- Chen, W.-T. Proteolytic Activity of Specialized Surface Protrusions Formed at Rosette Contact Sites of Transformed Cells. J. Exp. Zool. 1989, 251, 167–185. [Google Scholar] [CrossRef] [PubMed]
- Tarone, G.; Cirillo, D.; Giancotti, F.G.; Comoglio, P.M.; Marchisio, P.C. Rous Sarcoma Virus-Transformed Fibroblasts Adhere Primarily at Discrete Protrusions of the Ventral Membrane Called Podosomes. Exp. Cell Res. 1985, 159, 141–157. [Google Scholar] [CrossRef]
- Eckert, M.A.; Yang, J. Targeting Invadopodia to Block Breast Cancer Metastasis. Oncotarget 2011, 2, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Huttenlocher, A.; Horwitz, A.R. Integrins in Cell Migration TL-3. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Shukla, Y. Herbal Medicine: Current Status and the Future. Asian Pac. J. Cancer Prev. 2003, 4, 281–288. [Google Scholar] [PubMed]
- Saklani, A.; Kutty, S.K. Plant-Derived Compounds in Clinical Trials. Drug Discov. Today 2008, 13, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Koehn, F.E.; Carter, G.T. The Evolving Role of Natural Products in Drug Discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Feher, M.; Schmidt, J.M. Property Distributions: Differences between Drugs, Natural Products, and Molecules from Combinatorial Chemistry. J. Chem. Inf. Comput. Sci. 2003, 43, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lao, Y.; Zhao, Y.; Qin, J.; Fu, W.; Zhang, Y.; Xu, H. Screening Active Compounds from Garcinia Species Native to China Reveals Novel Compounds Targeting the STAT/JAK Signaling Pathway. BioMed. Res. Int. 2015, 2015, 910453. [Google Scholar]
- Wibowo, A.; Ahmat, N.; Hamzah, A.S.; Low, A.L.M.; Mohamad, S.A.S.; Khong, H.Y.; Sufian, A.S.; Manshoor, N.; Takayama, H. Malaysianol B, an Oligostilbenoid Derivative from Dryobalanops lanceolata. Fitoterapia 2012, 83, 1569–1575. [Google Scholar] [CrossRef]
- Wibowo, A. Phytochemical Study of Dryobalanops from Malaysian Dipterocarpaceae, and Structure-Activity Relationship Studies. Dr. Res. Abstr. 2014, 6, 18. [Google Scholar]
- Wibowo, A.; Ahmat, N.; Hamzah, A.S.; Sufian, A.S.; Ismail, N.H.; Ahmad, R.; Jaafar, F.M.; Takayama, H. Malaysianol A, a New Trimer Resveratrol Oligomer from the Stem Bark of Dryobalanops aromatica. Fitoterapia 2011, 82, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Wibowo, A.; Ahmat, N.; Hamzah, A. Oligostilbenoids from the Stem Bark of Dryobalanops aromatica. Planta Med. 2011, 77, PG9. [Google Scholar] [CrossRef]
- Jingwen, B.; Yaochen, L.; Guojun, Z. Cell Cycle Regulation and Anticancer Drug Discovery. Cancer Biol. Med. 2017, 14, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Gabrielli, B.; Brooks, K.; Pavey, S. Defective Cell Cycle Checkpoints as Targets for Anti-Cancer Therapies. Front. Pharmacol. 2012, 3, 1–6. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef]
- Pistritto, G.; Trisciuoglio, D.; Ceci, C.; Garufi, A.; D’Orazi, G. Apoptosis as Anticancer Mechanism: Function and Dysfunction of Its Modulators and Targeted Therapeutic Strategies. Aging 2016, 8, 603–619. [Google Scholar] [CrossRef] [PubMed]
- Hunter, K.W.; Crawford, N.P.; Alsarraj, J. Mechanisms of Metastasis. Breast Cancer Res. 2008, 10, S2. [Google Scholar] [CrossRef]
- Fidler, I.J. Tumor Heterogeneity and the Biology of Cancer Invasion and Metastasis. Cancer Res. 1978, 38, 2651–2660. [Google Scholar] [CrossRef]
- Makki, J. Diversity of Breast Carcinoma: Histological Subtypes and Clinical Relevance. Clin. Med. Insights Pathol. 2015, 8, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Redig, A.J.; McAllister, S.S. Breast Cancer as a Systemic Disease: A View of Metastasis. J. Int. Medcomput. 2013, 274, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Yates, L.R.; Knappskog, S.; Wedge, D.; Farmery, J.H.R.; Gonzalez, S.; Martincorena, I.; Alexandrov, L.B.; Van Loo, P.; Haugland, H.K.; Lilleng, P.K.; et al. Genomic Evolution of Breast Cancer Metastasis and Relapse. Cancer Cell 2017, 32, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Steeg, P.S. Metastasis Suppressors Alter the Signal Transduction of Cancer Cells. Nat. Rev. Cancer 2003, 3, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.Y.; Zhao, C.; Zhang, N.; Jin, M.N.; Tang, S.A.; Qin, N.; Kong, D.X.; Duan, H.Q. Alkaloids from Pachysandra Terminalis Inhibit Breast Cancer Invasion and Have Potential for Development as Antimetastasis Therapeutic Agents. J. Nat. Prod. 2012, 75, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, S.; Weinberg, R. Tumor Metastasis: Molecular Insights and Evolving Paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madsen, C.D.; Sahai, E. Cancer Dissemination-Lessons from Leukocytes. Dev. Cell 2010, 19, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Bracke, M.E.; Maeseneer, D.D.; Derycke, V.V.M.; Vanhoecke, B.; Wever, O.D.; Depypere, H.T. Cell Motility and Breast Cancer Metastasis. In Cancer Metastasis—Biology Treatment; Springer: Berlin, Germany, 2007; pp. 47–75. [Google Scholar]
- Scully, O.J.; Bay, B.-H.; Yip, G.; Yu, Y. Breast Cancer Metastasis OLIVIA. Cancer Genom. Proteom. 2012, 9, 311–320. [Google Scholar]
- Ryan, J. Effect of Different Fetal Bovine Serum Concentrations on the Replicative Life Span of Cultured Chick Cells. In Vitro 1979, 15, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Menyhárt, O.; Harami-Papp, H.; Sukumar, S.; Schäfer, R.; Magnani, L.; de Barrios, O.; Győrffy, B. Guidelines for the Selection of Functional Assays to Evaluate the Hallmarks of Cancer. Biochim. Et Biophys. Acta Rev. Cancer 2016, 1866, 300–319. [Google Scholar]
- Nyegaard, S.; Christensen, B.; Rasmussen, J.T. An Optimized Method for Accurate Quantification of Cell Migration Using Human Small Intestine Cells. Metab. Eng. Commun. 2016, 3, 76–83. [Google Scholar] [CrossRef]
- Gotsulyak, N.Y.; Kosach, V.R.; Cherednyk, O.V.; Tykhonkova, I.O.; Khoruzhenko, A.I. Optimization of Cell Motility Evaluation in Scratch Assay. Biopolym. Cell 2014, 30, 223–228. [Google Scholar] [CrossRef]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-Dimensional Cell Culture Systems and Their Applications in Drug Discovery and Cell-Based Biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albini, A. Tumor and Endothelial Cell Invasion of Basement Membranes. The Matrigel Chemoinvasion Assay as a Tool for Dissecting Molecular Mechanisms. Pathol. Oncol. Res. 1998, 4, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.S.; Weaver, A.M. A New Role for Cortactin in Invadopodia: Regulation of Protease Secretion. Eur. J. Cell Biol. 2008, 87, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Krakhmal, N.V.; Zavyalova, M.V.; Denisov, E.V.; Vtorushin, S.V.; Perelmuter, V.M. Cancer Invasion: Patterns and Mechanisms. Acta Nat. 2015, 7, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Martin, T.A.; Ye, L.; Sanders, A.J.; Lane, J.; Jiang, W.G. Cancer Invasion and Metastasis: Molecular and Cellular Perspective. In Metastatic Cancer: Clinical Biological Perspectives; Jandial, R., Ed.; Landes Bioscience: Austin, TX, USA, 2013; pp. 1–37. [Google Scholar]
- Rajan, V.E.J.S. Adhesion and Transendothelial Migration of Cancer Cells. Ph.D. Thesis, Universite Grenoble Alpes, Grenoble, France, 2016. [Google Scholar]
- Carr, H.S.; Zuo, Y.; Oh, W.; Frost, J.A. Regulation of Focal Adhesion Kinase Activation, Breast Cancer Cell Motility, and Amoeboid Invasion by the RhoA Guanine Nucleotide Exchange Factor Net1. Mol. Cell Biol. 2013, 33, 2771–2786. [Google Scholar] [CrossRef] [PubMed]
- Ki Choi, S.; Suk Kim, H.; Jin, T.; Kyung Moon, W. LOXL4 Knockdown Enhances Tumor Growth and Lung Metastasis through Collagen-Dependent Extracellular Matrix Changes in Triple-Negative Breast Cancer. Oncotarget 2017, 8, 11977–11989. [Google Scholar] [CrossRef]
- Hashim, N.F. Invadopodia Formation in Breast Cancer. Ph.D. Thesis, Division of Cancer Studies, King’s College London, London, UK, 2013. Unpublished. [Google Scholar]
- Block, M.R.; Badowski, C.; Millon-Fremillon, A.; Bouvard, D.; Bouin, A.P.; Faurobert, E.; Gerber-Scokaert, D.; Planus, E.; Albiges-Rizo, C. Podosome-Type Adhesions and Focal Adhesions, so Alike yet so Different. Eur. J. Cell Biol. 2008, 87, 491–506. [Google Scholar] [CrossRef]
- Kim, M.; Kim, M.; Kim, H.; Kim, Y.; Song, Y. Expression Profiles of Epithelial-Mesenchymal Transition-Associated Proteins in Epithelial Ovarian Carcinoma. Biomed Res. Int. 2014, 2014, 495754. [Google Scholar] [CrossRef]
- Parikh, A.; Lee, C.; Peronne, J.; Marchini, S.; Baccarini, A.; Kolev, V.; Romualdi, C.; Fruscio, R.; Shah, H.; Wang, F.; et al. MicroRNA-181a Has a Critical Role in Ovarian Cancer Progression through the Regulation of the Epithelial-Mesenchymal Transition. Natl. Commun. 2014, 5, 2977. [Google Scholar] [CrossRef]
- Adham, S.; Al Harrasi, I.; Al Haddabi, I.; Al Rashdi, A.; Al Sinawi, S.; Al Maniri, A.; Ba-Omar, T.; Coomber, B. Immunohistological Insight into the Correlation between Neuropilin-1 and Epithelial Mesenchymal Transition Markers in Epithelial Ovarian Cancer. J. Histochem. Cytochem. 2014, 62, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Liu, P.; Ding, F.; Yu, N.; Li, S.; Wang, S.; Zhang, X.; Sun, X.; Chen, Y.; Wang, F.; et al. Ampelopsin Reduces the Migration and Invasion of Ovarian Cancer Cells via Inhibition of Epithelial-to-Mesenchymal Transition. Oncol. Rep. 2015, 33, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Christofori, G. EMT, the Cytoskeleton, and Cancer Cell Invasion. Cancer Metastasis Rev. 2009, 28, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Eckert, M.A.; Lwin, T.M.; Chang, A.T.; Kim, J.; Danis, E.; Ohno-Machado, L.; Yang, J. Twist1-Induced Invadopodia Formation Promotes Tumor Metastasis. Cancer Cell 2011, 19, 372–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala, I.; Baldassarre, M.; Giacchetti, G.; Caldieri, G.; Tete, S.; Luini, A.; Buccione, R. Multiple Regulatory Inputs Converge on Cortactin to Control Invadopodia Biogenesis and Extracellular Matrix Degradation. J. Cell Sci. 2008, 121, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Linder, S. The Matrix Corroded: Podosomes and Invadopodia in Extracellular Matrix Degradation. Trends Cell Biol. 2007, 17, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Buccione, R.; Orth, J.D.; McNiven, M.A. Foot and Mouth: Podosomes, Invadopodia and Circular Dorsal Ruffles. Nat. Rev. Mol. Cell Biol. 2004, 5, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Wu, R.; Li, Y.; Zhang, L.; Tang, X.; Tu, J.; Zhou, W.; Wang, J.; Shou, Q. Safflower Yellow Prevents Pulmonary Metastasis of Breast Cancer by Inhibiting Tumor Cell Invadopodia. Am. J. Chin. Med. 2016, 44, 1491–1506. [Google Scholar] [CrossRef]
- Harun, S.; Israf, D.; Tham, C.; Lam, K.; Cheema, M.; Hashim, N.F.M. The Molecular Targets and Anti-Invasive Effects of 2,6-Bis-(4-Hydroxyl-3methoxybenzylidine) Cyclohexanone or BHMC in MDA-MB-231 Human Breast Cancer Cells. Molecules 2018, 23, 865. [Google Scholar] [CrossRef]
- Misteli, T.; Soutoglou, E. The Emerging Role of Nuclear Architecture in DNA Repair and Genome Maintenance. Nat. Rev. Mol. Cell Biol. 2009, 10, 243–354. [Google Scholar] [CrossRef]
- Nitiss, J.L. Targeting DNA Topoisomerase II in Cancer Chemotherapy. Nat. Rev. Cancer 2009, 9, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Doroshow, J.H. Role of Hydrogen Peroxide and Hydroxyl Radical Formation in the Killing of Ehrlich Tumor Cells by Anticancer Quinones. Proc. Natl. Acad. Sci. USA 1986, 83, 4514–4518. [Google Scholar] [CrossRef] [PubMed]
- Nestal, M.G.; Vasconcelos, F.C.; Delbue, D.; Mognol, G.P.; Sternberg, C.; Viola, J.P.; Maia, R.C. Doxorubicin Induces Cell Death in Breast Cancer Cells Regardless of Survivin and XIAP Expression Levels. Eur. J. Cell Biol. 2013, 92, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Cruet-Hennequart, S.; Prendergast, Á.M.; Shaw, G.; Barry, F.P.; Carty, M.P. Doxorubicin Induces the DNA Damage Response in Cultured Human Mesenchymal Stem Cells. Int. J. Hematol. 2012, 96, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Pichot, C.S.; Hartig, S.M.; Xia, L.; Arvanitis, C.; Monisvais, D.; Lee, F.Y.; Frost, J.A.; Corey, S.J. Dasatinib Synergizes with Doxorubicin to Block Growth, Migration, and Invasion of Breast Cancer Cells. Br. J. Cancer 2009, 101, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Martín-Villar, E.; Borda-D’Agua, B.; Carrasco-Ramirez, P.; Renart, J.; Parsons, M.; Quintanilla, M.; Jones, G.E. Podoplanin Mediates ECM Degradation by Squamous Carcinoma Cells through Control of Invadopodia Stability. Oncogene 2015, 34, 4531–4544. [Google Scholar] [CrossRef]
- Revach, O.-Y.; Weiner, A.; Rechav, K.; Sabanay, I.; Livne, A.; Geiger, B. Mechanical Interplay between Invadopodia and the Nucleus in Cultured Cancer Cells. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef]
- Oikawa, T.; Oyama, M.; Kozuka-Hata, H.; Uehara, S.; Udagawa, N.; Saya, H.; Matsuo, K. Tks5-Dependent Formation of Circumferential Podosomes/Invadopodia Mediates Cell-Cell Fusion. J. Cell Biol. 2012, 197, 553–568. [Google Scholar] [CrossRef]
- Huang, F.; Wang, D.; Yao, Y.; Wang, M. PDGF Signaling in Cancer Progression. Int. J. Clin. Exp. Med. 2017, 10, 9918–9929. [Google Scholar]
- Heldin, C. Targeting the PDGF Signaling Pathway in Tumor Treatment. Cell Commun. Signal. 2013, 11, 1–45. [Google Scholar] [CrossRef]
- Ekpe-Adewuyi, E.; Lopez-Campistrous, A.; Tang, X.; Brindley, D.N.; McMullen, T.P. Platelet Derived Growth Factor Receptor Alpha Mediates Nodal Metastases in Papillary Thyroid Cancer by Driving the Epithelial-Mesenchymal Transition. Oncotarget 2016, 7, 83684–83700. [Google Scholar] [CrossRef] [PubMed]
- Murphy, D.; Courtneidge, S. The’ins’ and’outs’ of Podosomes and Invadopodia: Characteristics, Formation and Function. Nat. Rev. Mol. Cell Biol. 2011, 12, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, D.M.; Medici, D. Signaling Mechanisms of the Epithelial-Mesenchymal Transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Wang, Z.; Kong, D.; Ali, R.; Ali, S.; Banerjee, S.; Sarkar, F.H. Platelet-Derived Growth Factor-D Contributes to Aggressiveness of Breast Cancer Cells by up-Regulating Notch and NF-ΚB Signaling Pathways. Breast Cancer Res. Treat. 2011, 126, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Cai, Y. Cis-Ampelopsin E, a Stilbene Isolated from the Seeds of Paeonias suffruticosa, Inhibits Lipopolysaccharide-Stimulated Nitric Oxide Production in RAW 264.7 Macrophages via Blockade of Nuclear Factor-Kappa B Signaling Pathway. Biol. Pharm. Bull. 2011, 34, 1501–1507. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.; Prekeris, R. The Regulation of MMP Targeting to Invadopodia during Cancer Metastasis. Front. Cell Dev. Biol. 2015, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.F.; Matrisian, L.M. Changing Views of the Role of Matrix Metalloproteinases in Metastasis. J. Natl. Cancer Inst. 1997, 89, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Poincloux, R.; Lizarraga, F.; Chavrier, P. Matrix Invasion by Tumor Cells: A Focus on MT1-MMP Trafficking to Invadopodia. J. Cell Sci. 2009, 122, 3015–3024. [Google Scholar] [CrossRef]
- Polette, M.; Nawrocki-Raby, B.; Gilles, C.; Clavel, C.; Birembaut, P. Tumour Invasion and Matrix Metalloproteinases. Crit. Rev. Oncol. Hematol. 2004, 49, 179–186. [Google Scholar] [CrossRef]
- Lebeau, A.; Nerlich, A.G.; Sauer, U.; Lichtinghagen, R.; Lohrs, U. Tissue Distribution of Major Matrix Metalloproteinases and Their Transcripts in Human Breast Carcinomas. Anticancer Res. 1999, 19, 4257–4264. [Google Scholar]
- Chen, W.T.; Wang, J.Y. Specialized Surface Protrusions of Invasive Cells, Invadopodia and Lamellipodia, Have Differential MT1-MMP, MMP-2, and TIMP-2 Localization. Ann. N. Y. Acad. Sci. 1999, 878, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, H.; Howard, L.; Thompson, E.W.; Sato, H.; Seiki, M.; Yeh, Y. Transmembrane/Cytoplasmic Domain-Mediated Membrane Type1-Matrix Metalloprotease Docking to Invadopodiais Required for Cell Invasion. Proc. Natl. Acad. Sci. USA 1997, 94, 7959–7964. [Google Scholar] [CrossRef] [PubMed]
- Desai, B.; Ma, T.; Chellaiah, M.A. Invadopodia and Matrix Degradation, a New Property of Prostate Cancer Cells during Migration and Invasion. J. Biol. Chem. 2008, 283, 13856–13866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruivo, J.; Francisco, C.; Oliveira, R.; Figueiras, A. The Main Potentialities of Resveratrol for Drug Delivery Systems. Braz. J. Pharm. Sci. 2015, 51, 499–513. [Google Scholar] [CrossRef]
- Walle, T.; Hsieh, F.; DeLegge, M.; Oatis, J., Jr.; Walle, U. High Absorption but Very Low Bioavailabiity of Oral Resveratrol in Humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Smoliga, J.M.; Blanchard, O. Enhancing the Delivery of Resveratrol in Humans: If Low Bioavailability Is the Problem, What Is the Solution? Molecules 2014, 19, 17154–17172. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Fan, J.; Chen, N. Potential Molecular Targets of Ampelopsin in Prevention and Treatment of Cancers. Anti-Cancer Agents Med. Chem. 2018, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.T.; Zheng, Y.; Liu, D.Y. Reversal Effect and Its Mechanism of Ampelopsin on Multidrug Resistance in K562/ADR Cells. China J. Chin. Mater. Med. 2009, 34, 761–765. [Google Scholar]
- Jiang, L.; Zhang, Q.; Ren, H.; Ma, S.; Lu, C.J.; Liu, B.; Liu, J.; Liang, J.; Li, M.; Zhu, R. Dihydromyricetin Enhances the Chemo-Sensitivity of Nedaplatin via Regulation of the P53/Bcl-2 Pathway in Hepatocellular Carcinoma Cells. PLoS ONE 2015, 10, e0124994. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Gebäck, T.; Schulz, M.M.P.M.; Koumoutsakos, P.; Detmar, M. TScratch: A Novel and Simple Software Tool for Automated Analysis of Monolayer Wound Healing Assays. Biotechniques 2009, 46, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Wang, Y.; Gu, L.; Duan, B.; Cui, J.; Zhang, Y.; Chen, Y.; Sun, S.; Dong, J.; Du, J. MICAL1 Controls Cell Invasive Phenotype via Regulating Oxidative Stress in Breast Cancer Cells. BMC Cancer 2016, 16, 1–11. [Google Scholar] [CrossRef]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro Cell Migration and Invasion Assays. J. Vis. Exp. 2014, 1–8. [Google Scholar] [CrossRef]
- Hashim, N.F.M.; Nicholas, N.S.; Dart, A.E.; Kiriakidis, S.; Paleolog, E.; Wells, C.M. Hypoxia-Induced Invadopodia Formation: A Role for -PIX. Open Biol. 2013, 3, 120159. [Google Scholar] [CrossRef] [PubMed]
- Martin, K.H.; Hayes, K.E.; Walk, E.L.; Ammer, A.G.; Markwell, S.M.; Weed, S.A. Quantitative Measurement of Invadopodia-Mediated Extracellular Matrix Proteolysis in Single and Multicellular Contexts. J. Vis. Exp. 2012, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, S.K.; Cabrera, R.; Mao, S.P.H.; Christin, J.R.; Wu, B.; Guo, W.; Bravo-Cordero, J.J.; Condeelis, J.S.; Segall, J.E.; Hodgson, L. Rac3 Regulates Breast Cancer Invasion and Metastasis by Controlling Adhesion and Matrix Degradation. J. Cell Biol. 2017, 216, 4331–4349. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.D.; Bratoeva, M.; Toole, B.P. Regulation of Invadopodia Formation and Activity by CD147. J. Cell Sci. 2012, 125, 777–788. [Google Scholar] [CrossRef]
- Sedgwick, A.E.; Clancy, J.W.; Olivia Balmert, M.; D’Souza-Schorey, C. Extracellular Microvesicles and Invadopodia Mediate Non-Overlapping Modes of Tumor Cell Invasion. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Díaz, B. Invadopodia Detection and Gelatin Degradation Assay. Bio-Protocol 2013, 3, 1–7. [Google Scholar]
- Wang, B.; Xing, Z.; Wang, F.; Yuan, X.; Zhang, Y. Fangchinoline Inhibits Migration and Causes Apoptosis of Human Breast Cancer MDA-MB-231 Cells. Oncol. Lett. 2018, 14, 5307–5312. [Google Scholar] [CrossRef]
Sample Availability: Not Available. |



 ) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with * indicated p < 0.05 and bar with ** indicated p < 0.01 when compared to untreated group.
) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with * indicated p < 0.05 and bar with ** indicated p < 0.01 when compared to untreated group.
) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with * indicated p < 0.05 and bar with ** indicated p < 0.01 when compared to untreated group.
) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with * indicated p < 0.05 and bar with ** indicated p < 0.01 when compared to untreated group.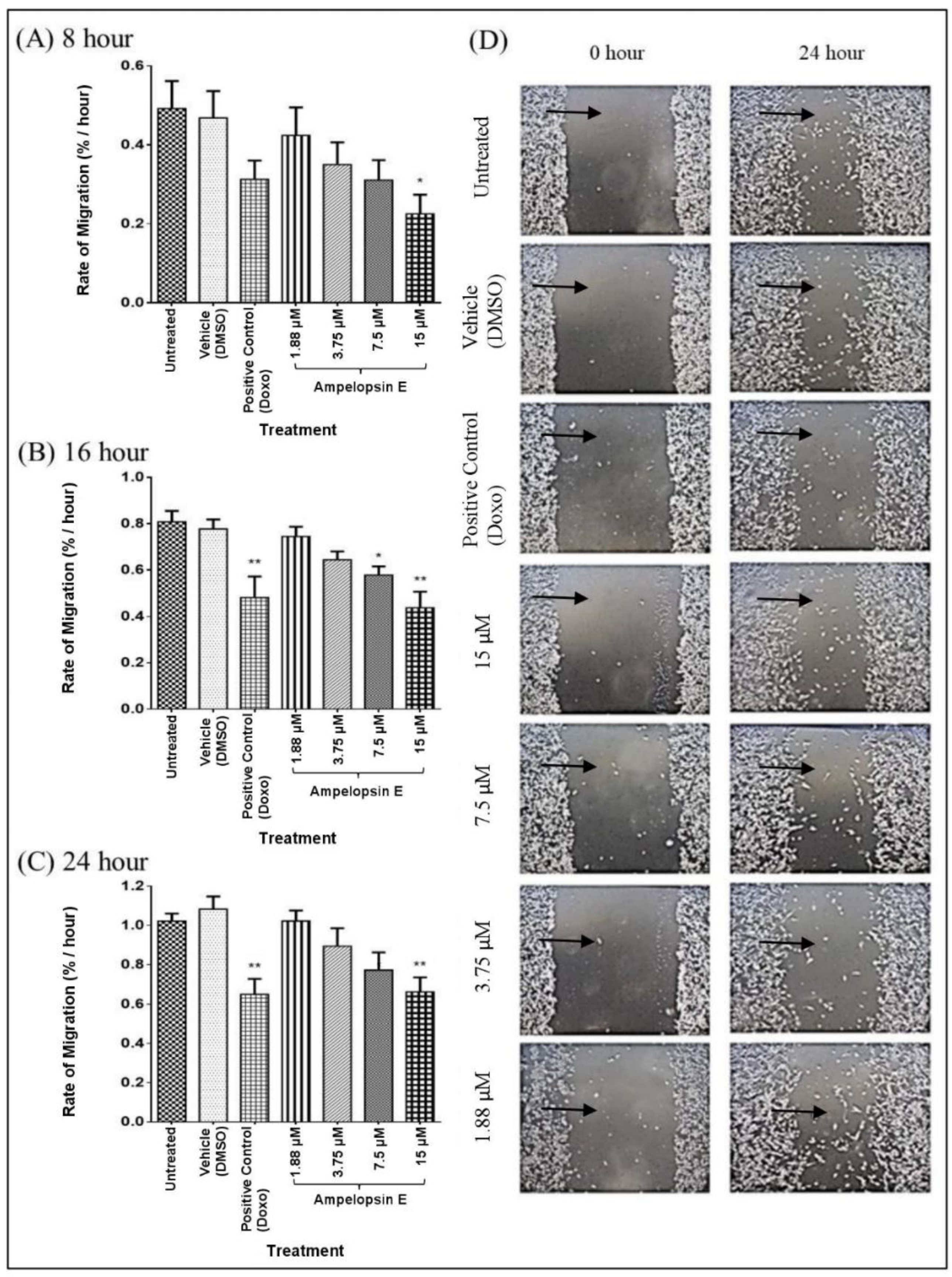 ) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05.
) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05.
) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05.
) over time using ‘Tscratch’ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05. ). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05, bars with ** indicate p < 0.01 and bars with *** indicate p < 0.001 when compared to untreated group.
). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05, bars with ** indicate p < 0.01 and bars with *** indicate p < 0.001 when compared to untreated group.
). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05, bars with ** indicate p < 0.01 and bars with *** indicate p < 0.001 when compared to untreated group.
). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with * indicate p < 0.05, bars with ** indicate p < 0.01 and bars with *** indicate p < 0.001 when compared to untreated group. ). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with *** indicated p < 0.001 when compared to untreated group.
). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with *** indicated p < 0.001 when compared to untreated group.
). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with *** indicated p < 0.001 when compared to untreated group.
). Number of fields = 12. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bar with *** indicated p < 0.001 when compared to untreated group. ) showed that invadopodia had degraded the coated gelatin. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. (B) Invadopodia formation in MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope. Total number of cells with invadopodia and total number of all cells were counted. Number of fields = 20 and number of cells > 100 per condition. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 when compared to untreated group.
) showed that invadopodia had degraded the coated gelatin. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. (B) Invadopodia formation in MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope. Total number of cells with invadopodia and total number of all cells were counted. Number of fields = 20 and number of cells > 100 per condition. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 when compared to untreated group.
) showed that invadopodia had degraded the coated gelatin. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. (B) Invadopodia formation in MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope. Total number of cells with invadopodia and total number of all cells were counted. Number of fields = 20 and number of cells > 100 per condition. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 when compared to untreated group.
) showed that invadopodia had degraded the coated gelatin. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. (B) Invadopodia formation in MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope. Total number of cells with invadopodia and total number of all cells were counted. Number of fields = 20 and number of cells > 100 per condition. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 when compared to untreated group. ) represent the area corresponds to degradation. Number of fields = 20 and number of cells > 100 per condition. Gelatin degradation was calculated by normalizing the area fraction to the number of nuclei. (B) Gelatin degradation of MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. Area of gelatin degradation was calculated using IMAGEJ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 and bars with **** indicate p < 0.0001 when compared to untreated group.
) represent the area corresponds to degradation. Number of fields = 20 and number of cells > 100 per condition. Gelatin degradation was calculated by normalizing the area fraction to the number of nuclei. (B) Gelatin degradation of MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. Area of gelatin degradation was calculated using IMAGEJ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 and bars with **** indicate p < 0.0001 when compared to untreated group.
) represent the area corresponds to degradation. Number of fields = 20 and number of cells > 100 per condition. Gelatin degradation was calculated by normalizing the area fraction to the number of nuclei. (B) Gelatin degradation of MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. Area of gelatin degradation was calculated using IMAGEJ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 and bars with **** indicate p < 0.0001 when compared to untreated group.
) represent the area corresponds to degradation. Number of fields = 20 and number of cells > 100 per condition. Gelatin degradation was calculated by normalizing the area fraction to the number of nuclei. (B) Gelatin degradation of MDA-MB-231 cells normalized to untreated group. Cells were divided into untreated group, vehicle, positive control and four concentrations of ampelopsin E (1.88, 3.75, 7.5 and 15 µM) at 24 h. Images were captured using a fluorescent light microscope at 200× magnification. Number of fields = 20 and number of cells > 100 per condition. Area of gelatin degradation was calculated using IMAGEJ analysis software. Results were expressed as mean ± SEM of four independent experiments, n = 4. Bars with *** indicate p < 0.001 and bars with **** indicate p < 0.0001 when compared to untreated group.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tieng, F.Y.F.; Latifah, S.Y.; Md Hashim, N.F.; Khaza’ai, H.; Ahmat, N.; Gopalsamy, B.; Wibowo, A. Ampelopsin E Reduces the Invasiveness of the Triple Negative Breast Cancer Cell Line, MDA-MB-231. Molecules 2019, 24, 2619. https://doi.org/10.3390/molecules24142619
Tieng FYF, Latifah SY, Md Hashim NF, Khaza’ai H, Ahmat N, Gopalsamy B, Wibowo A. Ampelopsin E Reduces the Invasiveness of the Triple Negative Breast Cancer Cell Line, MDA-MB-231. Molecules. 2019; 24(14):2619. https://doi.org/10.3390/molecules24142619
Chicago/Turabian StyleTieng, Francis Yew Fu, Saiful Yazan Latifah, Nur Fariesha Md Hashim, Huzwah Khaza’ai, Norizan Ahmat, Banulata Gopalsamy, and Agustono Wibowo. 2019. "Ampelopsin E Reduces the Invasiveness of the Triple Negative Breast Cancer Cell Line, MDA-MB-231" Molecules 24, no. 14: 2619. https://doi.org/10.3390/molecules24142619
APA StyleTieng, F. Y. F., Latifah, S. Y., Md Hashim, N. F., Khaza’ai, H., Ahmat, N., Gopalsamy, B., & Wibowo, A. (2019). Ampelopsin E Reduces the Invasiveness of the Triple Negative Breast Cancer Cell Line, MDA-MB-231. Molecules, 24(14), 2619. https://doi.org/10.3390/molecules24142619