
Hydrophobic Interactions Drive Binding between Vascular Endothelial Growth Factor-A (VEGFA) and Polyphenolic Inhibitors


Abstract
:
1. Introduction
2. Results and Discussion
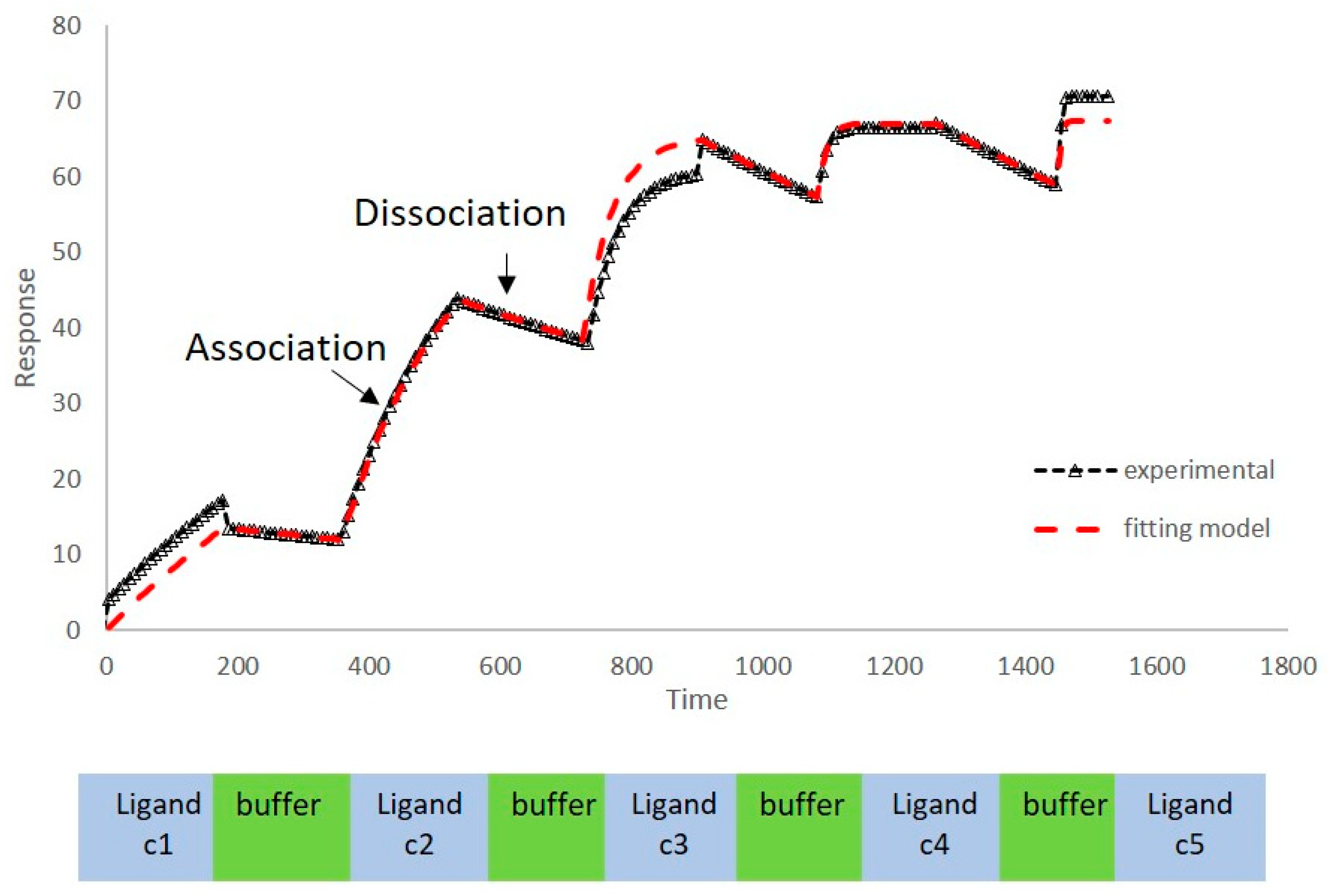
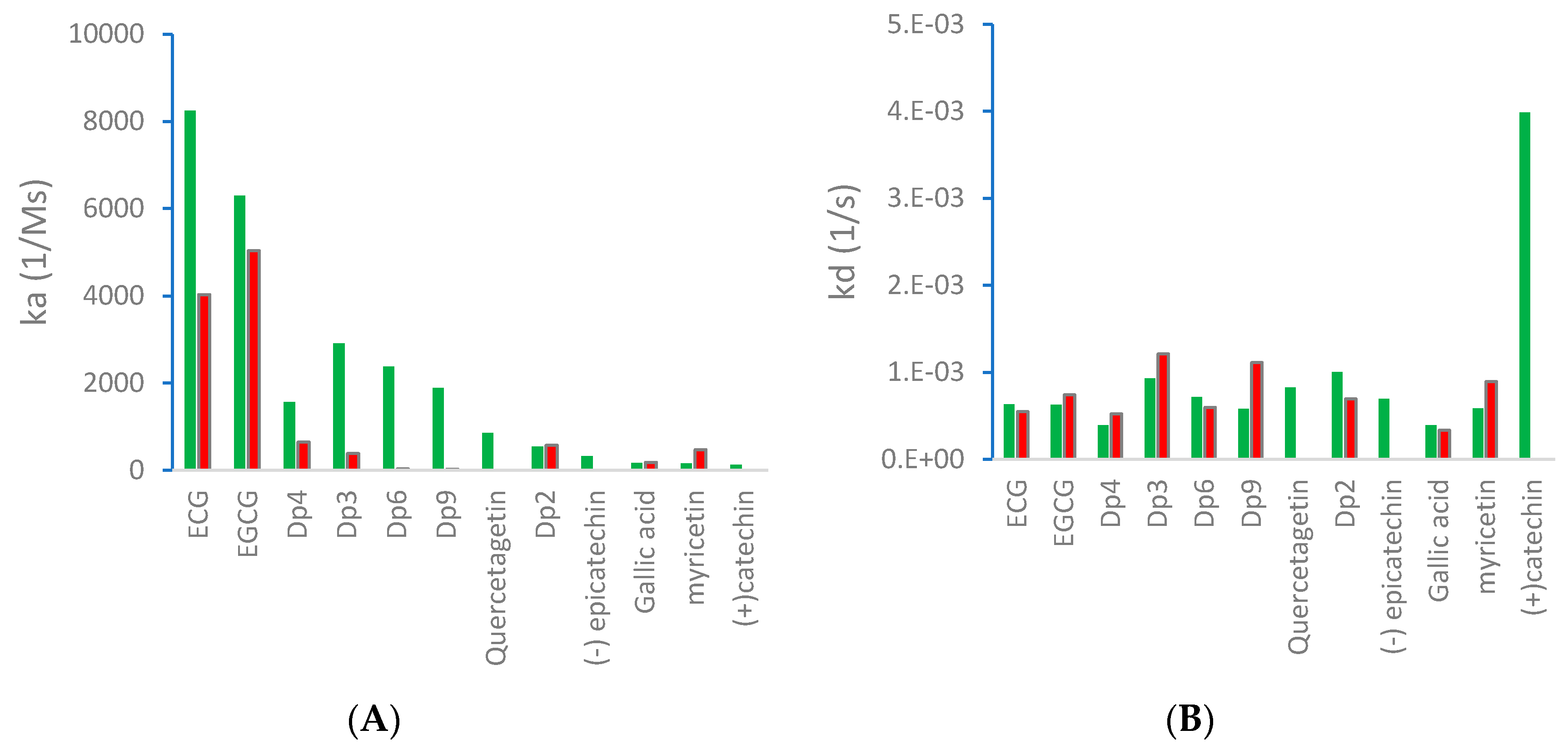
2.1. Kinetics of Binding between VEGF and Polyphenols
2.2. Effect of Physico-Chemical Conditions on Kinetics of Polyphenol-VEGF Interactions
2.2.1. Effect of Temperature in Binding of VEGF and Polyphenols
2.2.2. Effect of Environmental Conditions on VEGF-EGCG Interactions at 37 °C
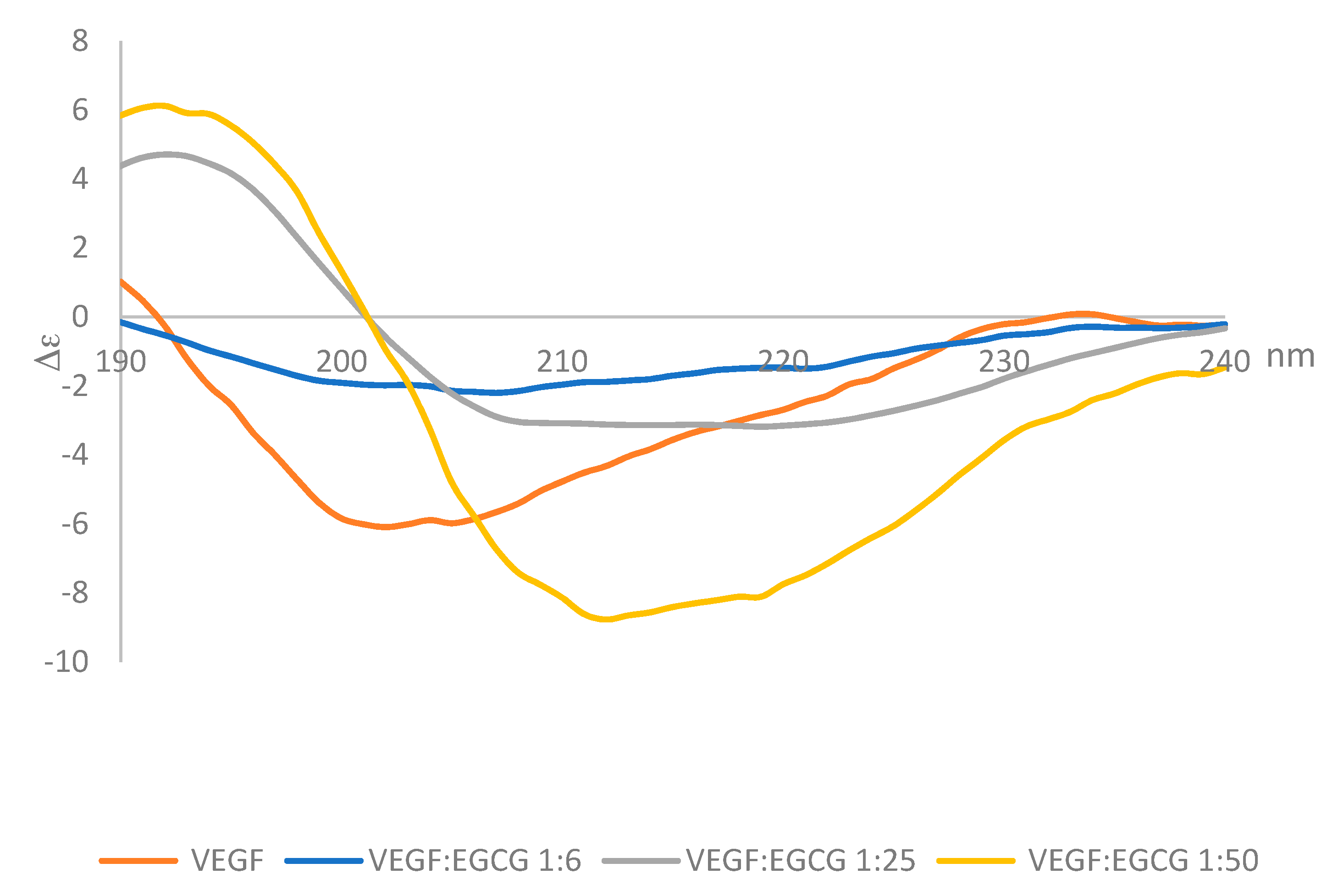
2.3. Circular Dichroism
3. Materials and Methods
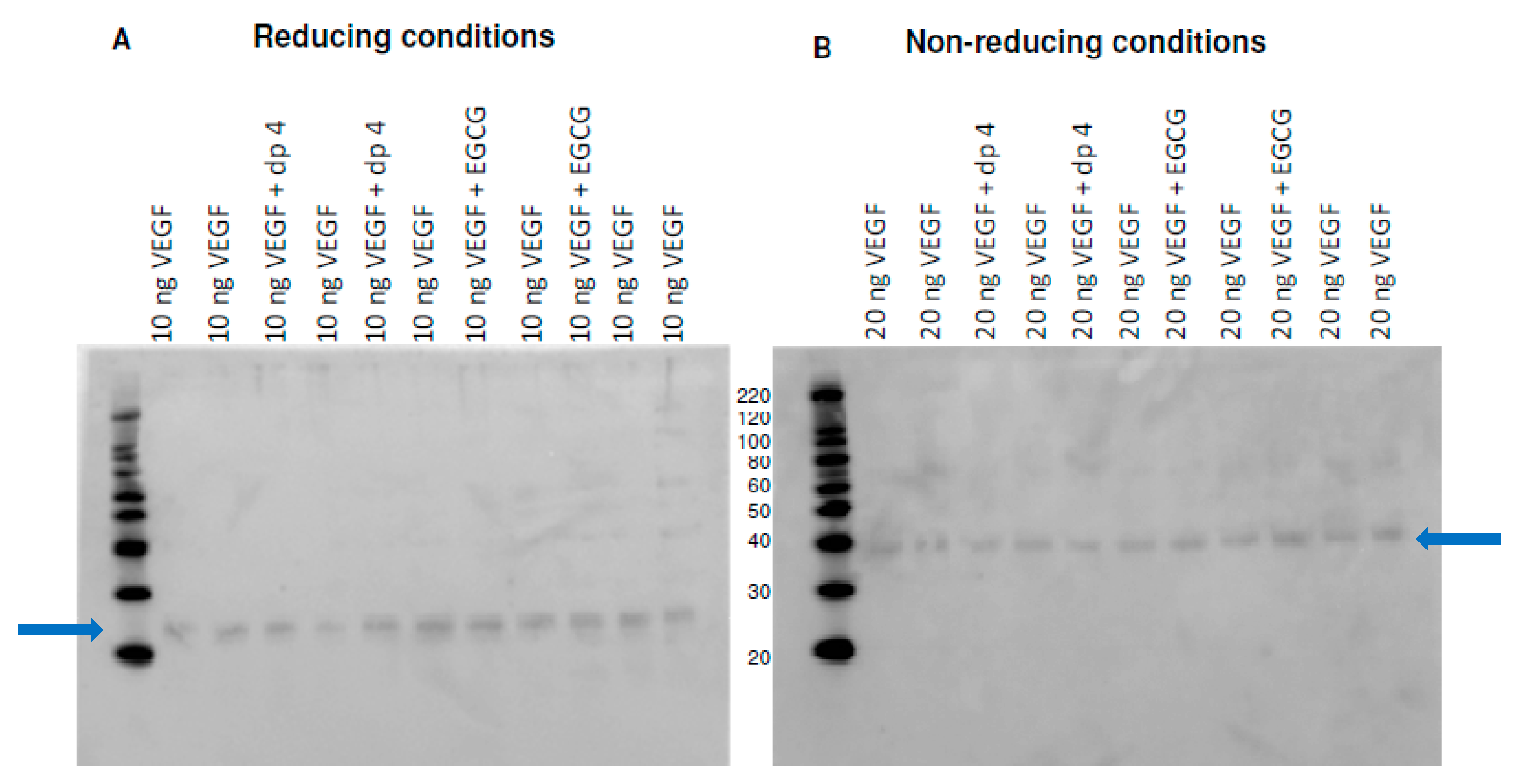
3.1. Materials
3.2. Surface Plasmon Resonance Studies
3.3. Far-UV Circular Dichroism Spectroscopy
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bhisitkul, R.B. Vascular endothelial growth factor biology: Clinical implications for ocular treatments. Br. J. Ophthalmol. 2006, 90, 1542–1547. [Google Scholar] [CrossRef] [PubMed]
- Moyle, C.W.; Cerezo, A.B.; Winterbone, M.S.; Hollands, W.J.; Alexeev, Y.; Needs, P.W.; Kroon, P.A. Potent inhibition of VEGFR-2 activation by tight binding of green tea epigallocatechin gallate and apple procyanidins to VEGF: Relevance to angiogenesis. Mol. Nutr. Food Res. 2015, 59, 401–402. [Google Scholar] [CrossRef] [PubMed]
- Cerezo, A.B.; Winterbone, M.S.; Moyle, C.W.; Veeds, P.W.; Kroon, P.A. Molecular structure-function relationship of dietary polyphenols for inhibiting VEGF-induced VEGFR-2 activity. Mol. Nutr. Food Res. 2015, 59, 2119–2131. [Google Scholar] [CrossRef]
- Edwards, R.L.; Kroon, P.A. Inhibition of VEGF signaling by polyphenols in relation to atherosclerosis and cardiovascular disease. In Recent Advances in Polyphenol Research; John Wiley & Sons, Ltd.: Chichester, UK, 2014; Volume 8, pp. 281–325. [Google Scholar]
- Brudzynski, K.; Maldonado-Alvarez, L. Polyphenol-Protein Complexes and Their Consequences for the Redox Activity, Structure and Function of Honey. A Current View and New Hypothesis–A Review. Pol. J. Food Nutr. Sci. 2015, 65, 71–80. [Google Scholar] [CrossRef]
- Chandler, D. Interfaces and the driving force of hydrophobic assembly. Nature 2005, 437, 640–647. [Google Scholar] [CrossRef]
- Hollands, W.J.; Voorspoels, S.; Jacobs, G.; Aaby, K.; Meisland, A.; Garcia-Villalba, R.; Tomas-Barberan, F.; Piskula, M.K.; Mawson, D.; Vovk, I.; et al. Development, validation and evaluation of an analytical method for the determination of monomeric and oligomeric procyanidins in apple extracts. J. Chromatogr. A 2017, 1495, 46–56. [Google Scholar] [CrossRef]
- Chen, Y.; Wiesmann, C.; Fuh, G.; Li, B.; Christinger, H.W.; McKay, P.; de Vos, A.M.; Lowman, H.B. Selection and analysis of an optimized anti-VEGF antibody: Crystal structure of an affinity matured Fab in complex with antigen. J. Mol. Biol. 1999, 293, 865–881. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. Protein Secondary Structure Analyses from Circular Dichroism Spectroscopy: Methods and Reference Databases. Biopolymers 2008, 89, 392–400. [Google Scholar] [CrossRef]
- Whitmore, L.; Wallace, B.A. DICHROWEB, an online server for protein secondary structure analyses from circular dichroism spectroscopic data. Nucleic Acids Res. 2004, 32, 668–673. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ka (1/Ms) | kd (1/s) | KD (M) | IC50 (µM) | ||
|---|---|---|---|---|---|
| Epicatechin gallate (ECG) | 8240 | 6.33 × 10−4 | 3.48 × 10−8 | 0.16 | 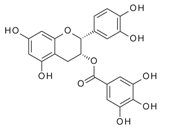 |
| Epigallocatechin gallate (EGCG) | 6300 | 6.25 × 10−4 | 1.12 × 10−7 | 0.09 |  |
| Procyanidin Dp3 | 2910 | 9.30 × 10−4 | 3.51 × 10−7 | 0.78 | 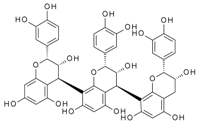 |
| Procyanidin Dp6 | 2377 | 7.14 × 10−4 | 9.79 × 10−6 | ND |  |
| Procyanidin Dp9 | 1893.5 | 5.81 × 10−4 | 3.41 × 10−7 | ND | 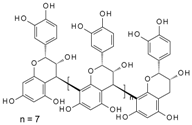 |
| Procyanidin Dp4 | 1566.5 | 3.91 × 10−4 | 2.77 × 10−7 | 0.28 | 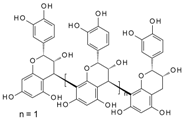 |
| Quercetagetin | 857.7 | 8.26 × 10−4 | 9.34 × 10−7 | 0.10 | 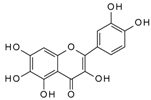 |
| Procyanidin Dp2 | 548.4 | 1.00 × 10−3 | 6.37 × 10−7 | 52.6 |  |
| (-) Epicatechin | 323.1 | 6.96 × 10-4 | 2.71 × 10-6 | ND | 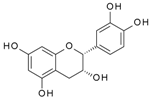 |
| Gallic acid | 166.3 | 3.91 × 10−4 | 5.52 × 10−6 | ND |  |
| Myricetin | 153.4 | 5.83 × 10−4 | 4.46 × 10−6 | 0.12 | 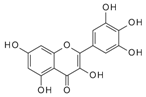 |
| (+) Catechin | 130.6 | 3.99 × 10−3 | 2.46 × 10−5 | 215 |  |
| ka (1/Ms) | kd (1/s) | KD (M) | |
|---|---|---|---|
| PBST | 5184 ± 64 | 8.8 × 10−4 ± 5.2 × 10−5 | 1.7 × 10−7 |
| PBST + 1 M urea | 5160 ± 100 | 3.6 × 10−4 ± 2.1 × 10−5 | 7.1 × 10−8 |
| PBST | 6147 ± 52 | 6.8 × 10−4 ± 2.9 × 10−6 | 1.1 × 10−7 |
| PBST + 0.5 M NaCl | 5184.3 ± 92 | 3.7 × 10−4 ± 4 × 10−5 | 6.0 × 10−8 |
| PBST + 1 M NaCl | 6502.2 ± 260 | 3.3 × 10−4 ± 4 × 10−5 | 3.5 × 10−8 |
| PBST | 6052 ± 64 | 7.2 × 10−4 ± 5 × 10−5 | 1.4 × 10−7 |
| PBST + 10% ethanol | 2464.7 ± 60 | 1.7 × 10−3 ± 7 × 10−5 | 8.1 × 10−7 |
| PBST + 30% ethanol | 481.9 ± 21 | 2.0 × 10−4 ± 5 × 10−5 | 4.7 × 10−7 |
| PBST | 6147 ± 52 | 6.8 × 10−4 ± 2.9 × 10−6 | 1.1 × 10−7 |
| PBST + 10 M ethylene glycol | 796.1 ± 45 | 2.6 × 10−4 ± 5.2 × 10−5 | 4.2 × 10−7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez-Moral, N.; Needs, P.W.; Moyle, C.W.A.; Kroon, P.A. Hydrophobic Interactions Drive Binding between Vascular Endothelial Growth Factor-A (VEGFA) and Polyphenolic Inhibitors. Molecules 2019, 24, 2785. https://doi.org/10.3390/molecules24152785
Perez-Moral N, Needs PW, Moyle CWA, Kroon PA. Hydrophobic Interactions Drive Binding between Vascular Endothelial Growth Factor-A (VEGFA) and Polyphenolic Inhibitors. Molecules. 2019; 24(15):2785. https://doi.org/10.3390/molecules24152785
Chicago/Turabian StylePerez-Moral, Natalia, Paul W. Needs, Christina W.A. Moyle, and Paul A. Kroon. 2019. "Hydrophobic Interactions Drive Binding between Vascular Endothelial Growth Factor-A (VEGFA) and Polyphenolic Inhibitors" Molecules 24, no. 15: 2785. https://doi.org/10.3390/molecules24152785