
Efficacy of ARV-1502, a Proline-Rich Antimicrobial Peptide, in a Murine Model of Bacteremia Caused by Multi-Drug Resistant (MDR) Acinetobacter baumannii


Abstract
:1. Introduction
2. Results
2.1. In Vitro Susceptibility of ARV-1502 Alone and in Combination with IPM
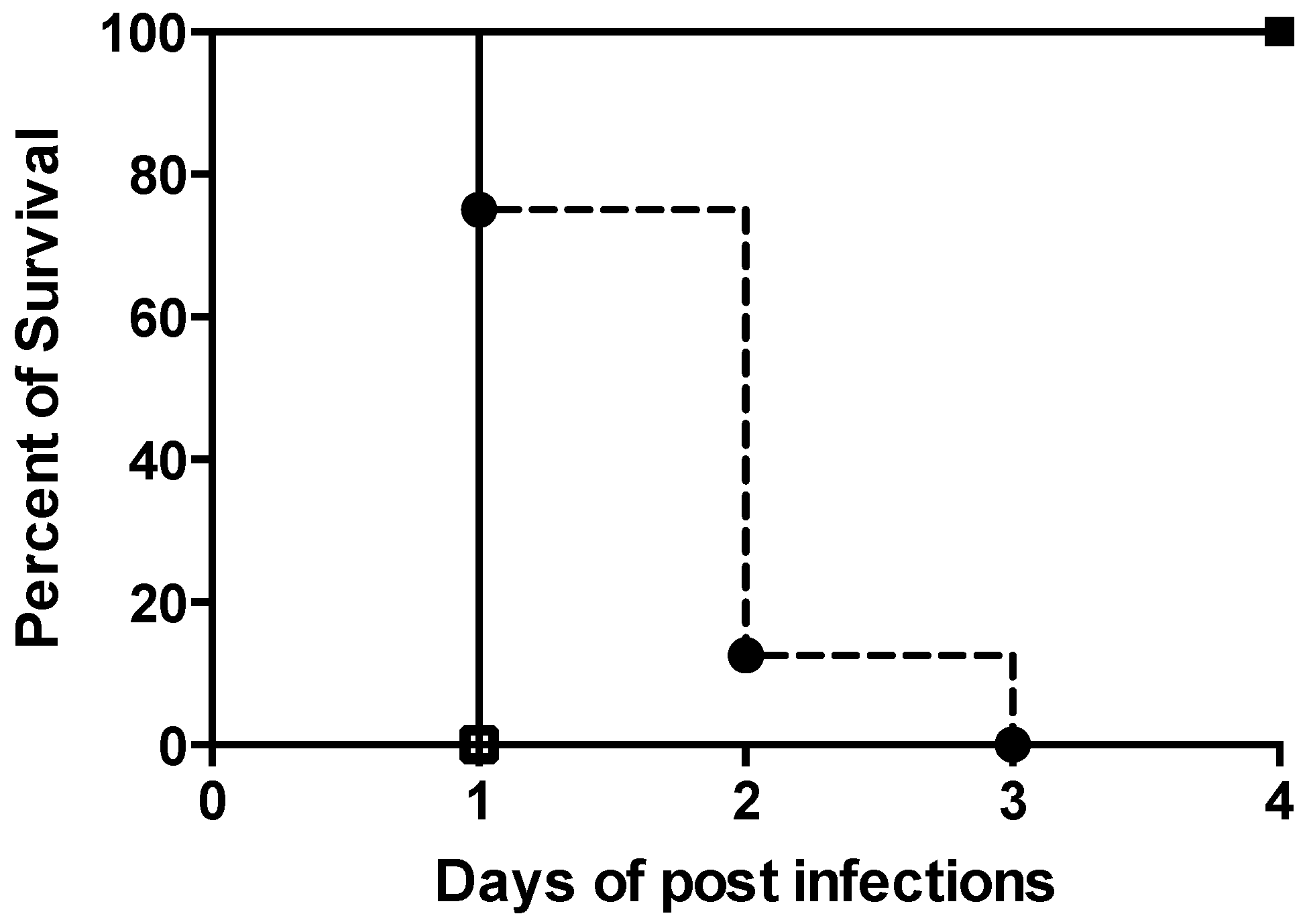
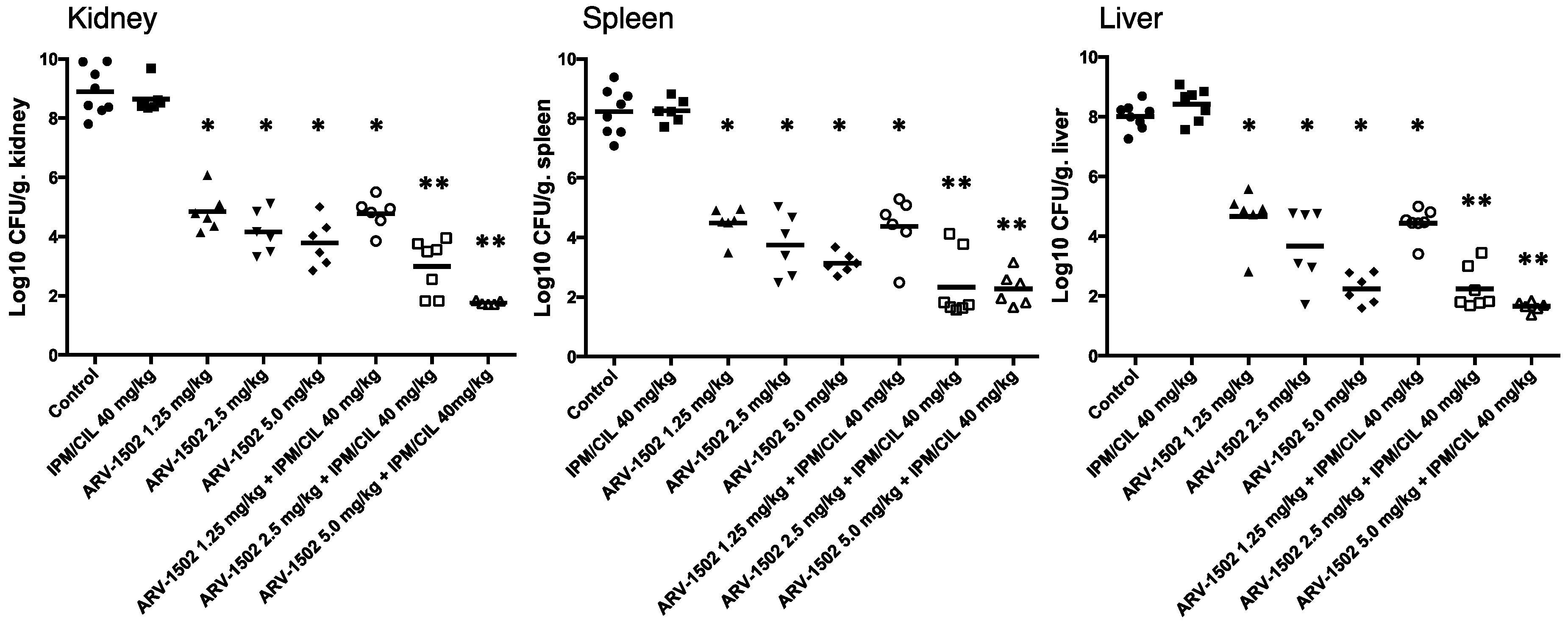
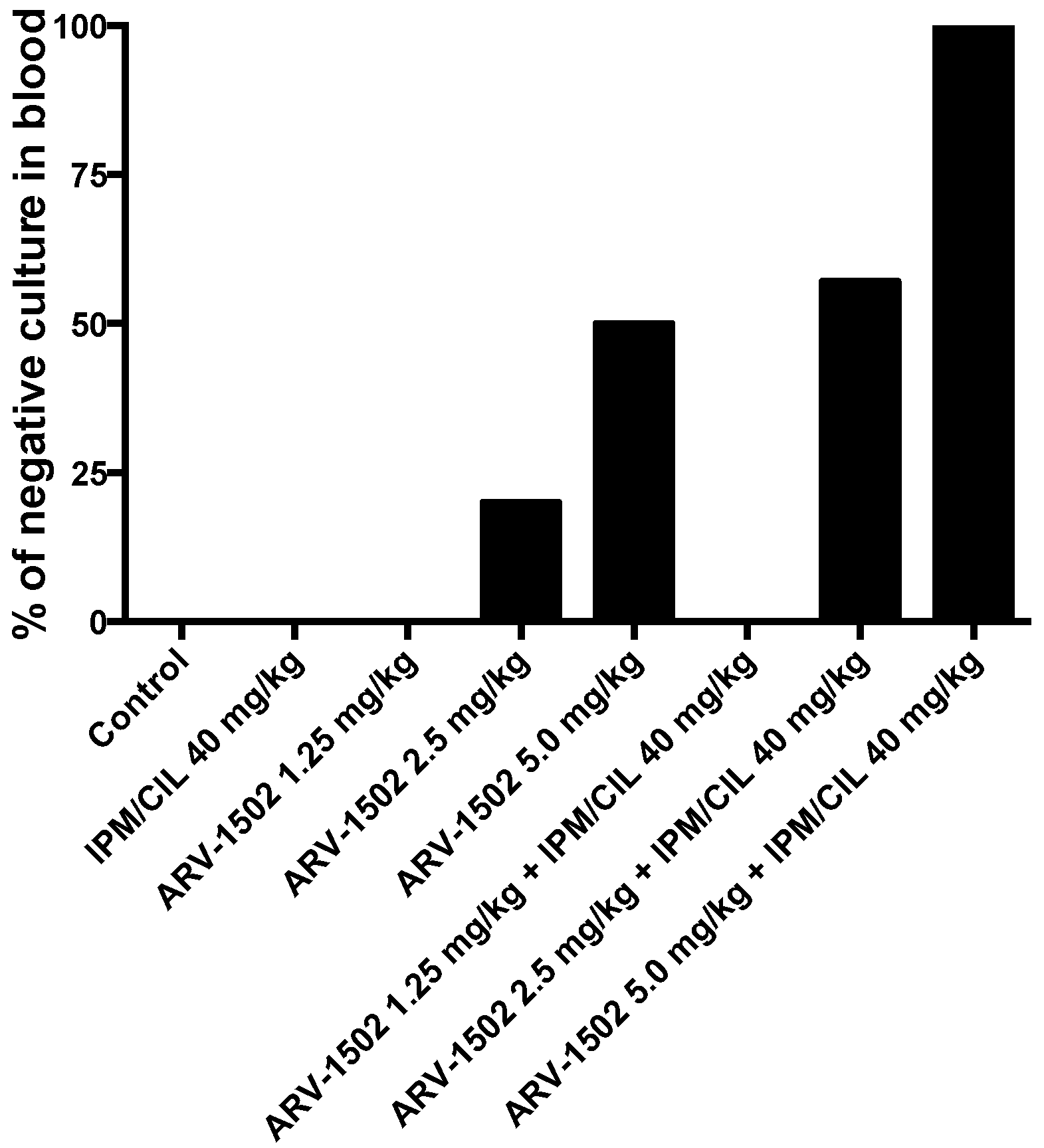
2.2. Therapeutic Efficacy of ARV-1502 Alone and in Combination with IPM/CIL in a Murine Model of Bacteremia due to the MDR A. Baumannii Strain HUMC1
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Growth Conditions
4.2. Molecular Design and Synthesis of ARV-1502
4.3. Determination of MICs
4.4. Fractional Inhibitory Concentration Index (FICI) Assay
4.5. ID95 in a Murine Bacteremia Model due to A. Baumannii (HUMC1)
4.6. Efficacy of ARV-1502 and IPM/CIL Alone, and in Combination in A. Baumannii (HUMC1)—Challenged Murine Bacteremia
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Akova, M. Epidemiology of antimicrobial resistance in bloodstream infections. Virulence 2016, 7, 252–266. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. P.T. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Pfalzgraff, A.; Brandenburg, K.; Weindl, G. Antimicrobial Peptides and Their Therapeutic Potential for Bacterial Skin Infections and Wounds. Front Pharmacol. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Bayer, A.S. Antimicrobial peptides versus invasive infections. Curr. Top Microbiol. Immunol. 2006, 306, 111–152. [Google Scholar] [PubMed]
- Otvos, L.; Otvos, L., Jr.; Rogers, M.E.; Consolvo, P.J.; Condie, B.A.; Lovas, S. Interaction between heat shock proteins and antimicrobial peptides. Biochemistry 2000, 39, 14150–14159. [Google Scholar] [CrossRef] [PubMed]
- Chiappori, F.; Fumian, M.; Milanesi, L.; Merelli, I. DnaK as Antibiotic Target: Hot Spot Residues Analysis for Differential Inhibition of the Bacterial Protein in Comparison with the Human HSP70. PLoS ONE 2015, 10, e0124563. [Google Scholar] [CrossRef] [PubMed]
- Zahn, M.; Berthold, N.; Kieslich, B.; Knappe, D.; Hoffmann, R.; Strater, N. Structural studies on the forward and reverse binding modes of peptides to the chaperone DnaK. J. Mol. Biol. 2013, 425, 2463–2479. [Google Scholar] [CrossRef]
- Otvos, L.J.; Wade, J.D.; Lin, F.; Condie, B.A.; Hanrieder, J.; Hoffmann, R. Designer antibacterial peptides kill fluoroquinolone-resistant clinical isolates. J. Med. Chem. 2005, 48, 5349–5359. [Google Scholar] [CrossRef]
- Kragol, G.; Lovas, S.; Varadi, G.; Condie, B.A.; Hoffmann, R.; Otvos, L. The antibacterial peptide pyrrhocoricin inhibits the ATPase actions of DnaK and prevents chaperone-assisted protein folding. Biochemistry 2001, 40, 3016–3026. [Google Scholar] [CrossRef]
- Otvos, L.J.; Snyder, C.; BCondie, B.; Bulet, P.; Wade, J.D. Chimeric antimicrobial peptides exhibit multiple modes of action. Int. J. Pept. Res. Ther. 2005, 11, 29–42. [Google Scholar] [CrossRef]
- Szabo, D.; Ostorhazi, E.; Binas, A.; Rozgonyi, F.; Kocsis, B.; Cassone, M. The designer proline-rich antibacterial peptide A3-APO is effective against systemic Escherichia coli infections in different mouse models. Int. J. Antimicrob. Agents 2010, 35, 357–361. [Google Scholar] [CrossRef]
- Rozgonyi, F.; Szabo, D.; Kocsis, B.; Ostorhazi, E.; Abbadessa, G.; Cassone, M. The antibacterial effect of a proline-rich antibacterial peptide A3-APO. Curr. Med. Chem. 2009, 16, 3996–4002. [Google Scholar] [CrossRef]
- Ostorhazi, E.; Rozgonyi, F.; Sztodola, A.; Harmos, F.; Kovalszky, I.; Szabo, D.; Bonomo, R. A Preclinical advantages of intramuscularly administered peptide A3-APO over existing therapies in Acinetobacter baumannii wound infections. J. Antimicrob. Chemother. 2010, 65, 2416–2422. [Google Scholar] [CrossRef]
- Cassone, M.; Vogiatzi, P.; La Montagna, R.; Inacio, V.D.O.; Cudic, P.; Wade, J.D.; Otvos, L., Jr. Scope and limitations of the designer proline-rich antibacterial peptide dimer, A3-APO, alone or in synergy with conventional antibiotics. Peptides 2008, 29, 1878–1886. [Google Scholar] [CrossRef]
- Noto, P.B.; Abbadessa, G.; Cassone, M.; Mateo, G.D.; Agelan, A.; Wade, J.D.; Otvos, L., Jr. Alternative stabilities of a proline-rich antibacterial peptide in vitro and in vivo. Protein Sci. 2008, 17, 1249–1255. [Google Scholar] [CrossRef]
- Otvos, L., Jr.; Ostorhazi, E.; Szabo, D.; Zumbrun, S.D.; Miller, L.L.; Halasohoris, S.L.; Kraus, C.N. Synergy Between Proline-Rich Antimicrobial Peptides and Small Molecule Antibiotics Against Selected Gram-Negative Pathogens in vitro and in vivo. Front Chem. 2018, 6, 309. [Google Scholar] [CrossRef]
- Li, W.; O’Brien-Simpson, N.M.; Tailhades, J.; Pantarat, N.; Dawson, R.M.; Otvos, L., Jr.; Wade, J.D. Multimerization of a Proline-Rich Antimicrobial Peptide, Chex-Arg20, Alters Its Mechanism of Interaction with the Escherichia coli Membrane. Chem. Biol. 2015, 22, 1250–1258. [Google Scholar] [CrossRef]
- Ostorhazi, E.; Holub, M.C.; Rozgonyi, F.; Harmos, F.; Cassone, M.; Wade, J.D.; Otvos, L., Jr. Broad-spectrum antimicrobial efficacy of peptide A3-APO in mouse models of multidrug-resistant wound and lung infections cannot be explained by in vitro activity against the pathogens involved. Int. J. Antimicrob. Agents. 2011, 37, 480–484. [Google Scholar] [CrossRef]
- Ostorhazi, E.; Hoffmann, R.; Herth, N.; Wade, J.D.; Kraus, C.N.; Otvos, L. Advantage of a Narrow Spectrum Host Defense (Antimicrobial) Peptide Over a Broad Spectrum Analog in Preclinical Drug Development. Front Chem. 2018, 6, 359. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sun, Z.; O’Brien-Simpson, N.M.; Otvos, L.; Reynolds, E.C.; Hossain, M.A. The Effect of Selective D- or N(alpha)-Methyl Arginine Substitution on the Activity of the Proline-Rich Antimicrobial Peptide, Chex1-Arg20. Front Chem. 2017, 5, 1. [Google Scholar] [CrossRef]
- Knappe, D.; Adermann, K.; Hoffmann, R. Oncocin Onc72 is efficacious against antibiotic-susceptible Klebsiella pneumoniae ATCC 43816 in a murine thigh infection model. Biopolymers 2015, 104, 707–711. [Google Scholar] [CrossRef]
- Knappe, D.; Fritsche, S.; Alber, G.; Kohler, G.; Hoffmann, R.; Muller, U. Oncocin derivative Onc72 is highly active against Escherichia coli in a systemic septicaemia infection mouse model. J. Antimicrob. Chemother. 2012, 67, 2445–2451. [Google Scholar] [CrossRef] [Green Version]
- Ostorhazi, E.; Rozgonyi, F.; Szabo, D.; Binas, A.; Cassone, M.; Wade, J.D.; Otvos, L., Jr. Intramuscularly administered peptide A3-APO is effective against carbapenem-resistant Acinetobacter baumannii in mouse models of systemic infections. Biopolymers 2011, 96, 126–129. [Google Scholar] [CrossRef] [Green Version]
- Ostorhazi, E.; Voros, E.; Nemes-Nikodem, E.; Pinter, D.; Sillo, P.; Mayer, B.; Otvos, L., Jr. Rapid systemic and local treatments with the antibacterial peptide dimer A3-APO and its monomeric metabolite eliminate bacteria and reduce inflammation in intradermal lesions infected with Propionibacterium acnes and meticillin-resistant Staphylococcus aureus. Int. J. Antimicrob. Agents 2013, 42, 537–543. [Google Scholar] [CrossRef]
- Otvos, L., Jr. Immunomodulatory effects of anti-microbial peptides. Acta Microbiol. Immunol. Hung. 2016, 63, 257–277. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, J.M.; Oliveira, M.D.; Franco, O.L.; Migliolo, L.; de Melo, C.P.; Andrade, C.A. Elucidation of mechanisms of interaction of a multifunctional peptide Pa-MAP with lipid membranes. Biochim. Biophys. Acta 2014, 1838, 2899–2909. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, K.W.; Pantapalangkoor, P.; Nielsen, T.; Tan, B.; Junus, J.; Hujer, K.M. Host fate is rapidly determined by innate effector-microbial interactions during Acinetobacter baumannii bacteremia. J. Infect. Dis. 2015, 211, 1296–1305. [Google Scholar] [CrossRef]
- Luo, G.; Lin, L.; Ibrahim, A.S.; Baquir, B.; Pantapalangkoor, P.; Bonomo, R.A.; Spellberg, B. Active and passive immunization protects against lethal, extreme drug resistant-Acinetobacter baumannii infection. PLoS ONE 2012, 7, e29446. [Google Scholar] [CrossRef]
- Sebe, I.; Ostorhazi, E.; Fekete, A.; Kovacs, K.N.; Zelko, R.; Kovalszky, I.; Otvos, L. Polyvinyl alcohol nanofiber formulation of the designer antimicrobial peptide APO sterilizes Acinetobacter baumannii-infected skin wounds in mice. Amino Acids 2016, 48, 203–211. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Test for Bacteria that Grow Aerobically; Approved StanSdard: Wayne, PA, USA, 2015. [Google Scholar]
- Jenkins, S.G.; Schuetz, A.N. Current concepts in laboratory testing to guide antimicrobial therapy. Mayo Clin. Proc. 2012, 87, 290–308. [Google Scholar] [CrossRef]
- Swanson, D.J.; DeAngelis, C.; Smith, I.L.; Schentag, J.J. Degradation kinetics of imipenem in normal saline and in human serum. Antimicrob. Agents Chemother. 1986, 29, 936–937. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Cell lines are available are upon request from the author. Samples of the compounds will be available once commercialized. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Doses (No. Animals) | Mean log10 CFU/g. Tissue ± SD (% Sterile Culture) | ||
|---|---|---|---|
| Kidney | Spleen | Liver | |
| 109 CFU (n = 6) | 8.71 ± 0.21 (0%) | 9.71 ± 0.31 (0%) | 9.33 ± 0.21 (0%) |
| 108 CFU (n = 8) | 8.90 ± 0.80 (0%) | 8.22 ± 0.79 (0%) | 8.01 ± 0.44 (0%) |
| 107 CFU (n = 6) | 2.41 ± 0.82 (50%) | 1.86 ± 0.15 (100%) | 2.08 ± 0.39 (66.7%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Y.Q.; Li, L.; Zhou, Y.; Kraus, C.N. Efficacy of ARV-1502, a Proline-Rich Antimicrobial Peptide, in a Murine Model of Bacteremia Caused by Multi-Drug Resistant (MDR) Acinetobacter baumannii. Molecules 2019, 24, 2820. https://doi.org/10.3390/molecules24152820
Xiong YQ, Li L, Zhou Y, Kraus CN. Efficacy of ARV-1502, a Proline-Rich Antimicrobial Peptide, in a Murine Model of Bacteremia Caused by Multi-Drug Resistant (MDR) Acinetobacter baumannii. Molecules. 2019; 24(15):2820. https://doi.org/10.3390/molecules24152820
Chicago/Turabian StyleXiong, Yan Q., Liang Li, Yufeng Zhou, and Carl N. Kraus. 2019. "Efficacy of ARV-1502, a Proline-Rich Antimicrobial Peptide, in a Murine Model of Bacteremia Caused by Multi-Drug Resistant (MDR) Acinetobacter baumannii" Molecules 24, no. 15: 2820. https://doi.org/10.3390/molecules24152820
APA StyleXiong, Y. Q., Li, L., Zhou, Y., & Kraus, C. N. (2019). Efficacy of ARV-1502, a Proline-Rich Antimicrobial Peptide, in a Murine Model of Bacteremia Caused by Multi-Drug Resistant (MDR) Acinetobacter baumannii. Molecules, 24(15), 2820. https://doi.org/10.3390/molecules24152820